
Abstract
Although the Ficoll-Paque method is classically used to isolate peripheral blood mononuclear cells (PBMCs), modifications in this method are required for a more rapid and economic output for biobanks and clinical laboratories, particularly in developing countries. In this study, we addressed this issue by modifying the Ficoll-Paque method for the isolation of PBMCs or mononuclear cells from the peripheral and the umbilical cord blood of healthy and diseased (infected, anemic, and chronic obstructive pulmonary disease) adult individuals. In the modified method, we initiated the cell isolation process from the buffy coat layer, which appears in the interface between the plasma and sediments after centrifugation, instead of using the whole blood as described in the classic method. Although the PBMC yield by the modified method was about 12% less than in the classic method, the number of PBMCs isolated by the modified method was more than one million, which is enough for different research/diagnostic purposes, such as multi-omics detection. Assessment of cell viability and purity by hematology analyzer and trypan blue showed no significant difference between the viability and purity of the PBMCs isolated by these two methods in almost all groups, except samples from the infected and cord blood groups, where lower PBMC purity with higher granulocyte contamination were observed. In addition, at delayed processing time points, all parameters for the two methods were decreased in a time-dependent manner, especially at 8, 12, or 24 hours after the sample collection. In summary, the performance of PBMC isolation by the classic and modified methods mainly relies on the PBMC ratio in original samples. The modified method could be preferred for PBMC isolation because of its time and cost savings, especially for the biobanks and clinical laboratories in developing countries.
Introduction
B
According to the manufacturer's instructions and several published protocols, classic PBMC isolation from 2 mL whole blood uses 4 mL Ficoll reagent, several 15 mL conical tubes, and two to three types of swinging bucket rotors.13,21 Using large volumes of Ficoll medium becomes expensive for large-scale sample processing and rotor changes are labor intensive for routine work. With the improvement in specificity and sensitivity of detection technology, it is necessary for us to explore a new solution for PBMC isolation. Several studies have applied the buffy coat as a substitute for the whole blood for PBMC isolation,9,22,23 but to date, no study has systematically established and evaluated the difference of PBMC quality isolated from buffy coat or whole blood, especially in patients suffering from a variety of diseases. In this study, we collected peripheral whole blood from the healthy donors and patients with common physiological and pathological conditions. Each individual blood specimen was aliquoted into two equal parts, then PBMCs were isolated simultaneously by using the classic and modified methods. Critical parameters were assessed, including cell yield, purity, and viability. Several factors potentially affecting lymphocyte isolation by the Ficoll procedure were also investigated, such as a delay to processing time and centrifugal force. Overall, our findings provide an economical and feasible modified Ficoll method that uses a reduced quantity of Ficoll, thereby promoting the sustainable development of biobanks and clinical laboratories in developing countries.
Materials and Methods
Ethics statement
Healthy donors and patients from January 2017 to June 2017 in Gansu Provincial Hospital, China, were selected in accordance with the study protocols. The age and sex were matched among groups. The average age of subjects was 35.31 ± 12.82 years. The study was approved by the Ethics Committee of Gansu Provincial Hospital (approval no. syll20130308), and all clinical investigation was conducted according to the principles expressed in the Declaration of Helsinki. All participants gave written informed consent before the blood draw.
Blood sample preparation
All samples were collected in 2 mL ethylene-diaminetetraacetic acid-K2 vacuum blood tubes from donors fasting for more than 8 hours, mixed immediately after the collection by inverting 10 times, and shipped to the lab at 22°C–26°C in less than 30 minutes. All unprocessed blood samples were counted by an XN-1000 Automated Hematology Analyzer (Sysmex, Japan), including complete blood counts and differential hematological counts.
The study was divided into two parts. In the first part, based on the data of cell differential counts and clinical diagnosis, the participants were classified into six groups: a normal group (NG, n = 6, white blood cells: 4–10 × 109 cells/L), an infected group (IG, n = 6, white blood cells: 11–25 × 109 cells/L), an anemia group (AG, n = 6, hemoglobin: 60–95 g/L), a chemotherapy group (CG, n = 6, white blood cells: 2–3.7 × 109 cells/L), a chronic obstructive pulmonary disease (COPD) group (n = 6, erythrocytes: 6–7.5 × 1012 cells/L), and an umbilical cord blood group (UG, n = 6, lymphocytes: 3–6.5 × 109 cells/L). Two blood samples, each of 2 mL volume, were collected from each participant. After counting, all samples were processed immediately for PBMC isolation. In the second part, to investigate the effects of delayed processing time on PBMC isolation performance, 12 whole blood samples, each of 2 mL volume, were collected from healthy donors (n = 6) and umbilical cord blood was collected from women in labor (n = 6). After cell counting, the blood tubes were stored for 4, 8, 12, or 24 hours at 4°C before PBMC isolation, and 0 hour was regarded as the control group.
PBMC isolation and cell counts
PBMC isolation was performed by using Ficoll density gradient centrifugation. The density of Ficoll medium was 1.077 g/mL (Solarbio, China). The protocols in which PBMCs were isolated from whole blood and buffy coat are hereafter called the “classic method” and the “modified method,” respectively. Each method used one of the duplicate blood tubes (2 mL) from each donor. In both methods, the centrifugation in Ficoll was carried out by using the same swinging bucket centrifuge (L530R; Cence, China) and the same acceleration and deceleration settings (3 and 0 respectively). For obtaining buffy coat, both settings were 3. The operators in the study were well trained according to a Standard Operating Protocol and randomly selected for sample processing. The general outline of PBMC isolation for both methods is presented in Figure 1.
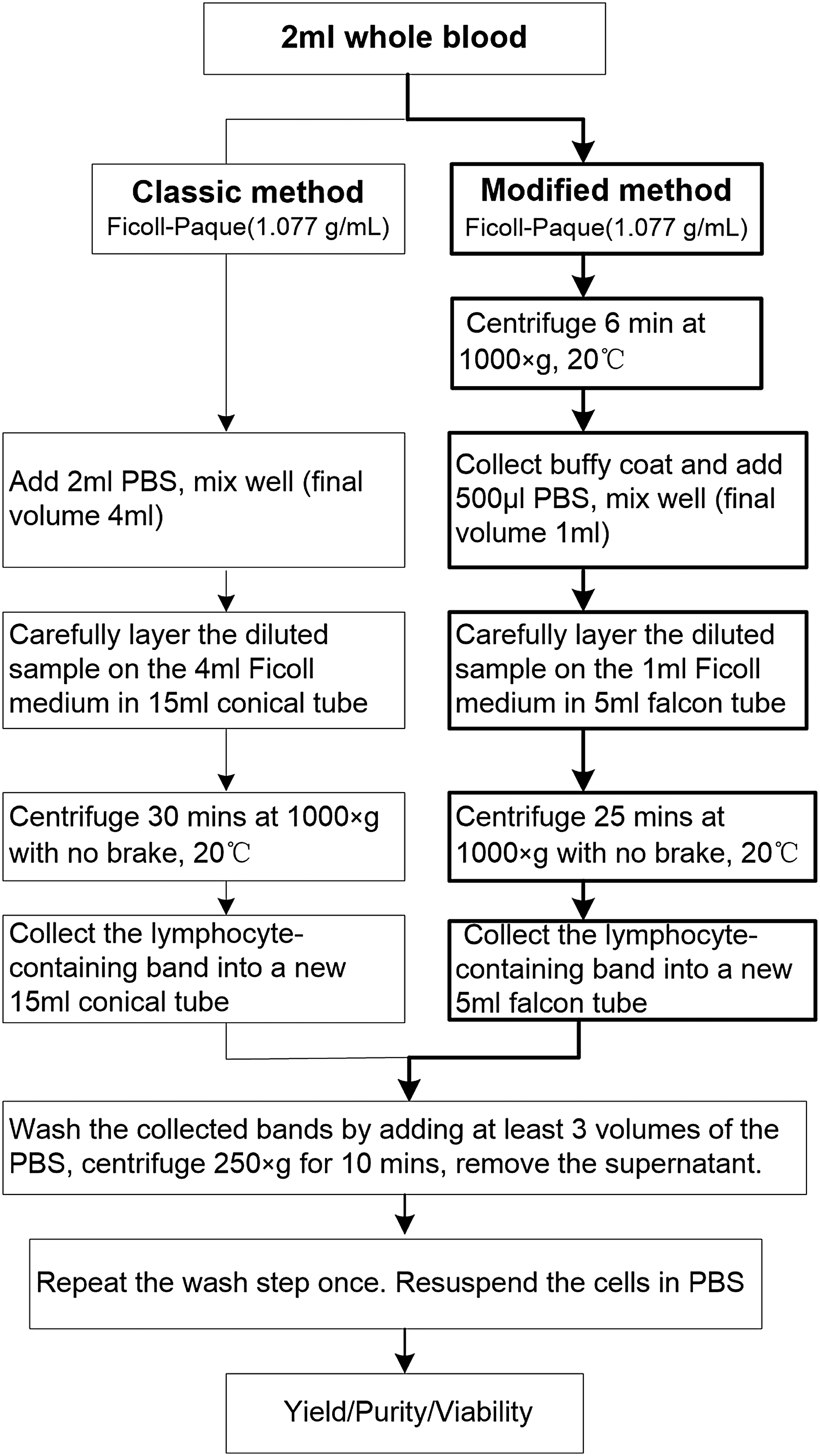
Schematic outline of classic and modified PBMC Ficoll-Paque density gradient centrifugation. PBMC, peripheral blood mononuclear cell.
In the classic method, 4 mL of Ficoll was placed in a centrifuge tube, then topped with the diluted fresh whole blood (2 mL in 2 mL phosphate-buffered saline [PBS], final volume 4 mL), and centrifuged at 1000 × g for 30 minutes and 20°C. This caused the cellular separation pattern typical of Ficoll density centrifugation to form, with the plasma uppermost, followed by a PBMC/platelet layer, a Ficoll layer, a granulocyte layer, and finally, at the base of the centrifuge tube, the erythrocytes. As much of the upper plasma layer as possible was drawn off by using a sterile pipette, the PBMC layer was transferred into a new 15 mL centrifuge tube, by using another clean pipette. The PBMCs were washed by adding three volumes of PBS, centrifuged at 250 × g for 10 minutes at 20°C, and the supernatant was removed. This washing step was repeated once and then finally, the PBMCs were resuspended in PBS. Thus, the classic method used a total of 4 mL Ficoll and two centrifuge tubes per blood sample.
In the modified method, the 2 mL fresh whole blood was centrifuged for 6 minutes at 1000 × g and 20°C. As no Ficoll had been added, this caused the typical three-layer separation seen when whole blood collected in anticoagulant tubes is centrifuged: an upper phase containing plasma, an interphase containing the buffy coat (PBMCs, granulocytes, and platelets), and a lower phase containing erythrocytes. The buffy coat interphase layer was collected as carefully as possible by using a sterile pipette, minimizing the aspiration of erythrocytes and plasma, then placed in a 1.5 mL centrifuge tube, and mixed with an equal volume of PBS (final volume 1 mL). Then, 1 mL Ficoll was placed in a 5 mL falcon tube (BD), the diluted buffy coat was carefully layered on top of it, and the tube was centrifuged for 25 minutes at 1000 × g and 20°C with no brake. This caused the buffy coat to separate into its constituent layers below the uppermost layer of residual plasma. As in the classic method, the plasma layer was discarded and the PBMC interphase layer was transferred into a new 5 mL falcon tube, by using another clean pipette. The PBMCs were washed by adding three volumes of PBS, and they were then centrifuged at 250 × g for 10 minutes at 20°C. Then, the supernatant was removed. This wash step was repeated once. Finally, the cells were resuspended in PBS. Thus, the modified method used 1 mL Ficoll and three centrifuge tubes per blood sample.
After their isolation, cells were analyzed on the Hematology Analyzer at low WBC mode and in manual operation. The absolute cell numbers of lymphocyte plus monocytes represented the whole PBMC count. The recovery rate was the ratio of the numbers of PBMCs before and after isolation. The percentage of lymphocytes plus monocytes was regarded as the purity of the recovered PBMCs. Cell viability was determined by the Trypan blue exclusion test (Sigma).
Statistical analysis
GraphPad Prism software v 6.0 (GraphPad Software) was used for statistical analysis. Analysis of statistical significance was performed by one-way ANOVA followed by Tukey's test among groups, or by an unpaired t-test between two groups. Results were evaluated with 95% confidence intervals, and the difference was considered statistically significant if p < 0.05.
Results
Yield and recovery rate of isolated PBMCs by classic and modified methods
To systematically evaluate the modified PBMC Ficoll isolation method, a series of blood samples were selected to cover various disease conditions. Based on the data from the counts by the Hematology Analyzer, the blood parameters in whole blood samples from the six groups are shown in Table 1 and Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/bio). Compared with the PBMC number in unprocessed whole blood, the PBMC isolation process led to some cell loss in both the classic and modified methods, with higher losses in the modified method (Fig. 2). However, absolute PBMC numbers were more than one million in all groups, even in the CG patients, regardless of the method of isolation.

The yield of PBMCs isolated by classic and modified methods in each subject. All cells before and after isolation were analyzed by the Hematology Analyzer. The PBMCs were defined as the sum of lymphocyte and monocyte.
Human peripheral whole blood and umbilical cord blood samples were classified by the Five Classification Automatic Hematology Analyzer. All data were presented as mean ± SD. In comparison with NG by the unpaired t-test, the significant differences were shown in the table. The level of significance was 0.05. *p < 0.05; **p < 0.01; ***p < 0.001.
AG, anemia group; CG, chemotherapy group; COPD, chronic obstructive pulmonary disease; IG, infected group; NG, normal group; SD, standard deviation; UG, umbilical cord blood group.
To correct for the discrepancies in initial cell populations in the unprocessed whole blood among the groups, a relative recovered PBMC yield was calculated for each patient by dividing the number of PBMCs recovered by using either the classic or the modified method by the number of PBMCs in the unprocessed blood from the same patient. The recovery rate from the classic method (60%–80%) was acceptable based on the manufacturer's instructions (Fig. 3A). In both the classic and modified methods, a statistically significant difference between the samples collected from the UG and CG group as well as between the UG and AG group was found (Fig. 3A, B). Across all the groups, the recovery rates using the classic and modified methods were 60%–80% versus 50%–70%. The recovery rate of isolated PBMC was 1.2–1.5 times greater in the classic method on average than that in the modified method (Fig. 3C), and the change in recovery rate was ∼12% among groups between the classic and modified methods (Fig. 3D). Reduced recovery rates using the modified method were found in the AG, CG, and COPD groups (Fig. 3C), whereas the rates of PBMCs isolated from CG and COPD patients had higher standard deviations (Fig. 3D).
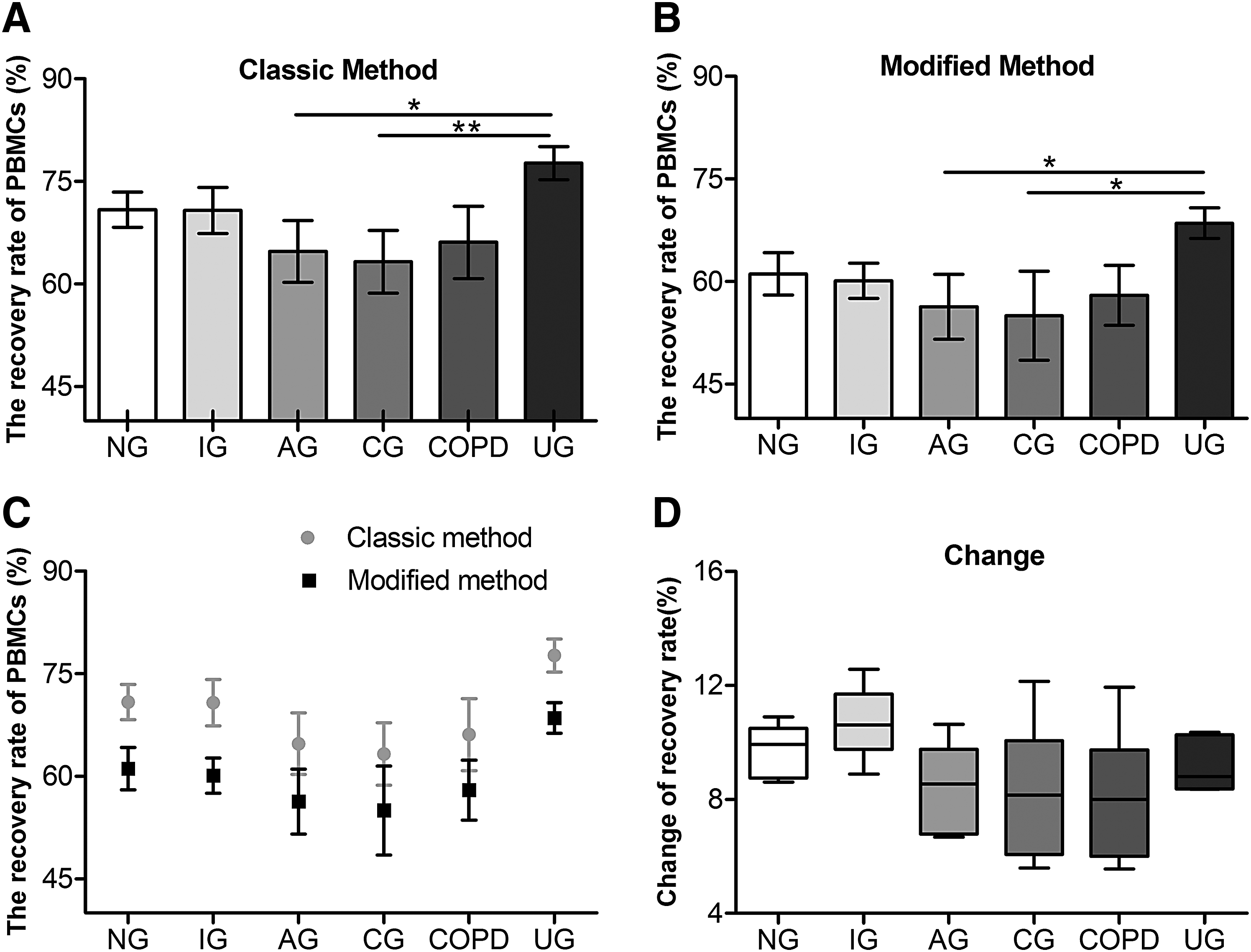
Comparison of recovery rate of PBMCs isolated by classic or modified method among groups.
Purity and viability of isolated PBMCs by classic and modified methods
The extent of contamination by other cells in isolated PBMC fractions varied depending on the pathological and physiological grouping of the patient, but were similar in the classic and modified methods for each pathological group (Fig. 4). So, both methods had similar purity (Fig. 4A). However, in different situations, the purity of recovered PBMCs showed a significant difference, especially in IG and UG, which had higher granulocyte contamination (purity of IG was 87% and UG was 84% in either method) and a greater coefficient of variances (Fig. 4B, C). The overall contamination by erythrocytes (red blood cells; RBCs) in isolated PBMCs was very low in all groups, and indeed undetectable by the Analyzer (Fig. 5); no difference was found between the classic method and the modified method, even in COPD patients.

Comparison of purity of PBMCs isolated by classic and modified method among groups.
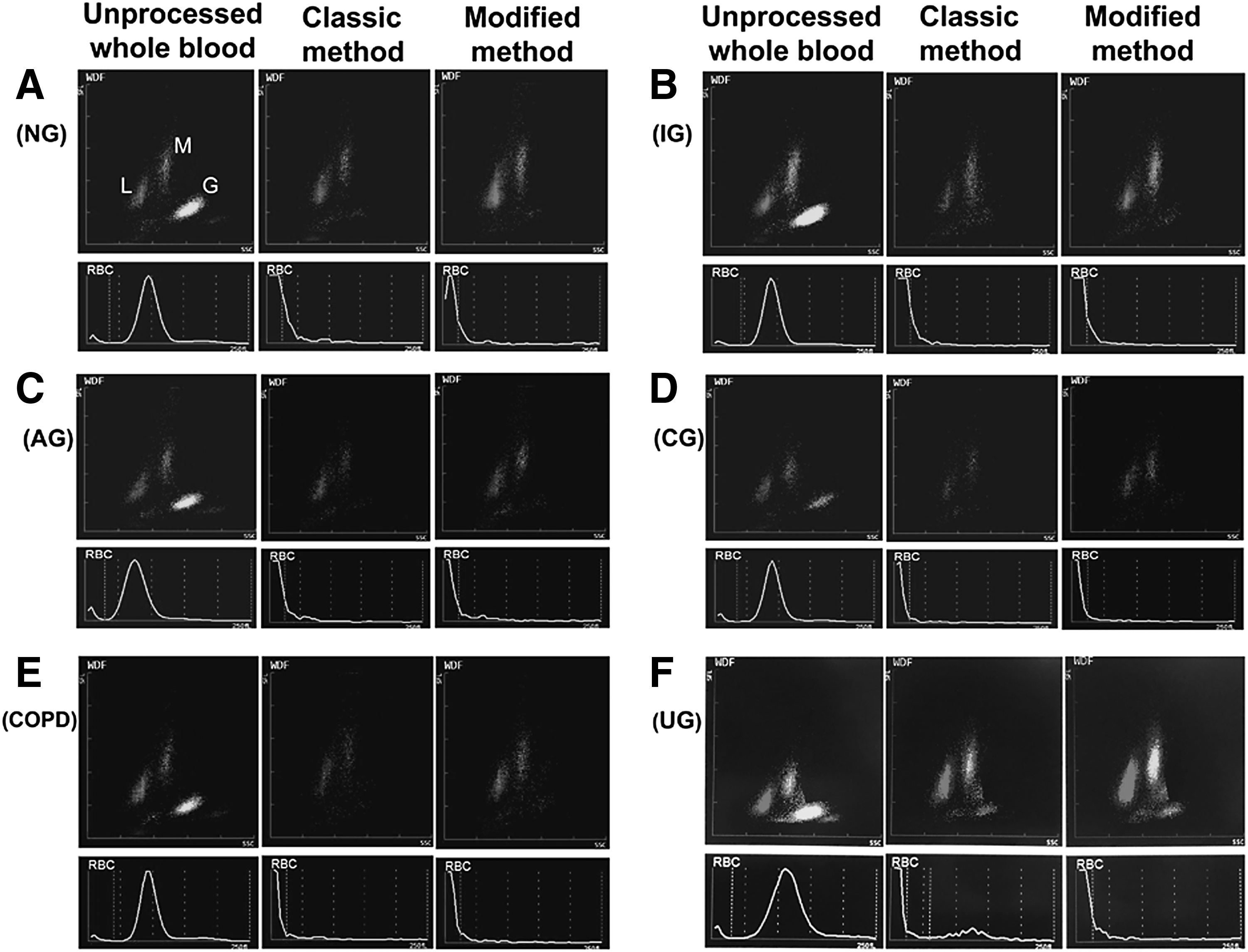
The contamination of erythocyte and granulocyte in isolated PBMC fractions by classic and modified methods among groups. All scatter diagrams were conducted on the hematology analyzer.
After isolation, cell viability was assessed by Trypan blue and was found to be >95% in both classic and modified methods. The data showed no significant difference between the two methods, or between the pathological and physiological groups (Supplementary Table S2).
Further, the effects of centrifugal force, 500 × g and 1000 × g, were assessed in modified method. No significant difference has been found in recovery rate and purity of PBMCs (Supplementary Fig. S1).
Effects of delayed processing time on yield and recovery rate of PBMCs isolated by classic and modified methods
Given the possibility of delayed processing time on isolation performance, whole blood from healthy individuals and umbilical cord blood was stored for 0, 4, 8, 12, and 24 hours at 4°C before isolation by both classic and modified methods. The recovery rates of isolated PBMCs were dramatically lower when delays to processing occurred (Fig. 6A, E). Regardless of peripheral whole blood or umbilical cord blood, the recovery rates dropped significantly after 8 hours of delay to processing time in both classic and modified methods (Fig. 6). Moreover, with a longer extension of delayed processing time, the yield became lower in both groups (Fig. 6C, D, G, H). For peripheral whole blood, at the 24 hours delay to processing time point, the recovery rate declined from 75% to 45% when the classical method was used to isolate PBMCs and from 65% to 35% when the modified method was used (Fig. 6A). Also, for umbilical cord blood, the recovery rate of the PBMCs by the classic method dropped from 80% to 45%, and from 70% to 35% by the modified method (Fig. 6E). However, there was no significant difference in the changes of recovery rate between the classic and modified methods at different delayed processing time in each group (Fig. 6B, F). Meanwhile, the data showed that there was a drop in cell count with the extension of delayed processing time, but in a less dramatic pattern (Fig. 7).
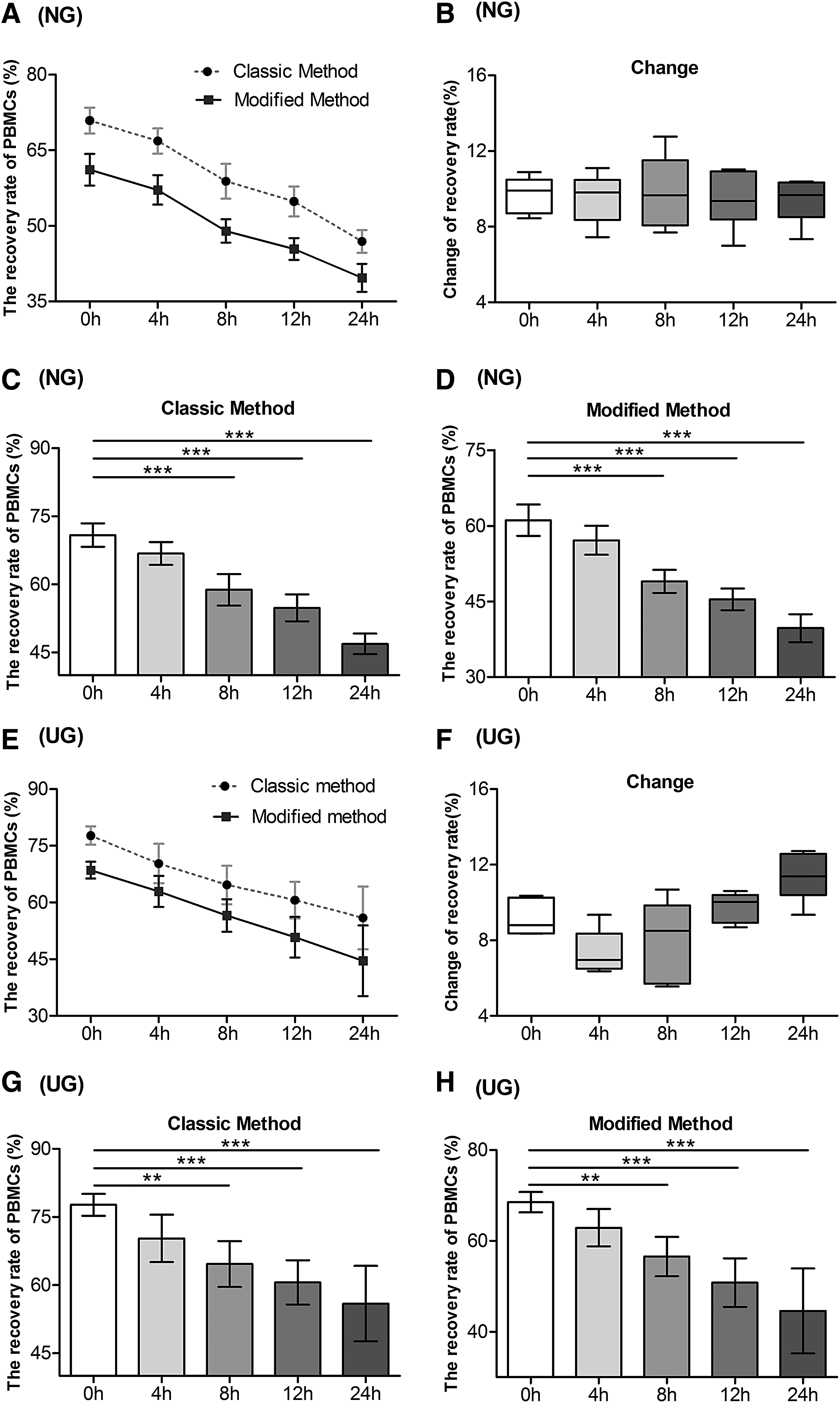
The recovery rate of PBMCs isolated by classic and modified methods at various delayed processing times in NG and UG.

The yield of recovered PBMCs isolated by classic and modified methods at delayed processing times in NG and UG.
Effects of delayed processing time on purity of PBMCs isolated by classic and modified methods
The purity of PBMCs isolated by the classic and modified methods when a delay to processing occurred was also evaluated. At each time point, the data showed no significant difference between the purity of PBMCs isolated by two methods in NG and UG (Fig. 8A, D). However, there was a significant decrease in PBMC purity after the 8 hours of delay in processing time in both groups (p < 0.01), in both the classic method (Fig. 8B, E) and the modified method (Fig. 8C, F). Cell viability was satisfactory at all delay to processing time points. Delays to processing time before the isolation did not alter cell viability (Supplementary Table S3).
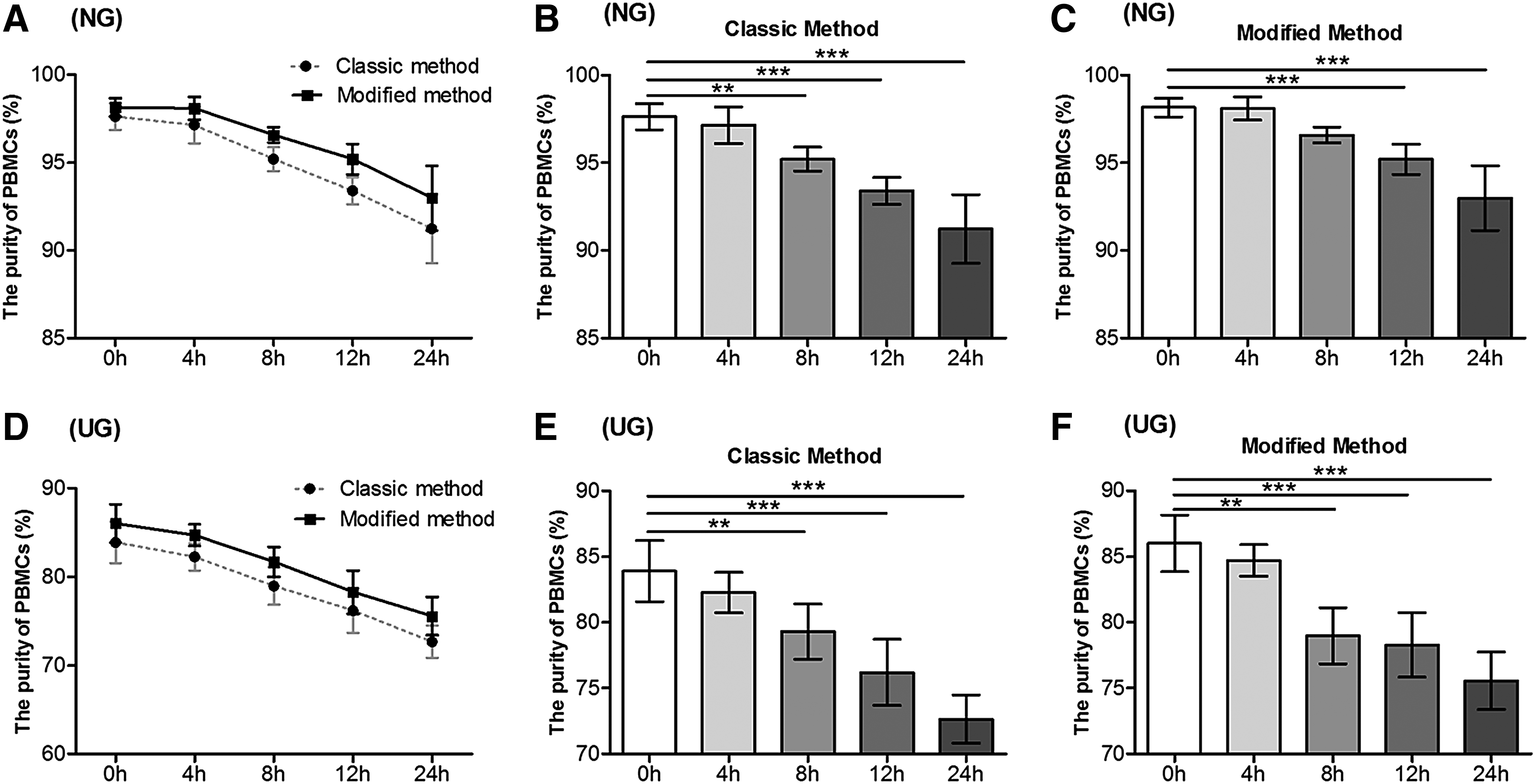
Comparison of purity of PBMCs isolated by classic and modified methods at various delayed processing time in NG and UG.
Discussion
To compare the modified with classic methods, critical parameters were systematically evaluated, including cell yield, recovery rate, purity, and viability. First, for cell yield and recovery rate, the results showed that there were higher yield and recovery rates of PBMCs isolated by the classic method than for the modified method. Although the PBMC yield in the modified method was about 12% lower than that in the classic method, more than one million PBMCs were recovered from 2 mL blood in all samples. Therefore, the number of cells recovered still meets the needs of downstream research.11,20,24 Higher recovery rates were found in IG and UG. It is well known that erythrocyte aggregation can reduce PBMC yield,12–14 but we did not get improved recovery rates in AG where the RBC cell counts were lowest, or poorer recovery rates in COPD, which had higher RBC numbers than the other groups. A big deviation of cell yield was also found in CG, AG, and COPD groups. This could be caused by the cell phase in different pathological conditions and suggests that the ratio of PBMCs contained in original samples could be a key factor for cell yield.
For cell purity and contamination, the proportion of major blood cell subsets in samples played a key role in isolation performance (Figs. 4 and 5). Our data showed that purity was highly dependent on the pathological and physiological condition. In the same groups, the two purification methods did not demonstrate any significant difference. However, in different groups, it was readily observed that lower purity and higher granulocyte contamination were found in IG and UG (Fig. 5), due to larger numbers of granulocytes contained in IG and UG than the other groups. There are methodological limitations for the Ficoll isolation method and, to some extent, granulocyte contamination has not been shown to have a significant influence on somatic mutation research and DNA methylation studies.25,26
Erythrocyte contamination was very limited in both the classic and modified methods, although some patients such as COPD and UG had higher RBC counts and larger hematocrit values than the others (Fig. 5). Lower RBC contamination in isolated PBMCs enables users to avoid the RBC lysis step in their nucleic acid extraction protocol, which likely improves the quality of the extracted nucleic acid.24,27
Our data show no significant differences in cell viability among the groups. These findings were consistent with the results provided from the manufacturer's instructions or published data, which suggested that more than 95% greater cell viability can be found.12,13,19,28,29
In addition, the study also assessed the impact of delayed processing time on critical parameters of isolated PBMCs by two methods, which is important for preserving and processing samples for biobanking and clinical assays. Our results showed that delayed processing time reduced the yield, recovery rate, and purity of the PBMCs isolated by the classic or modified methods, especially at delays of 8 hours or longer when the cells have been stored at 4°C before their purification. However, cell viability was not affected. These findings are in agreement with previously published studies.28,29 Also, the temperature of sample shipping, processing and storage, and centrifugal forces have been shown to affect the performance of the PBMC isolation method.12,13,19
Although the impact on PBMC yield as a consequence of the delay to processing would have been ameliorated if we had stored the whole blood at room temperature as opposed to 4°C (because apoptosis would be lower), the lower temperature is preferred by users who want to utilize plasma for proteomic and metabolomics studies. In biobanking, the aims of downstream studies are often unknown at the point of sample collection, and consequently it can be inadvisable to store blood at room temperature before processing. We therefore recommend, so that as many research needs as possible are fulfilled, that blood samples should be kept at 4°C but processed within 4 hours after collection.
Conclusions
This study demonstrated that classic Ficoll isolation of PBMCs could be replaced by our modified protocol. The modified method not only satisfies the demand of many research objectives, such as multi-omics analysis, but also offers a less expensive option because it uses less Ficoll. The original cellular subset ratio and the length of storage time had an impact on the PBMC isolation performance in both the classic and modified methods. Briefly, isolating PBMCs from buffy coat and then assessing them by a hematology analyzer, rather than insolating them from whole blood and analyzing them by a cytometer, could be an economical, practical, and efficient method in biobanks and clinical laboratories.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81360244) and Gansu Provincial Hospital (No. 16GSSY6-4).
Authors' Contributions
All authors have accepted responsibility for the entire content of this submitted manuscript and approved its submission.
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.