
Abstract
Successful cryopreservation of avian gonads is important not only for avian breeding but is also crucial for preservation of species, especially of endangered birds. The aim of this study was to evaluate the effect of vitrification by several cryoprotectants on the ovarian tissues of laying hens. Ovarian tissues were randomly divided into six groups: control (nonvitrified: C), dehydrated using ethylene glycol (EG), dehydrated with propylene glycol (PROH), dehydrated using dimethyl sulfoxide (DMSO), and two combined groups, EG+DMSO and EG+PROH. The composition of vitrification solutions was as follows: EG group: V1 = 7.5% EG and V2 = 15% EG +0.5 M sucrose, DMSO group: V1 = 7.5% DMSO and V2 = 15% DMSO +0.5 M sucrose, PROH group: V1 = 7.5% PROH and V2 = 15% PROH +0.5 M sucrose, EG+DMSO group: V1 = 7.5% EG +7.5% DMSO and V2 = 15% EG +15% DMSO +0.5 M sucrose and EG+PROH group: V1 = 7.5% EG +7.5% PROH and V2 = 15% EG +15% PROH +0.5 M sucrose. Ovarian tissues of each group were dehydrated for 10 minutes with V1 solution and 2 minutes with V2. Among the vitrified groups, intact primordial and primary follicles showed significant increase in EG+DMSO, but follicular attrition had the highest rate in the PROH group (p < 0.05). Immunohistochemical analysis showed that the percentage of active caspase 3-positive cells was lower (p < 0.05) when using EG+DMSO versus PROH. Further gene expression of caspase 3, 8, and 9 was highest in the PROH group (p < 0.05). Vitrification of ovaries of laying hens using EG+DMSO can afford effective protection of primordial and primary follicles during preservation and may therefore be successfully used for storing avian gonadal tissues.
Introduction
O
The use of freezing methods helps facilitate this process by allowing access to the genome. Thus, recent years have seen a rise in research involving the use of freezing techniques to cryopreserve primordial germ cells. An important issue in cryopreservation is identifying the best kind of cryoprotectant that leads to minimal damage to the tissue for each animal species. 13 Thus, numerous studies compare various cryoprotectants in human, 14 mouse, 15 and rat 16 ovaries.
Among several cryoprotectants, propylene glycol (PROH), dimethyl sulfoxide (DMSO), and ethylene glycol (EG) are viable options and researchers have commonly used them due to their efficiency of cryopreserving reproductive organs. 17 While DMSO (C2H6OS) has a molecular weight and density of 78.13 g/mol and 1.1004 g/cm3, respectively, EG (C2H6O2) has the least molecular weight and is the most dense (1.1132 g/cm3). PROH (1,2-propanediol, C3H8O2), which has a molecular weight of 76.06 g/mol and density of 1.0597 g/cm3, is less penetrable than EG. EG is more penetrable than other cryoprotectants, 18 but the size and imbalance density of cells specifically in the cells of ovarian tissue change the effect of this penetration. 19 The effects of cryoprotectants vary according to the type of substance and animal species. 19 For instance, DMSO is a secure cryoprotectant for several species such as mouse oocytes, 20 bovine oocytes, 21 and human germinal vesicle oocytes, 22 but in some species such as zebra fish it is poisonous. 23
Using the adult laying hen as a model, it may be that results can be extended to cryopreservation of ovarian tissues of other avian species. The single ovary of the chicken is irregularly shaped. This organ is attached to the dorsal surface of the left side of the abdominal cavity wall. 24 Follicles of the ovary have different sizes, and usually an ovary contains 5–10 follicles greater than 1 cm in diameter. Primary oocytes are enclosed by the vitelline membrane. These oocytes become organized into a primordial follicle (up to ∼80 μm in diameter). The average diameter of primary follicles is 0.8 to 1 mm and ovulation occurs in the largest follicle. The accumulation of lipoprotein-rich, white yolk and differentiation of the theca into internal and external layers occur and further growth leads to the prehierarchical follicle stage (1–8 mm). 25
As cryoprotectants are cytotoxic, 26 the concentration of exposure to cryoprotectants should be selected carefully, because it is a key factor to minimize the toxic effects. Meanwhile, there are a few reports about quantitative assessment of follicles at different categories immediately after vitrification warming of laying hen ovarian tissue. For this purpose, in the present study, we used adult hen ovaries, rather than the small, juvenile ovaries used by the other studies. Some previous studies have illustrated that ovarian tissue cryopreservation of 1-week-old Japanese quail has successfully been done. 12 More studies on avian gonadal transplantation 27 have an important role in improving the functional recovery of cryopreserved gonadal tissue.
Although the vitrification of avian ovarian tissue seems to be well established, 28 a few studies report differences in the sensibility of types of follicles (primordial, primary, and prehierarchical) to a variety of cryoprotectants and gene expression mechanisms regarding vitrification of the laying hen ovary. Furthermore, numerous studies have confirmed that vitrification could result in the emergence of apoptosis in vitrified ovaries and embryos and change the epiphany of apoptosis-related genes.29,30 The objective of the current investigation was to assess the efficiency of vitrification solution, which contains different cryoprotectants (EG, DMSO, and PROH), for cryopreservation of mature laying hen ovaries, and quantitative and qualitative assessment of different follicular categories after vitrification. Furthermore, gene expression (Bak, Bcl-2, Bak/Bcl-2, Cas3, Cas8, and Cas9) of ovarian tissue from adult hen ovaries after vitrification was compared to noncryopreserved control tissue.
Materials and Methods
Birds, tissue preparation, and study design
Seven White Leghorn hens (6 months old) were handled in compliance with existing guidelines and subject to the approval of Royan Institute Ethics Committee (IR.ACECR.ROYAN.REC.1394.30). Laying hens were euthanized by cervical dislocation and ovarian tissues were removed from the body, immediately transferred to the laboratory in phosphate-buffered saline (PBS), and washed three times in fresh PBS. Follicles larger than 3 mm were separated from the ovarian tissue. Then, the ovarian tissues were randomly distributed into six experimental groups: nonvitrified control (C), dehydrated by using EG, dehydrated with PROH, dehydrated with DMSO, and two combined groups: EG+DMSO and EG+PROH. The size of each piece of ovarian tissue was ∼3 × 3 × 2 mm. The experiments were followed by morphological evaluations, follicular counts, and gene expression studies (Bak, Bcl-2, Cas3, Cas8, and Cas9) (Fig. 1).
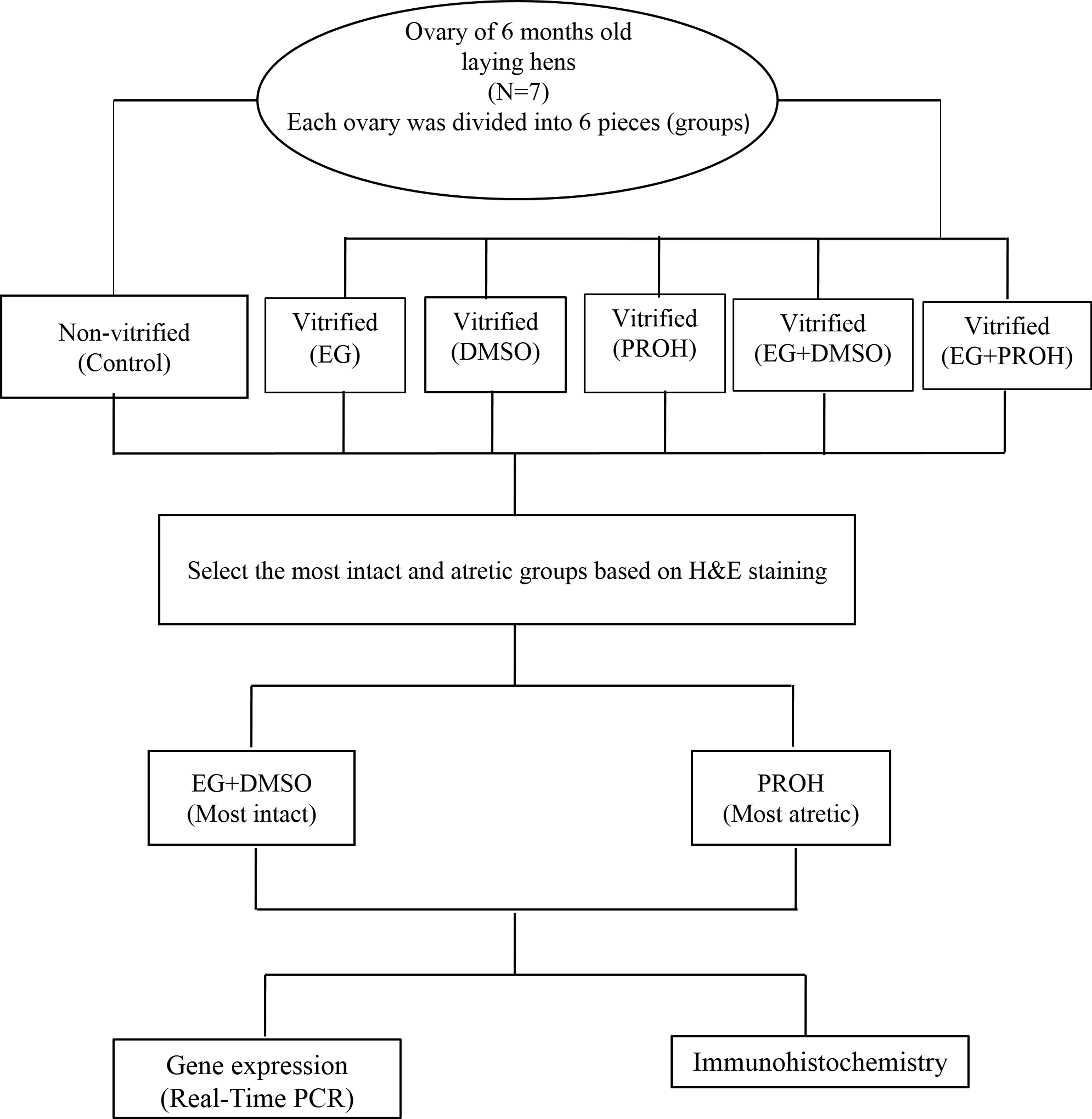
Outline of the experimental procedure.
Vitrification procedure (dehydration and plunging of samples into liquid nitrogen)
The vitrification procedure was performed according to Liu et al., 12 with a minor modification. Base medium (BM) in equilibrium (V1) and vitrification (V2) solutions were composed of Dulbecco's modified Eagle's medium (Gibco, Invitrogen, Carlsbad, CA) and 10% fetal bovine serum (FBS) (Gibco, Grand Island, NY) (Table 1). First, all the ovarian tissues from vitrified groups were dehydrated through immersing in V1 and V2 solutions for 10 and 2 minutes, respectively, at room temperature (RT), and the tissue samples were picked up with the needle of an insulin syringe (cryopin) 31 and immersed in liquid nitrogen. Dehydrated ovarian tissues were placed in cryovials and stored in a nitrogen tank for more than 3 weeks.
DMSO, dimethyl sulfoxide; EG, ethylene glycol; FBS, fetal bovine serum; PROH, propanediol.
Warming procedure (rehydration and reviving the vitrified samples)
The warming procedure was carried out as described by Liu et al., 12 with some modifications from our pretests. In this study, there were no acceptable results regarding warming for 5 minutes according to Liu's study. Therefore, 1 and 3 minutes of warming were also examined and histological evaluations showed that the optimum time for warming is 1 minute.
The BM in warming solutions was composed of Dulbecco's modified Eagle's medium and 10% FBS. During the warming process, first the vitrified ovaries were transferred from the liquid nitrogen tank to W1 solution (BM+sucrose 1 M) and kept for 1 minute. Then, the tissues were transferred from W1 solution to W2 solution (BM+sucrose 0.5 M) and then to W3 solution (BM+sucrose 0.25 M) and kept in each solution for 1 minute. The entire washing process was performed at RT. Eventually, to complete the warming process, the washed ovaries were transferred to W4 solution (BM) and incubated in a 37°C, 5% CO2 humidified incubator (New Brunswick Scientific Co., Inc., Edison, NJ) for 30 minutes until histological assessments were made.
Morphology and follicle structure
The ovary samples were fixed at RT in Bouin's fluid and formalin for 24 and 72 hours, respectively. Then, they were dehydrated in alcohol and embedded in paraffin wax. Thick sections (5 μm) were cut and stained with hematoxylin and eosin (H&E) for histopathological studies.
In normal follicles, the layer of granulosa cells is attached to the spherical oocyte surrounding it. Furthermore, the homogenous ooplasm (cytoplasm of the oocyte) contains a tiny granulated nucleus. In contrast to normal follicles, in atretic follicles, aggregation and shrinkage of nuclear chromatin and wrinkling of the nuclear membrane were regarded as signs of atresia. To avoid double counting, only the follicles with a visible nucleus were counted. Determination of the different types of follicles was done according to a study by Johnson and Woods 25 (Fig. 2). Accordingly, follicles with a maximum size of 80 μm were counted as primordial follicles and those 0.08–1 mm were considered primary follicles. Also, follicles with a size greater than 1 mm were considered prehierarchical follicles. The best preserved group (EG+DMSO) and the worst group (PROH) as determined by morphological evaluations were selected for immunohistochemical and gene expression assays.

Vitrified warmed laying hen ovarian tissues. Atretic primordial follicles (white arrows) and intact primordial follicles (dash arrows). Atretic primary follicles (arrowhead) and intact primary follicles (black arrows). Intact prehierarchical follicles (star). Control (nonvitrified)
Caspase 3-positive follicles detected by immunohistochemistry
For immunohistochemistry, the section of each tissue was randomly selected and stained with active caspase 3 antibody. To suppress endogenous peroxidase activity, tissues were treated with 3% H2O2 in methanol for 30 minutes. 32 Then, the sections were soaked in citrate buffer (pH = 6.0) and warmed up to 98°C for 1 hour. After that, tissue sections were blocked for 1 hour with protein block and incubated at 4°C overnight in 1:200 dilution of active caspase 3 antibody (ab4051). They were washed well and incubated with a secondary antibody (ab97051) for 1 hour at RT. Eventually, the blocks were washed and the attached antibody was visible after the addition of 3, 3-diaminobenzidine tetrachloride (DAB) solution. For accurate cell counting by light microscopy, counterstaining of slides with hematoxylin was carried out. 32 In this study, follicles in which oocytes or more than 50% of granulosa cells showed positive reaction for caspase 3 were considered apoptotic follicles (Fig. 3). 31

Immunohistochemical staining for caspase 3 in vitrified warmed laying hen ovarian sections. Thymus (negative control)
Quantitative real-time polymerase chain reaction for apoptotic genes
Soon after warming, fragments from each group were stored in RNAlater reagent (Ambion) and preserved at −80°C until RNA extraction. Total RNA was extracted from the samples through a manual method using TRIzol (Sigma-Aldrich) as follows: small ovarian fragments were added to TRIzol and homogenized. The fragments were incubated on ice for 5 minutes and centrifuged (12,000 rpm, 15 minutes, 4°C). Then, chloroform (Merck, Germany) was added followed by cold isopropanol (Merck, Germany). The mixture was incubated for 60 minutes at −20°C and then centrifuged (12,000 rpm, 15 minutes, 4°C). Finally, the RNA pellets were washed with 70% ethanol, air-dried, and dissolved in diethyl pyrocarbonate-treated water. Evaluations of concentration and purity of the total RNA were done using a spectrophotometer at 260/280 nm.
A cDNA Synthesis Kit (Fermentas, Leon-Rot, Germany) and random hexamers were used to synthesize complementary DNA (Table 2). Gene expression levels were measured using Applied Biosystems (ABI 7500 thermocycler) Step One and Step One Plus real-time PCR systems. Expression levels of Bak, Bcl-2, Cas3, Cas8, and Cas9 were evaluated. The housekeeping gene GAPDH was used for normalization. Design of specific primers was done using the software Allele ID (Premier Biosoft, Palo Alto, CA) (Table 2).
Statistical analysis
Statistical analysis was carried out using SPSS 22.0 (IBM Crop., Armonk, NY). Continuous variables were expressed as mean ± standard error of mean. The Kolmogorov–Smirnov test was used to evaluate the normality of the data. A one-way ANOVA, followed by the Duncan posthoc test, was used to compare the groups. All statistical tests were two sided and a p-value <0.05 was considered statistically significant.
Results
Morphological evaluations (intact, atretic, and apoptotic follicles)
The effects of different cryoprotectants on the follicle count of hen ovarian tissues were compared between vitrified and nonvitrified groups (Figs. 4 and 5). The proportion of intact primordial and primary follicles in EG+DMSO group was significantly higher than other vitrified groups and it was the lowest in PROH group (p < 0.05; Fig. 4). The number of intact primordial, primary, and prehierarchical follicles in all vitrified groups was significantly decreased compared to the control group (p < 0.05; Fig. 4). The use of EG+DMSO compared to other cryoprotectants significantly contributed to the number of primordial and primary follicles, but these changes were less pronounced in prehierarchical follicles. Therefore, EG and combination of EG and DMSO did not alter the prehierarchical follicle number significantly.

The percentage of intact primordial, primary, and prehierarchical follicles in the control and vitrified groups. Values are given as mean ± SE. Groups followed by the same letter are not significantly different at p < 0.05.
Immunostaining results showed that caspase 3-positive primordial and primary follicles were significantly lower after using EG+DMSO vs. PROH (p < 0.05; Fig. 5). Apoptotic primordial and primary follicles were increased in the ovaries treated with PROH compared to both EG+DMSO and control groups (p < 0.05; Fig. 5). There was no significant difference in the number of apoptotic prehierarchical follicles between the EG+DMSO group and the nonvitrified control group.

The percentage of apoptotic follicles (caspase 3 positive) in laying hen ovarian tissue. Nonvitrified (control), EG+DMSO, and PROH. Values are given as mean ± SE. Groups followed by the same letter are not significantly different at p < 0.05.
Gene expression
The expression of Bak was significantly higher in PROH group than the other groups (p < 0.05; Fig. 6). No significant difference was observed between EG+DMSO and nonvitrified groups. The analysis of Bcl-2 gene expression indicated a considerable downregulation in EG+DMSO and also PROH groups with regard to the control group (p < 0.05), with a clear change between EG+DMSO and PROH groups. Vitrified groups did not differ considerably in apoptotic index (Bak/Bcl-2). The expression levels followed a similar pattern in all groups for caspase 3, 8, and 9 genes (Fig. 6). The expression of all three caspases (3, 8, and 9) was the highest (p < 0.05) in the PROH group compared to EG+DMSO and control groups. This variable was also significantly higher in the EG+DMSO group against the control specimens (p < 0.05; Fig. 6).
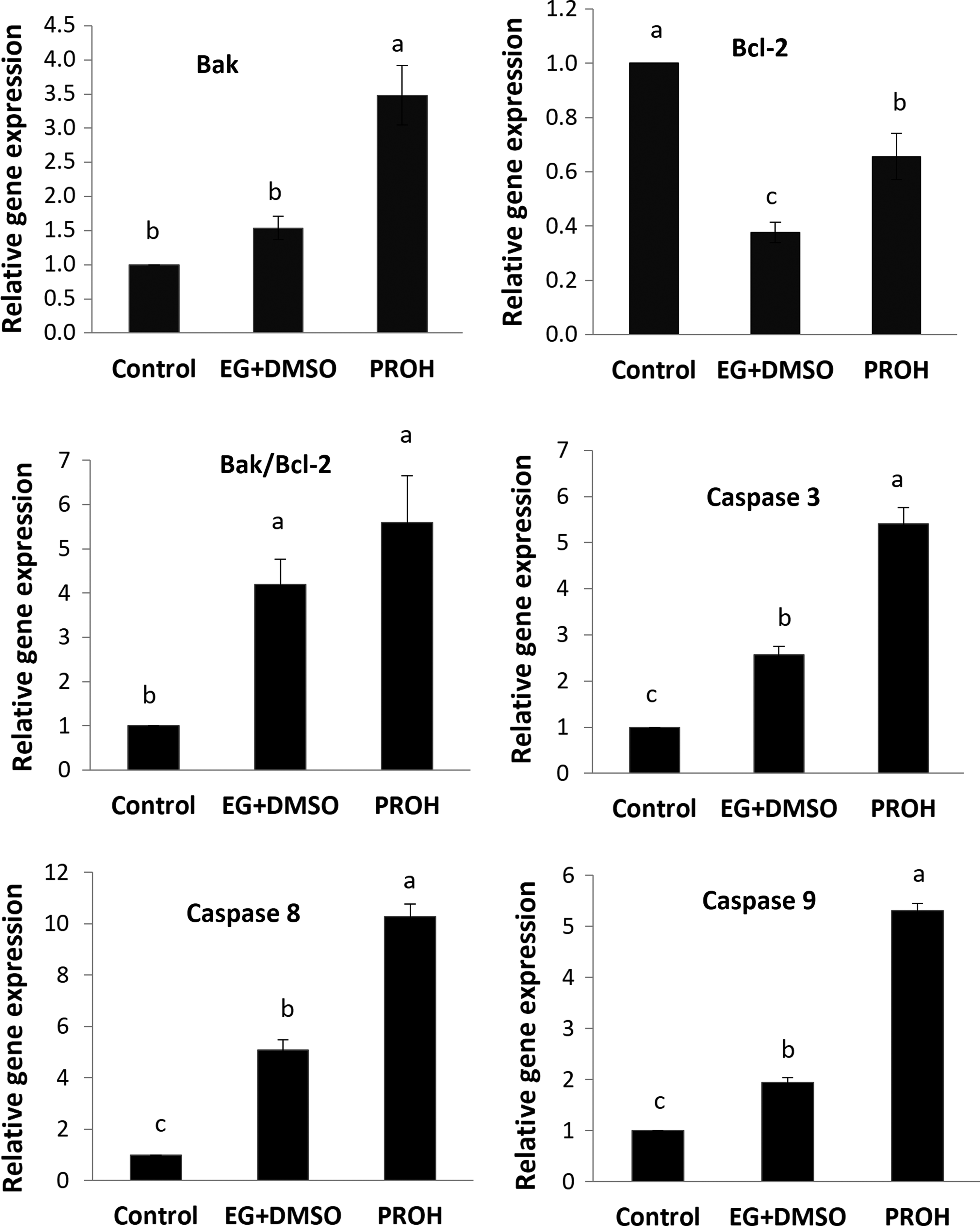
Relative gene expression of Bak, Bcl-2, Bak/Bcl-2, caspase 3, caspase 8, and caspase 9 in laying hen ovarian tissue. Nonvitrified (control), EG+DMSO, and PROH. Values are given as mean ± SE. Groups followed by the same letter are not significantly different at p < 0.05.
Discussion
Ovarian tissue cryopreservation would be a very useful tool for the poultry industry and the successful conservation of the poultry genetic reservoirs that still exist. We can store the current generation of superior line breeding stock as conservation against natural disasters and insurance against inbreeding and unintended changes in breeding process. 2 The key component in desirable vitrification is cryoprotectant. Ice crystals damage cells, so high concentrations of cryoprotectants inside cells under ultracooling situations become hyaloid to prevent their formation.33,34 The cryoprotectants cause enhancement in viscosity, as a result, the solution does not crystallize but becomes amorphous ice. 33 Although methods for vitrification of juvenile ovaries have been established, 28 cryopreservation of adult avian ovaries is challenging.
Five vitrification protocols were used for the vitrification of laying hen ovarian tissue. Considerable differences were reported among the cryoprotectant agents used, but composition of EG+DMSO was the best adapted for hen ovarian tissue cryopreservation and formed a greater conservation against freezing than other vitrified groups. This approach was confirmed by previous reports on quail, 35 rat, 36 bovine, 7 and caprine 10 ovarian tissues.
It has been observed that the composition of cryoprotectants makes less toxic effects and proved more appropriate than independent cryoprotectants. 37 There is another report that suggests cryoprotectant mixtures might induce better results compared to the media containing one penetrable cryoprotectant. 38 Investigators have reported that the combination of EG and DMSO has successfully been used in vitrification of human embryos. 39 Also, numerous studies have shown that EG was selected as a basic penetrable cryoprotectant because it has relatively low toxicity and very low molecular weight. 40 Another research reported the high rates of survival in mature or immature mouse oocytes following vitrification using EG. 41 Moreover, it was shown that when 2–4 cell mouse embryos were cryopreserved using EG, the blastocyst formation, survival, and blastocyst hatching rates are considerably high compared to PROH, DMSO as cryoprotectant. 42
DMSO is a permeating cryoprotectant, which protects cells from intracellular ice crystal formation when used at higher concentrations. 43 Some researchers have reported EG is better than DMSO, 9 while other results contradict this claim. 19 It was suggested that EG is the most effective cryoprotectant in vitrification, 44 however, PROH is the most efficient cryoprotectant in slow-freezing. 45 In our study, propanediol damaged the ovarian tissue and provided less conservation during vitrification. This may be due to applying propanediol alone, because PROH and EG combination increased the rate of follicular preservation.
The results of a recent study indicated that PROH group has the lowest oocyte survival rate, but in the group of propanediol and glycerin mixture, the survival rate of oocytes was significantly higher than other vitrification groups. 34 In addition, similar results 46 showed higher implantation and clinical pregnancy rates of human 2–4 cell embryos in EG group compared to those in PROH group. Indeed, investigation on calf testicular tissue within vitrification reported that cell viability in the group treated with 10% DMSO was more than the groups treated with 10% PROH and 7.5% glycerol. 47 However, the selection of a suitable protocol for ovarian tissue vitrification depends on histological ovarian structure, number of follicles, and the permeability of different cryoprotectants.
The comparison of the number of follicles between different groups showed that EG+DMSO preserved the largest number of intact primordial and primary follicles compared to other vitrified groups. The smallest number of intact primordial and primary follicles was observed in the PROH-treated group. Although we could not show considerable difference between the EG+DMSO and EG group in the number of prehierarchical follicles, this difference was dramatically considerable between the EG+DMSO and PROH group. Generally, primordial and primary follicles were the most damaged among all vitrified groups, and the prehierarchical follicles following vitrification had the lowest rate of damage. So, this result indicated that the prehierarchical follicles are more resistant against other types of follicles and less susceptible to various cryoprotectants.
It is contrary to the results of some studies stating that the large ovarian follicles are more sensitive and expected to cryoinjury compared to smaller follicles such as primordials because of their larger size and fat volume and higher active metabolism. 19 This may be attributed to the anatomical differences between the ovary of the hen and that of other species, including humans. The human ovarian surface is smooth and the cortex under the surface epithelium surrounds the primordial and primary follicles. 48 Studies have shown that the cholesterol in egg yolk can replace the missing cholesterol of cell membranes during vitrification, similar to natural plasma membrane reconstruction, 49 which protects follicles during vitrification. Besides, it has been shown that egg yolk is a key component that protects the integrity of the cumulus complex. Preservation of the cumulus complex may arise from the protection, by the plasma membrane of oocyte, of the junctions of the cumulus cell processes. 41 This may be the reason for better preservation of larger follicles in our study.
However, in this study, occurrence of follicular atresia following vitrification/warming of ovarian tissue was significantly higher than in nonvitrified tissue, reflecting damage to the follicles during the vitrification/warming procedure. This finding is in agreement with the study on vitrification of human ovarian tissues. 50 In contrast, a few other studies have observed no significant differences in the percentage of intact follicles between the vitrified and nonvitrified tissues. 10 This discrepancy may be attributed to the technique of cryopreservation and type of cryoprotectants. In addition, these changes require different criteria for evaluation of morphological abnormalities, and H&E protocols in different studies have been distinct due to the same reason.
For the first time, the expression of some apoptosis-related genes in vitrified laying hen ovarian tissue was measured. Ovarian tissue apoptosis occurs due to two mechanisms 51 : the intrinsic (mitochondrial) pathway of apoptosis that occurs by means of signals generated from Bcl-2 family members and the mitochondria, and the extrinsic (cell death signaling) pathway induced by the attachment of death molecules to the Fas/FasL system (as one of the cell surface receptors). Generally, in apoptosis, caspase (cysteine-dependent aspartate-directed proteases) has a determinative role. Activation of the caspase cascade results in cleavage and disintegration of substrate proteins and subsequent removal of the degenerating cell. 52 The Bcl-2 family is known for its regulation of apoptosis. This family of proteins consists of members that either promote (Bax and Bak) or inhibit (Bcl-2) apoptosis and control apoptosis by governing mitochondrial outer membrane permeabilization. In addition, cytochrome c binds to cardiolipin in the inner mitochondrial membrane, anchoring its presence and keeping it from being released from the mitochondria and initiating apoptosis (by the intrinsic pathway). 53
Our findings indicated that Bak was strongly expressed in the PROH group when compared to control and EG+DMSO groups. The Bcl-2 expression was downregulated in both vitrification groups, but was higher in PROH group than EG+DMSO group. Based on our knowledge, there are no reports on cell death assessment at the molecular level following cryopreservation of laying hen ovaries. Nevertheless, Turathum et al. 54 and Kuzmany et al. 55 reported that expression of Bax was raised in response to vitrification in bovine blastocysts, the same increase was seen in the present study. In contrast, it was shown that vitrification does not have any effect on expression of Bax and Bcl-2 in sheep oocytes 56 and mouse embryos. 30 Indeed, there are some reports that suggest when Bcl-2 is in excess, the cells are preserved and when Bax is in excess, the cells are susceptible to apoptosis. 57 However, coincident with our results, some investigations have reported that the expression level of Bcl-2 gene raises after vitrification. 58 However, our results regarding the expression of the Bcl-2 gene were unexpected. It was expected that the expression rate of Bcl-2 in the group treated by propanediol would be less than EG and DMSO mixture. This can be due to a compensatory mechanism preventing stress induced with cell apoptosis. 59
Caspase, a family of cysteine proteases, is a key factor in mediating apoptosis. Major caspases identified so far have been classified into initiators (caspase 2, 8, 9, and 10) and executioners or effectors (caspase 3, 6, and 7). 60 In this investigation, caspase 3, 8, and 9 were assessed. The results derived from this study represent that the expression levels of all three caspases (3, 8, and 9) were the highest in PROH group than in EG+DMSO and control groups. This variable was also considerably higher in EG+DMSO group compared to the control samples. In this regard, the study 61 reported that caspase 8 expression as an initiator was raised in vitrified ovarian tissue specimens, but it was not similar in caspase 3 cases.
Conclusion
Our findings show that applying the cryoprotectant PROH alone induces apoptosis to a greater extent than the EG+DMSO combination. In addition, in terms of apoptosis incidence rate and the lowest number of atretic follicles, EG+DMSO appears to be the suitable composition among cryoprotectants for preservation of follicles within avian ovaries, and this warrants further studies.
Footnotes
Acknowledgment
The research was scientifically and financially supported by grants from Royan Institute and Gorgan University of Agricultural Science and Natural Resource.
Author Disclosure Statement
No conflicting financial interests exist.