
Abstract
High-quality biosamples are valuable resources for biomedical research. However, some tissues are stored without being sectioned into small aliquots and have to undergo repeated freeze–thaw cycles throughout prolonged experimentation. Little is known regarding the effects of repeated freeze–thaw cycles on the quality of biomolecules in tissues. The aim of this study was to evaluate the impact of repeated freeze–thaw (at room temperature or on ice) cycles on biomolecules and gene expression in four different types of tissues. Each fresh tissue was sectioned into seven aliquots and snap-frozen before undergoing repeated freeze–thaw cycles at room temperature or on ice. Biomolecules were extracted and analyzed. Both relative and absolute quantification were used to detect the changes in gene expression. The results indicated that the impact of repeated freeze–thaw cycles on RNA integrity varied by tissue type. Gene expression, including the housekeeping gene, was affected in RNA-degraded samples according to absolute quantification rather than relative quantification. Furthermore, our results suggest that thawing on ice could protect RNA integrity compared with thawing at room temperature. No obvious degradation of protein or DNA was observed with repeated freeze–thaw cycles either at room temperature or on ice. This research provides ample evidence for the necessity of sectioning fresh tissues into small aliquots before snap-freezing, thus avoiding degradation of RNA and alteration of gene expression resulting from repeated freeze–thaw cycles. For frozen tissue samples that were already in storage and had to be used repeatedly during their lifecycle, thawing on ice or sectioned at ultralow temperature is recommended.
Introduction
H
A number of studies have evaluated the impact of warm ischemia and preservation conditions on the quality of tissue samples.2–4 However, only a few studies have explored the impact of repeated freeze–thaw cycles on the quality of biomolecules, including RNA, protein, and DNA, in different types of tissues. It is known that the thawing process is capable of rupturing most cell structures and activating intrinsic enzymes, resulting in detrimental effects on biomolecules while also introducing exogenous RNase that could rapidly damage intracellular RNA.5,6 For most biobanking projects, storage in liquid nitrogen is regarded as the gold standard. With respect to the thawing process, however, there are no unified standards. So far, frozen tissue samples are generally thawed and processed at room temperature.4,7,8 Given that most biomolecules are temperature sensitive, the issue of whether a lower thawing temperature is more suitable for the thawing process needs to be investigated.
Therefore, the aim of our study is to investigate (1) the influence of repeated freeze–thaw cycles on biomolecules, including RNA, protein, and DNA, within four different types of tissues; (2) the influence of repeated freeze–thaw cycles on gene expression; and (3) potentially more suitable thawing methods to be employed when tissues have to undergo repeated freeze–thaw cycles.
Methods
Ethics statement
The study was approved by the Ethics Review Board of Shanghai Children's Hospital Affiliated to Shanghai Jiao Tong University. We performed all animal surgeries under anesthesia and made all efforts to reduce animal suffering.
Tissue collection and processing
Fifteen 3-week-old Sprague-Dawley rats were used in these experiments. Each part of the molecular experiments (RNA, protein, and DNA) used five rats as biological replicates. All rats were euthanized with sodium pentobarbital and then four types of tissues (liver, muscle obtained from the hind leg, brain, and testis) were obtained. Each sample was sectioned into seven fragments of about 0.4 × 0.4 × 0.4 cm (∼40 mg) and frozen in liquid nitrogen (−196°C) before being exposed to repeated freeze–thaw cycles. All procedures were performed on ice and finished within 30 minutes. The next day, the first fragment of each sample was extracted fresh frozen (FF), then three fragments underwent 1, 2, or 3 freeze–thaw cycles (RT1, RT2, and RT3, respectively) before extraction. Cycles consisted of 30 minutes of incubation at room temperature (thaw), followed by 30 minutes in liquid nitrogen (freeze). The remaining three aliquots were exposed to 2, 4, or 6 freeze–thaw cycles (ice2, ice4, and ice6, respectively) before extraction. Cycles consisted of 30 minutes of incubation on ice (thaw), followed by 30 minutes in liquid nitrogen (freeze).
RNA extraction and evaluation of quality
RNA extraction
Total RNA was isolated with TRIzol reagent (Invitrogen, SanDiego, CA) according to the manufacturer's instructions. Briefly, tissues were homogenized with 1 mL of TRIzol reagent. Subsequently, 200 μL of chloroform was added to extract RNA and 500 μL of isopropyl alcohol was added to precipitate RNA. Finally, pelleted RNA was washed with 75% ethanol twice, dried at room temperature, and dissolved in RNase-free water treated with diethylpyrocarbonate in a 55°C electro-thermostatic water bath for ∼10 minutes before being stored at −80°C.
RNA purity and integrity
The concentration and purity of RNA were measured by a NanoDrop™2000c spectrophotometer (Thermo Scientific, Waltham, MA). RNA samples at each time point were pooled and analyzed on the Agilent 2100 Bioanalyzer instrument using the RNA 6000 NanoLabChip kit (Agilent Technologies, Santa Clara, CA). The results are presented as RNA integrity number (RIN) values and virtual gel-like images, both of which were calculated from the Agilent software. RIN values range from 1 to 10, with 1 being the most degraded and 10 being the most intact RNA. In virtual gel-like images, two distinct bands of 28S and 18S represent good integrity of RNA. Smears behind the two main bands indicate severe RNA degradation.
Complementary DNA synthesis and quantitative real-time polymerase chain reaction
Complementary DNA synthesis
Complementary DNA (cDNA) was transcribed from 1 μg of total RNA in a final volume of 20 μL with a Prime Script™RT Master Mix kit (Takara, Japan) according to manufacturer's instructions. The newly synthesized cDNA was diluted fivefold in nuclease-free water and stored at −20°C.
Relative quantification
Real-time polymerase chain reaction (PCR)-based relative quantification was performed to measure the expression of nontissue-specific genes (Fos and Bcl-2) and tissue-specific genes (liver specific: Gpt and Alb, muscle specific: Tnni2 and Mb, brain specific: Gfap and NeuN, and testis specific: Acr and Ddx4) with SYBR Green Master Mix (Roche, Basel, Switzerland) in a final volume of 10 μL. Briefly, reactions consisted of 1 μL of cDNA, 2 μL of nuclease-free water, 5 μL of 2 × SYBR Super Mix, and 1 μL of forward and reverse primers (Sangon Biotech, Shanghai, China). The PCR reaction was conducted as follows: 10 min at 95°C, followed by 45 cycles at 95°C for 10 s and 60°C for 30 s. All reactions were performed in duplicate and amplified using the LightCycler 96 real-time PCR System (Roche, Basel, Switzerland). Finally, gene expression levels were normalized to the housekeeping gene (Gapdh) using the ΔΔCT method.
Absolute quantification
To investigate gene expression changes in the four types of tissues, the housekeeping gene (Gapdh), one nontissue-specific gene (Fos), and four tissue-specific genes (liver specific: Gpt, muscle specific: Tnni2, brain specific: NeuN, and testis specific: Ddx4) were chosen for real-time PCR-based absolute quantification. In the absolute quantification experiment, six recombinant plasmids, including the six genes (Sangon Biotech, Shanghai, China), were constructed. The copy numbers per volume were calculated using the average product molecular weight and Avogadro's constant. Seven orders of magnitude serial dilutions of plasmids, ranging from 1 × 108 to 1 × 102 copies/μL, were calculated using a standard curve. Real-time PCR reactions were performed in triplicate for each sample and the procedure was the same as described for the relative quantification experiments.
Protein extraction and western blot analysis
Protein extraction
Samples were divided into aliquots of 20 mg each and then submerged in 1 mL of Mammalian Protein Extraction Reagent (Thermo Scientific) with the protease inhibitor cocktail. After homogenization, the samples were incubated on ice for 30 minutes and centrifuged at 4°C for 15 minutes at 14,000 rpm. The supernatant was collected and the protein concentration was measured using a Pierce™ BCA Protein Assay Kit (Thermo Scientific).
Western blot analysis
Western blot for the FOS and GAPDH proteins was performed on all aliquots of the four types of tissues. Protein samples (10 μg) were separated by electrophoresis on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto a nitrocellulose filter membrane (YEASEN, Shanghai, China). After blocking for 1 hour with 5% nonfat milk in Tris-buffered saline with Tween-20 (TBS-T; Sigma Aldrich, St Louis, MO) at room temperature, the membranes were incubated overnight with rabbit anti-FOS antibody (1:4000 dilution; Abcam, Cambridge, United Kingdom) and mouse anti-GAPDH antibody (YEASEN, Shanghai, China) at 4°C. Then, the membranes were washed with TBS-T and incubated with peroxidase-conjugated secondary antibody: goat anti-rabbit and goat anti-mouse (both 1:2500 dilution; Jackson Immuno Research, Suffolk, United Kingdom) for 2 hours at room temperature. Image analysis was performed using a Chemi Scope 3500 (Clinx, Shanghai, China).
DNA extraction and agarose gel electrophoresis
DNA extraction
DNA was extracted using a DNeasy Blood and Tissue Kit (Qiagen, Venlo, Netherlands) according to the instructions provided by the manufacturer. Approximately 10 mg of each aliquot was submerged in a grind tube that contained 180 μL of lysis buffer and 20 μL of proteinase K. After homogenization, the homogenate was heated in a 56°C electro-thermostatic water bath for 10 minutes and then transferred to a DNA extraction column. Extracted DNA was dissolved with 100 μL of Tris-EDTA buffer. The concentration and purity of DNA samples were measured by a NanoDrop2000c spectrophotometer (Thermo Scientific).
Agarose gel electrophoresis
Agarose gel electrophoresis was conducted to investigate the integrity of the DNA. Extracted DNA samples from four tissues were evaluated by loading 150 ng of DNA on a 0.8% agarose gel for 45 min at 100 V. Image analysis was performed using a Chemi Scope 3500.
Statistical analysis
Statistical analysis was performed using SPSS Statistics 19.0 (IBM, Chicago, IL). The difference in RIN values among the different groups was analyzed using one-way ANOVA; *p values <0.05 were considered to be statistically significant.
Results
The effects of repeated freeze–thaw cycles on RNA integrity varied with tissue type
To evaluate the impact of repeated freeze–thaw cycles on the quality of biomolecules, four different types of tissues (liver, muscle, brain, and testis) were obtained. In addition, each sample was sectioned into seven aliquots before being exposed to repeated freeze–thaw cycles. All sample processing procedures are described clearly in Figure 1. The RNA quality was assessed for its purity and integrity. In this experiment, the A260/280 and A260/230 ratios of RNA 9 from all groups were between 1.8 and 2.1 (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/bio), indicating high purity of RNA. All four types of tissues with no freeze–thaw cycles had RIN values >7. For the liver and muscle tissues, a significant decrease of mean RIN values (to 6.36 ± 0.36 and 5.88 ± 0.54, respectively) after one freeze–thaw cycle at room temperature was detected. Severe degradation occurred after three freeze–thaw cycles at room temperature, as indicated by the mean RIN values of 3.18 ± 0.15 and 3.12 ± 0.12. RNA degradation was also observed when liver and muscle tissues were thawed on ice, with poor RIN values of 4.10 ± 0.58 and 3.44 ± 0.14 after six freeze–thaw cycles on ice (Fig. 2A). Accordingly, the more freeze–thaw cycles the liver and muscle tissues went through, the more smears of RNA fragments that were present (Fig. 2B). However, for brain and testis tissues, the effect of repeated freeze–thaw cycles was not as significant as observed with liver and muscle, displaying mean RIN values of 5.60 ± 0.50 and 7.36 ± 0.96 after three freeze–thaw cycles at room temperature or 6.30 ± 0.13 and 8.32 ± 0.54 after six freeze–thaw cycles on ice (Fig. 2A). Meanwhile, the gel-like images of brain and testis tissue samples showed two clearly distinct bands of 28S and 18S RNA without obvious smearing (Fig. 2B). All of these results indicate that repeated freeze–thaw cycles have detrimental effects on RNA integrity and that this effect varies by tissue type.
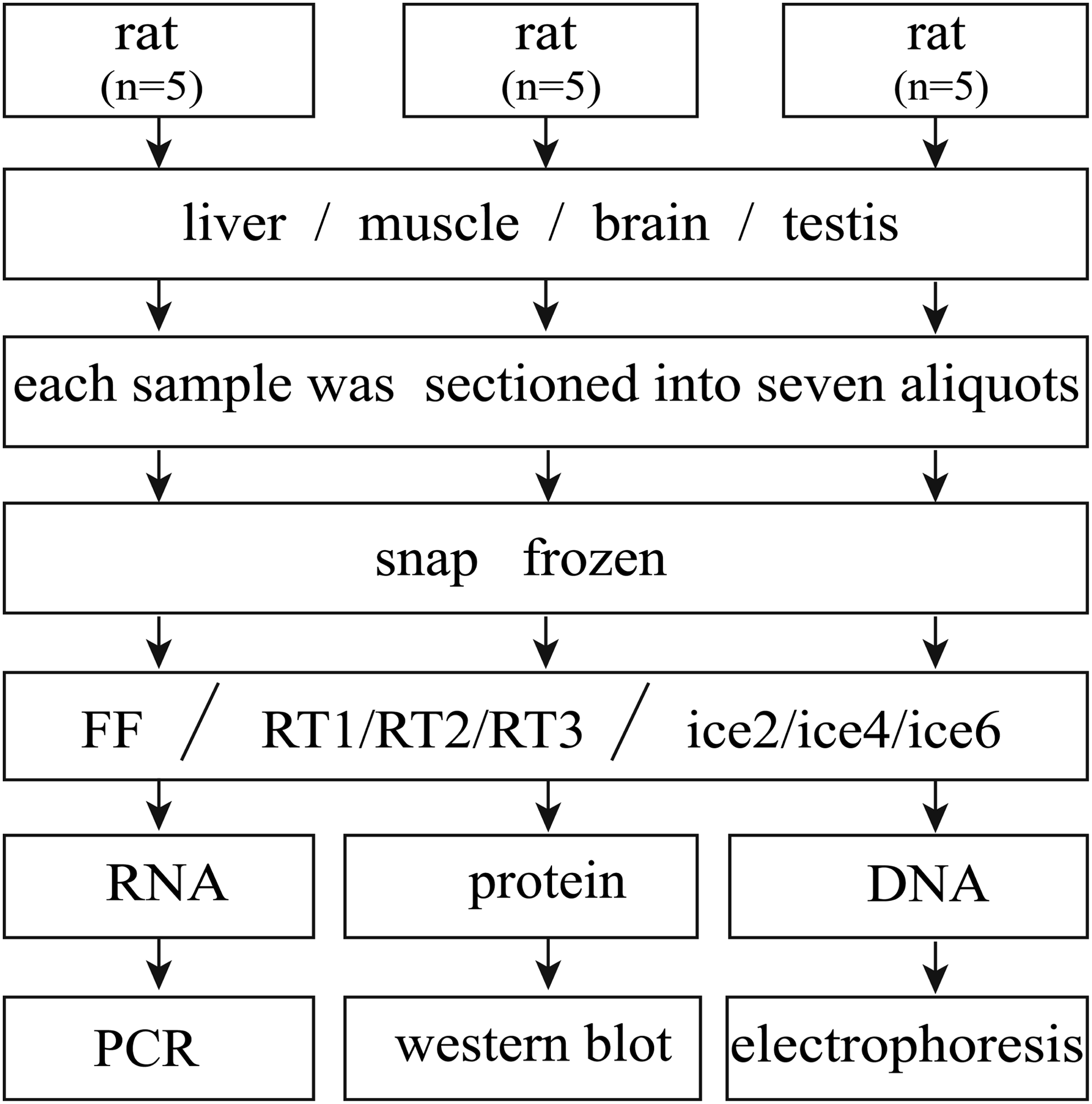
Experimental design. All molecular experiments used five rats. Fresh tissues of liver, muscle, brain, and testis were obtained and each was cut into seven aliquots before being snap-frozen. The first aliquot of each sample experienced 0 cycle FF tissues. Then, three aliquots underwent 1, 2, or 3 freeze–thaw cycles (RT1, RT2, and RT3, respectively) and the thawing process was done at room temperature. The remaining three aliquots of each were exposed to 2, 4, or 6 freeze–thaw cycles (ice2, ice4, and ice6, respectively) and the thawing process was done on ice. Then, the biomolecules of RNA, protein, and DNA were extracted and analyzed for quality. RT, tissues thawed at room temperature; ice, tissues thawed on ice, the number after RT and ice is the freeze–thaw cycle number.

RNA integrity number (RIN) and representative virtual gel-like images of all four types of tissues that had undergone repeated freeze–thaw cycles at room temperature or on ice.
For the same freeze–thaw cycle, RNA integrity was better when tissues were thawed on ice than at room temperature
For all types of tissues, the line graph showed a decline in the tendency of RIN values along with the increased freeze–thaw cycles whether thawed at room temperature or on ice (Fig. 2C–F). For the same freeze–thaw cycle, higher RIN values were obtained from thawing on ice compared with room temperature. Moreover, for liver and muscle tissues, measurements of RNA integrity suggest that there were significant differences between the two thawing methods in the same freeze–thaw cycle, as indicated by the mean RIN values of 5.16 ± 0.59 and 4.08 ± 0.60 after two freeze–thaw cycles at room temperature or 6.78 ± 0.36 and 6.42 ± 0.25 after two freeze–thaw cycles on ice (Fig. 2C, D). This result demonstrates that thawing on ice could protect RNA from degradation compared with thawing at room temperature.
Expression of genes, including the housekeeping gene, was affected in RNA-degraded samples due to repeated freeze–thaw cycles when measured with absolute quantification rather than relative quantification
First, real-time PCR-based relative quantification was applied to evaluate gene expression. Through relative quantification, irregular gene expression alteration was observed along with repeated freeze–thaw cycles. In liver, in which RNA degraded significantly with increased freeze–thaw cycles, gene expression remained relatively stable (Fig. 3A). However, in muscle tissue, in which RNA also degraded significantly, a significant increase of Bcl-2 expression was accompanied by a decrease in Mb expression (Fig. 3B). For the brain and testis samples, most of the gene expression remained relatively stable with increased freeze–thaw cycles (Fig. 3C, D).
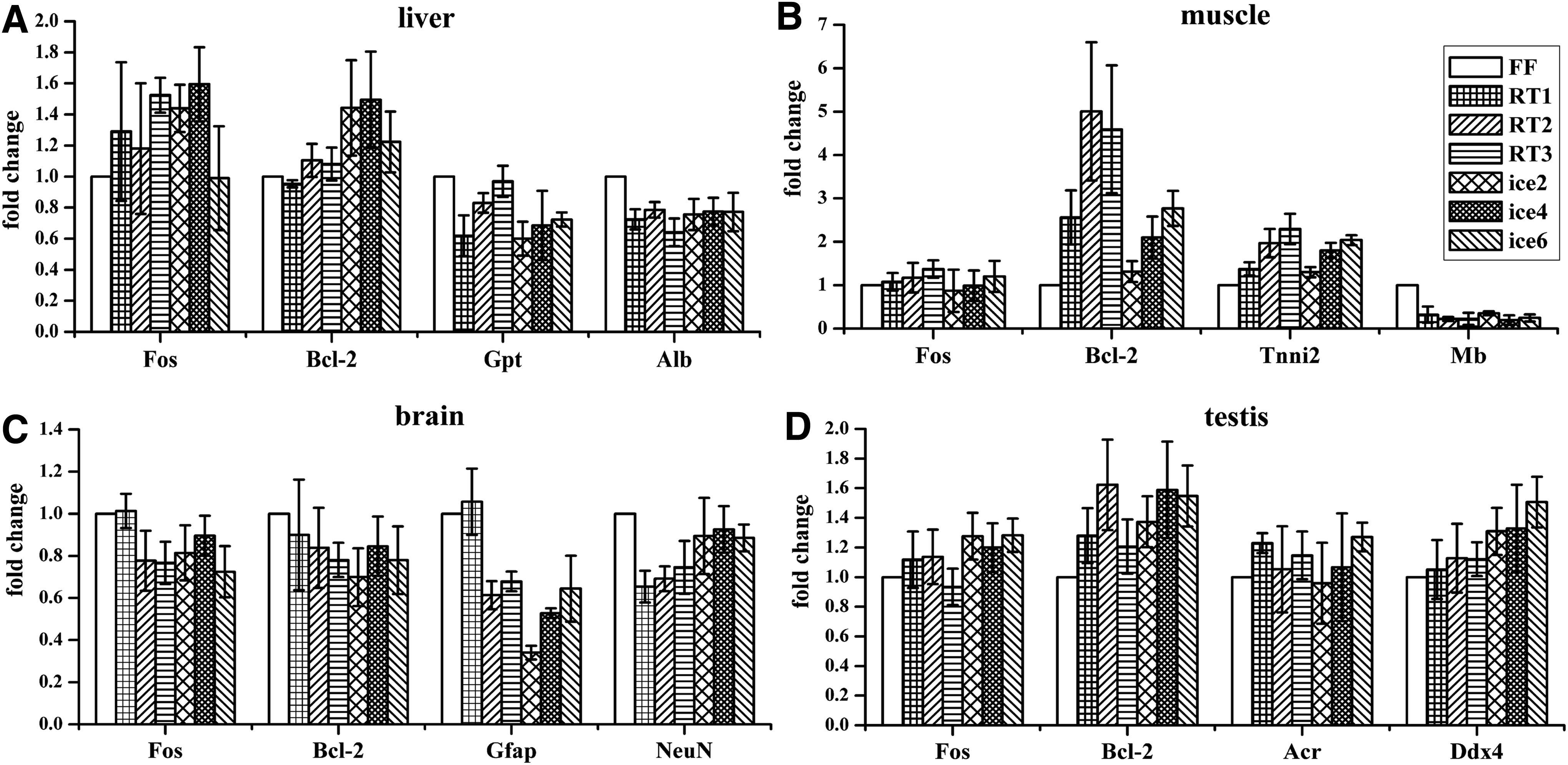
Relative quantitative gene expression detection of all four types of tissues that had undergone repeated freeze–thaw cycles at room temperature or on ice
Second, real-time PCR-based absolute quantification was applied to evaluate gene expression since the housekeeping gene was probably affected by freeze–thaw cycles and thus affected gene expression analysis when it was used for normalization. 10 The copy numbers of the housekeeping gene (Gapdh) were first detected through absolute quantification, which demonstrated a tendency to decline in liver and muscle and remain stable in brain and testis (Fig. 4A). Similar results were observed with copy numbers of FOS, indicating a tendency to decline in liver and muscle and remain stable in brain and testis (Fig. 4B). In addition, the copy numbers of tissue-specific genes in liver and muscle demonstrated a tendency to decline (Fig. 4C, D), whereas tissue-specific genes in brain and testis were relatively stable (Fig. 4E, F).

Absolute quantitative gene expression detection of all four types of tissues that had undergone repeated freeze–thaw cycles at room temperature or on ice.
No obvious degradation of protein and DNA was observed in the four types of tissues that underwent repeated freeze–thaw cycles
Protein quality was evaluated by determining the level of FOS and GAPDH. These results indicated that there was no obvious degradation of FOS and GAPDH in the four types of tissues even after three freeze–thaw cycles at room temperature or six freeze–thaw cycles on ice (Fig. 5).The A260/280 and A260/230 ratios of DNA samples extracted from all groups were between 1.8 and 2.1 (Supplementary Table S2), indicating that high purity DNA samples were obtained. In agarose gel electrophoresis, all of the DNA samples showed high-molecular-weight bands with very few smears, demonstrating the satisfactory integrity of all DNA samples (Fig. 6).
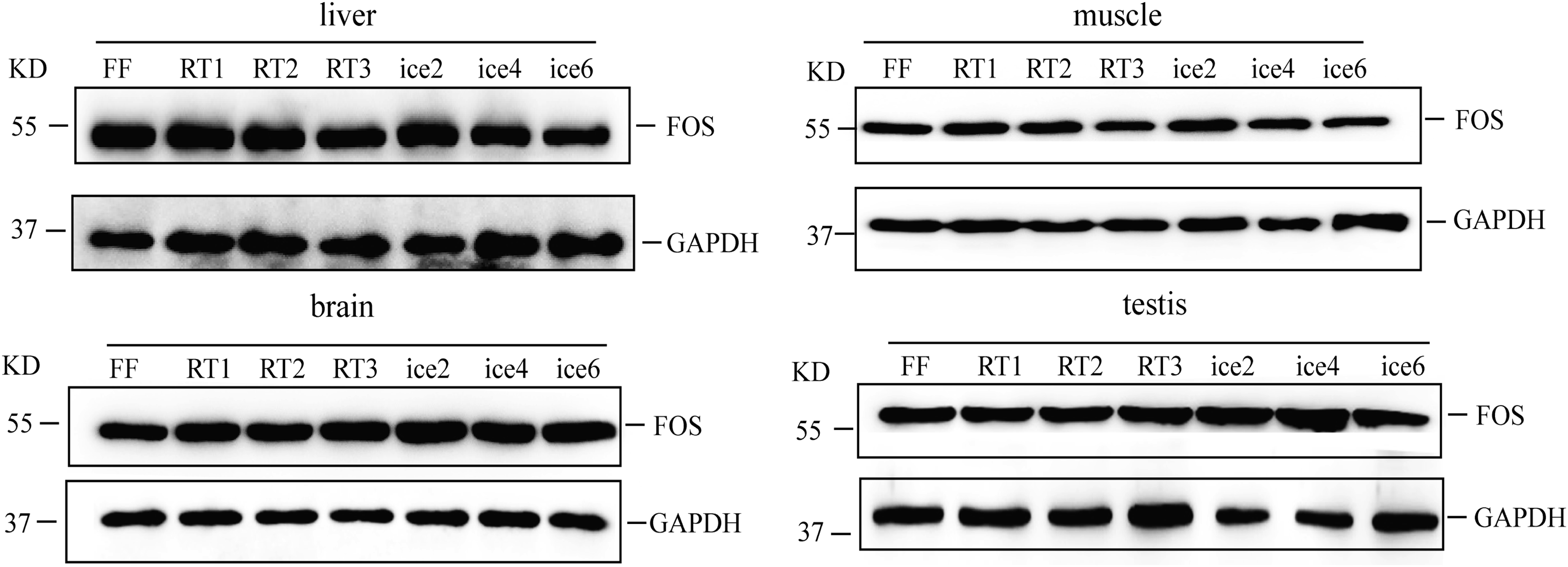
Western blot analysis of all four types of tissues that had undergone repeated freeze–thaw cycles at room temperature or on ice. Protein levels of FOS and GAPDH were detected. FF, fresh frozen tissues; ice, tissues thawed on ice, the number after RT and ice is the freeze–thaw cycle number; RT, tissues thawed at room temperature.
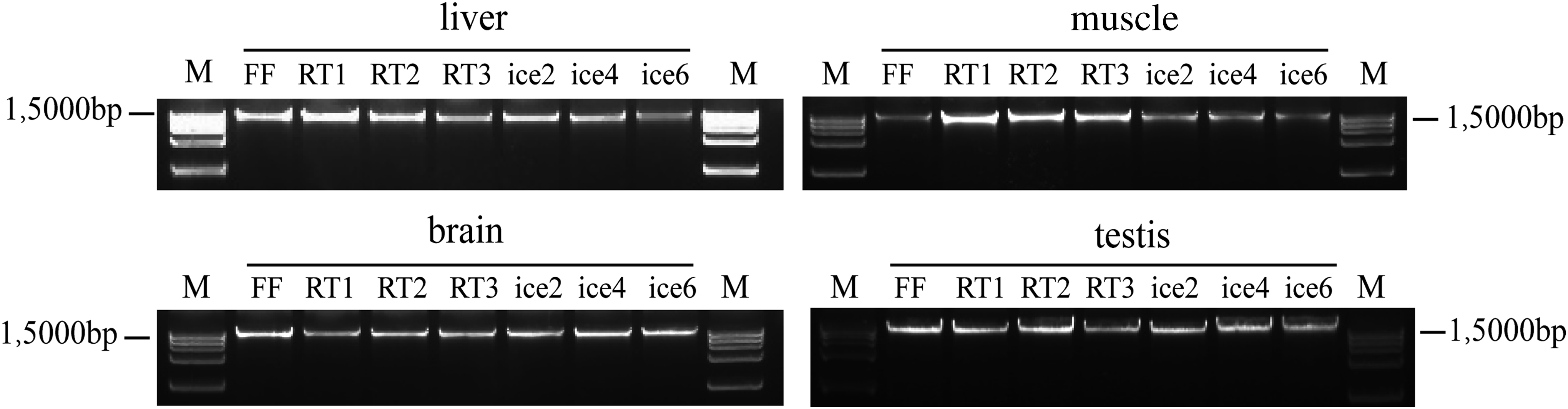
DNA integrity of all four types of tissues that had undergone repeated freeze–thaw cycles at room temperature or on ice were measured by agarose gel electrophoresis. In each case, the lanes were in order of freeze–thaw cycles (FF, RT1, RT2, RT3, ice2, ice4, and ice6, respectively). M represents the DNA ladder. FF, fresh frozen tissues; ice, tissues thawed on ice, the number after RT and ice is the freeze–thaw cycle number; RT, tissues thawed at room temperature.
Discussion
The preservation of high-quality biomolecules in tissue samples is essential for downstream assays. Many sample processing procedures, including the repeated freeze–thaw cycles investigated in this study, could affect the quality of tissue samples. For example, Wang et al. 4 found that the RNA integrity of breast cancer tissue was significantly influenced by repeated freeze–thaw cycles that occurred at room temperature. Hu et al. 11 found that the RNA integrity of gastrointestinal cancer samples and matched adjacent tissues were also severely affected by frequent freeze–thaw cycles at room temperature. However, some researchers reported opposite results. Jochumsen et al. 12 found that the RNA integrity of ovarian tumor tissue was not impacted by three freeze–thaw cycles on cold marble. Additionally, Franca et al. 8 demonstrated that the RNA integrity of bovine reproductive tissue remained intact after six freeze–thaw cycles on ice. These inconsistencies were probably due to the different tissue types selected and the thawing procedure used by different research groups. In our experiments, we first chose four different tissue types for our investigation of the tissue-specific variable effects of repeated freeze–thaw cycles. We then performed experiments comparing room temperature and ice thawing procedures in the same tissue type. Finally, 30 minutes was chosen as the thawing period because some researchers had observed that 30 minutes of thawing time is acceptable for tissues lacking any protective reagent. 7
The kinetics of RNA degradation after repeated freeze–thaw cycles was likely to vary by tissue type. For liver and muscle tissues, RNA degraded significantly with each repeated freeze–thaw cycle regardless of the temperature at which the sample was thawed. The results were consistent with previous studies that demonstrated that liver and muscle tissues contain high activity of RNase, resulting in significant RNA degradation and large variation in RIN values under numerous conditions. 13 For testis tissue, RNA was demonstrated to be stable through repeated freeze–thaw cycles in this study, which is in accordance with previous research by Fitzpatrick et al. 14 who had reported that a delay of up to 24 hours did not impact the integrity of RNA extracted from bovine reproductive tissue. Franca et al. 8 further confirmed this result. There seems to be some controversy over the effects of freeze–thaw cycles on brain samples. Many researchers have speculated that the brain was a metabolically active organ and that the thawing and refreezing process could be injurious to RNA because of the damage done to cellular membranes and the release of RNase. 15 For example, Castells et al. 16 found a high percentage of RNA degradation in a large dataset of brain tumor samples. Sherwood et al. 17 also observed that unfixed, snap-frozen brain samples were generally of poor RNA quality. However, Johnson et al. 18 found that in humans and rats, RNA extracted from brain tissue seemed to be particularly stable over time. Beach et al. 19 observed that RIN values of brain tissue underwent repeated freeze–thaw cycles at room temperature with a small decrease of no statistical significance. In our study, the brain sample remained relatively stable with increased freeze–thaw cycles compared with liver and muscle tissues. However, the mechanism underlying this tissue's variation needs to be further explored in our future studies.
Although repeated freeze–thaw cycles could affect RNA integrity, how the process influences gene expression needs to be investigated. First, relative quantification was applied to analyze gene expression as it is a commonly used method with high sensitivity and good reproducibility. 20 Results indicated that irregular gene expression alteration and even increase of gene expression were observed with increased freeze–thaw cycles. Considering that the housekeeping gene used for normalization could be affected by RNA degradation10,21 and therefore impact gene expression results, absolute quantification was applied to evaluate gene expression as it is a powerful method to detect minimal copy number changes of the target gene without been normalized to a housekeeping gene. 22 Our results indicate that the copy numbers of Gapdh through absolute quantification declined with RNA degradation in the liver and muscle, which could probably explain their irregular gene expression alteration when it was used as the housekeeping gene in relative quantification experiments. The tendency for declining copy numbers of Fos and tissue-specific genes through absolute quantification was also observed, which was in accordance with the changing tendency of RIN values. The results of absolute quantification reflected that expression levels of all genes, including the housekeeping gene, were influenced in RNA-degraded samples caused by repeated freeze–thaw cycles, indicating that using the housekeeping gene for normalization in relative quantification might bias the experimental results in such circumstances.
The proteins of GAPDH and FOS, which were both highly expressed and commonly used in protein experiments, were chosen to evaluate the protein quality. Although no obvious influence of repeated freeze–thaw cycles on proteins of GAPDH and FOS was detected through western blot in this study, we suggest a more global proteome approach to detect the detailed alteration of the entire set of proteins or tissue-specific proteins in specific research projects. For detection of DNA integrity, results indicated no obvious DNA degradation through gel electrophoresis, which is considered to be the most commonly applied method to detect DNA degradation. 23
Temperature has a significant influence on the quality of tissue samples throughout the transportation, storage, and thawing processes. 24 Considering that most biomolecules are temperature sensitive,25,26 tissue samples thawed at a lower temperature could probably be better protected from enzyme degradation. In our preliminary experiment, the results indicated that under the same freeze–thaw cycle condition, RIN values were higher when thawed on ice than that at room temperature. To make the comparison more important and valid, more common freeze–thaw cycles should be compared between thaw-on ice and thaw-at room temperature. However, due to the limited volume of testis tissue, we had to pick some key points to evaluate the change of RIN values. We calculated the same thawing interval at different thawing temperatures from the samples being taken out of the freezer, as most researchers have done in their studies.4,11,27As we expected, when tissues were thawed on ice, the speed of RNA degradation could be slowed down and RNA integrity protected. We reason that a lower temperature environment could probably reduce the RNase metabolic rate, thereby protecting the RNA from degradation. However, it is technically difficult to accurately calculate the thawing rate and detect the time point when the tissue is completely thawed. Future research could focus on a more detailed investigation of RNA degradation once it is technically possible to detect the thorough thaw state of a tissue. In addition, a new CryoXtract technology was recently applied to section frozen biosamples into aliquots at −160°C. 28 We hope that it could be widely applied in the biobank platform to avoid repeated freeze–thaw cycles and prevent the RNA from degradation.
In conclusion, repeated freeze–thaw cycles could influence RNA integrity and gene expression. For new tissue samples to be analyzed, it is generally suggested in the current standard operation procedure that it should be sectioned into small aliquots before snap-freezing to avoid RNA degradation and gene expression alteration. For the old valuable tissue samples, which are already in storage and need to be used repeatedly during their lifecycle, thawing on ice or being sectioned at ultralow temperature is recommended.
Footnotes
Acknowledgments
This study was supported by funding from the National Nature Science Foundation of China (81270742, 81370700); funding from Shanghai Municipal Commission of Health and Family Planning (201540389); Professional and Technical Services Platform for Biobank of Critical Disease in Shanghai (15DZ2292100); and Interdisciplinary Funding of Medical and Engineering from Shanghai Jiao Tong University (YG2016MS32).
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.