
Abstract
In vitro culture of ovarian follicles is a new technique in reproductive technology, which helps in understanding the process of folliculogenesis. The in vitro culture of follicles could be carried out using three-dimensional (3D) natural scaffolds that mimic the ovarian tissue stroma. Selection of the right matrix and culture media in these scaffolds could increase the survival and maturation of the follicles. In this work, the applicability of matrigel-alginate (MA) and fibrin-alginate (FA) 3D scaffolds for folliculogenesis was assessed. The ovaries of 13-day-old Naval Medical Research Institute (NMRI) mice were isolated and distributed into control and vitrification groups. Preantral follicles (mean diameter: 120–140 μm) were mechanically isolated from control and vitrified-warmed ovaries, encapsulated in MA or FA scaffold and cultured for 12 days. Follicle survival, growth, maturation, and quantitative expression of oocyte maturation genes (Gdf9, Bmp15, Fgf8, KitL, Kit, and Amh) and proteins (GDF9 and BMP15) were assessed. Survival rate of culture preantral follicles in control groups was found to be significantly higher than vitrified follicles. Antrum formation was similar in all groups. Follicle diameters were significantly increased in all groups during culture period. A decreasing pattern of gene expression was seen for all genes in all groups. This trend was verified through evaluation of protein expression, during which there was strong staining in antral follicles from all groups in the last day of in vitro culture. The better survival and maturation rate of follicles in the MA compared to FA scaffold indicates that the MA matrix, being rich in extracellular matrix components, could mimic the ovarian condition better and presents a good environment for follicle development.
Introduction
A
To encapsulate follicles for 3D in vitro culture, several matrices have been used. Alginate, extracted from brown algae, is one of the most frequently applied hydrogels.3,8–10 Despite the promising results obtained with this matrix, 11 alginate is an inert polymer 12 and must be supplemented to support follicle adhesion and improve cell survival and development.
A combination of alginate and extracellular matrix as a scaffold could result in better follicle growth and survival. For instance, matrigel, extracted from the Engelbreth-Holm-Swarm mouse sarcoma, is composed of a combination extracellular matrix (ECM) materials such as laminin, type IV collagen, heparan sulfate proteoglycans, and nidogen, which are some of the proteins and proteoglycans present in the ovarian tissue and follicle basement membrane.13–15
Another option is to combine alginate and fibrin. Fibrinogen is a soluble 340 kDa protein that is involved in blood clot formation, when it is coagulated into fibrin through the enzymatic activity of thrombin, in the presence of calcium. It is a biocompatible and nontoxic naturally derived scaffold, the features of which can be controlled by changing the components according to the requirements of the application. 16
The combination of fibrinogen with alginate for follicle culture has been reported in the literature. 17 Fibrin, by itself, cannot support in vitro follicle development because encapsulated follicles secrete proteolytic enzymes that degrade the matrix, which leads to extrusion of follicles. However, the Fibrin–Alginate interpenetrating network consists of degradable and nondegradable components and can be successfully applied for in vitro follicle culture; it has been reported to be a suitable scaffold for this purpose. 18
Follicle development is a complicated process that is driven by different factors including hormones and growth factors produced by follicular cells and oocytes. Paracrine and autocrine signals, such as members of the transforming growth factor β (Tgfβ) family, Kit-Kit ligand, and Fgf mediate mutually between the cellular compartments and orchestrate follicle development. 19 Growth differentiation factor 9 (Gdf9) and bone morphogenic protein15 (Bmp15) are the two most popular members of the transforming growth factor β superfamily genes that are known to regulate granulosa cells proliferation and differentiation. 20 Anti mullerian hormone (Amh) is another member of this superfamily, the expression of which influences follicle development and function. 21
Kit ligand (KL) is produced by granulosa cells while its receptor, Kit, is expressed on the surface of the oocytes. Kit-KitL is found to be expressed during all follicular stages. 22 Fgf8 is involved in different processes such as cell growth, embryo development, and morphogenesis. Fgf8 is expressed by oocytes in preantral to antral follicles and is necessary for the functioning of the granulosa cells.
The objective of this study was to evaluate survival, antrum formation and maturation rate, and relative expression of oocyte genes and proteins in three dimensionally cultured preantral follicles derived from vitrified-warmed and fresh mouse ovarian tissue.
Materials and Methods
Animals
Male and female adult Naval Medical Research Institute (NMRI) mice were purchased from the Pasteur Institute of Iran. They were housed and bred in rooms with controlled temperature (20°C–25°C) and lighting (12 hours light: 12 hours dark) and provided with food and water ad libitum. They were handled according to the ethical guidelines set by Royan Institute. A total of 200 ovaries were removed from13-day-old female NMRI mice and randomly allocated in two groups: fresh control and vitrification.
Study design
To conduct the study, two main groups were designed: Fresh control and vitrification.
Isolated preantral follicles in each group were cultured in two different 3D matrices comprising matrigel-alginate (MA) and fibrin-alginate (FA) scaffolds. The groups were accordingly named:
Control/matrigel-alginate scaffold: Cont/MA
Control/fibrin-alginate scaffold: Cont/FA
Vitrification/matrigel-alginate scaffold: Vit/MA
Vitrification/fibrin-alginate scaffold: Vit/FA
Vitrification and warming
Vitrification and warming were performed as previously described, with slight modifications to the warming process. 23 Ovaries were first equilibrated in 1.5 mL α-minimal essential medium (α-MEM; Gibco, Paisley, United Kingdom) supplemented with 7.5% ethylene glycol (EG; Sigma, MO), 7.5% dimethyl sulfoxide (DMSO; Sigma), and 20% fetal bovine serum (FBS; Gibco) for 15 minutes at 4°C. They were then transferred to 1.5 mL vitrification solution (α-MEM supplemented with 15% EG, 15% DMSO, 0.5 M sucrose (Sigma), and 20% FBS) for 30 minutes at 4°C. After this, they were loaded in an acupuncture needle (Dong Bang, Boryeong, Korea), immediately plunged into liquid nitrogen (LN2) and stored in LN2 for a week. For warming, vitrified ovaries were washed in a descending concentration of sucrose (1, 0.5 and 0.25 M in α-MEM and 20% FBS) at room temperature and were transferred to α-MEM and 10% FBS. They were incubated at 37°C for 30 to 60 minutes.
Follicle isolation
Preantral follicles (diameter: 120–140 μm) were mechanically isolated in both fresh and vitrified-warmed ovaries under a stereomicroscope.
MA scaffold preparation
Lyophilized Matrigel (QGel SA, Lausanne, Switzerland) was dissolved in its buffer (buffer was accompanied with lyophilized matrigel pack). Then, the liquid form of matrigel was maintained at 4°C for further use. Simultaneously, 15 g sodium alginate (Sigma) was dissolved in 10 mL deionized water. This was purified of organic impurities by adding activated charcoal (0.5 g charcoal/g alginate). The solution was then filtered under sterile conditions through a 0.22 mm filter and diluted with PBS solution (phosphate buffered saline) to reach a concentration of 1% (w/v). For preparation of the matrigel alginate scaffold, the matrigel and alginate solution was used in a 1:1 ratio. Five microliters droplets of matrigel alginate solution were placed on glass slides covered with parafilm and individual preantral follicles of control and vitrification groups were inserted into each droplet. Then, the droplet was immersed in a calcium bath [140 mM NaCl (Sigma) and 50 mM CaCl2 2H2O (Sigma)] for 2 minutes to achieve gelatinous state of scaffold.
FA scaffold preparation
Fibrinogen (50 mg/mL; Sigma) was dissolved in aprotinin solution (3000 kIU/mL; Roche, Mannheim, Germany). The 1% (w/v) alginate solution was prepared by dissolving 1 g sodium alginate (Sigma) in 10 mL deionized water and purified with activated charcoal as described earlier. The solution was filtered and diluted to 0.5% (w/v) with PBS solution and then mixed with fibrin solution in a 1:1 ratio. Thrombin (Sigma) was dissolved in 40 mM CaCl2 solution. 7.5 μL Fibrin–alginate droplets were pipetted onto a glass slide covered with parafilm and a single follicle was inserted into a droplet. Then, 7.5 μL thrombin solution was added to every droplet. The glass slide was placed upside down on a petri dish and transferred to the 37°C incubator for 5 minutes to achieve the gelatinous state of the scaffold.
Follicle culture
MA and FA scaffolds containing individual follicles were transferred to a 96-well plate and cultured for 12 days in 100 μL α-MEM supplemented with 10 mIU/mL follicular stimulating hormone (FSH; Merck; Germany), 1% ITS (5 mg/mL insulin, 5 mg/mL transferrin, 5 ng/mL sodium selenite; Gibco), and 10% human follicular fluid and 50 ng/mL human activin-A (Sigma) at 37°C, 100% humidity, and 5% CO2. Half of the culture media was exchanged every other day. Follicle diameter, antrum formation, and follicle survival rates were measured using an inverted microscope (Nikon, Düsseldorf, Germany) at 0 and 12 days of culture. On the 12th day of culture, the medium was exchanged with the maturation medium (α-MEM supplemented with 10 mIU/mL FSH, 1% ITS, 10% human follicular fluid and 50 ng/mL activin A, 1.5 IU/mL human chorionic gonadotropin (HCG, Merck; Germany), and 5 ng/mL epidermal growth factor (Sigma). Oocyte maturation (MII) rate was evaluated 14–16 hours later by checking for the presence of the first polar body under an inverted microscope.
Gene expression evaluation
For gene expression evaluation, follicles from control and vitrification groups were collected on days 0 and 12 of culture. They were pooled in Cell Reagent RNA Protect (Qiagen, Hilden, Germany) in three replicates, and stored at −70°C until RNA extraction. Purification of total RNA from follicle pools was performed using a RNeasy Micro Kit (Qiagen) according to the manufacturer's instructions. cDNA synthesis was done by using RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas, Leon-Rot, Germany) and random hexamers according to the manufacturer's instructions.
Specific primers for Gdf9, Bmp15, Fgf8, Kit, KitL, and Amh genes were used according to our previous studies.10,23 β-tubulin was used as an endogenous control.
Immunohistochemistry
The expression of GDF9 and BMP15 proteins was detected qualitatively by immunohistochemistry. Fresh control and vitrified-warmed 13-day-old mouse ovarian tissue (for preantral follicle assessment) and in vitro cultured follicles on 12th day of culture in all groups were fixed overnight in 4% paraformaldehyde at 4°C. Samples were trimmed, dehydrated with ethanol, cleared in xylene, and infiltrated with paraffin wax. Each paraffin-embedded ovary or follicle was sectioned and placed on the plus slide using a microtome. Polyclonal antibodies (Abcam, Cambridge, United Kingdom) were used for GDF-9 and BMP-15 protein expression. Ovarian tissue was used as a positive control. Antigen retrieval was performed by incubation in citric acid solution at 95°C for 20 minutes. Blocking against nonspecific binding was done using a solution consisting 10% normal goat serum in PBS for 1 hour at 37°C. Samples were incubated with primary antibody (1 μg/mL) for 24 hours at 4°C. After rinsing, the slides were incubated with secondary antibodies for 1 hour at 37°C.
Statistical analysis
In this study, descriptive statistics for continuous variables were presented as mean ± SD. A one-way analysis of variance, followed by the Duncan post hoc test was used to compare groups. Data analysis was conducted using SPSS version 16.0 (SPSS, Inc., Chicago, IL). All statistical tests were two-sided and p < 0.05 was considered statistically significant.
Results
Follicular growth and morphology
Preantral follicles with two layers of granulosa cells were isolated from 200 ovaries (Control and Vitrification) and randomly distributed in MA and FA groups for in vitro culture. The spherical structure of follicles was preserved during the 12 days culture period. Antrum cavity appeared on day 8 and completed with acentric oocyte position on day 12 of culture in all four experimental groups (Fig. 1). In the FA scaffold, the fibrin component gradually started to degrade due to secreted proteolytic enzymes by growing follicles, from day 4 until day 12. So a transparent circle was created around the follicle margin and developed toward the edges of the surrounding scaffold with follicle growth.

In vitro follicle development on days 0 and 12 of culture. Scale bar = 50 μm. Cont/FA, control/fibrin-alginate; Cont/MA, control/matrigel-alginate; Vit/FA, vitrification/fibrin-alginate; Vit/MA, vitrification/matrigel-alginate.
Follicular survival, antrum formation, and oocyte maturation
The survival rate of follicles was assessed at the end of culture period (day 12) in all experimental groups and it was significantly decreased in vitrification groups compared with the control groups and also in FA groups compared with the MA groups. Antrum formation was assessed on day 12 of culture and there were no significant differences between all groups. Oocyte maturation rates were similar in control and vitrification groups but higher in the MA group compared with the FA group (Table 1). Follicular diameter increased during the culture period and the mean values on day 12 of culture were 399.1 ± 4.71, 418.2 ± 11.5, 367.9 ± 16.36, and 363.9 ± 17.6 μm (mean ± SD) in Cont/MA, Vit/MA, Cont/FA, and Vit/FA respectively (Fig. 2).

Growth rate of follicles during in vitro culture. A versus a shows significant difference. p < 0.05.
Groups with same superscripts did not show significant difference at the 0.05 level.
Cont/FA, control/fibrin-alginate; Cont/MA, control/matrigel-alginate; Vit/FA, vitrification/fibrin-alginate; Vit/MA, vitrification/matrigel-alginate.
Quantitative gene expression
The expression of all assessed genes (Gdf9, Bmp15, Fgf8, KitL, Kit, and Amh) showed a reducing pattern and decreased at the end of the culture period in Cont/MA, Cont/FA, Vit/MA, and Vit/FA groups. Expression levels of Gdf9 and Fgf8 in vitrification groups were higher than control ones on day 0, however, significant differences were not seen between control and vitrification groups in other genes on both day 0 and 12 of culture. There were no differences between FA and MA matrices except the reduction in expression of Gdf9 in FA groups compared with MA on day 12 of culture (Fig. 3).
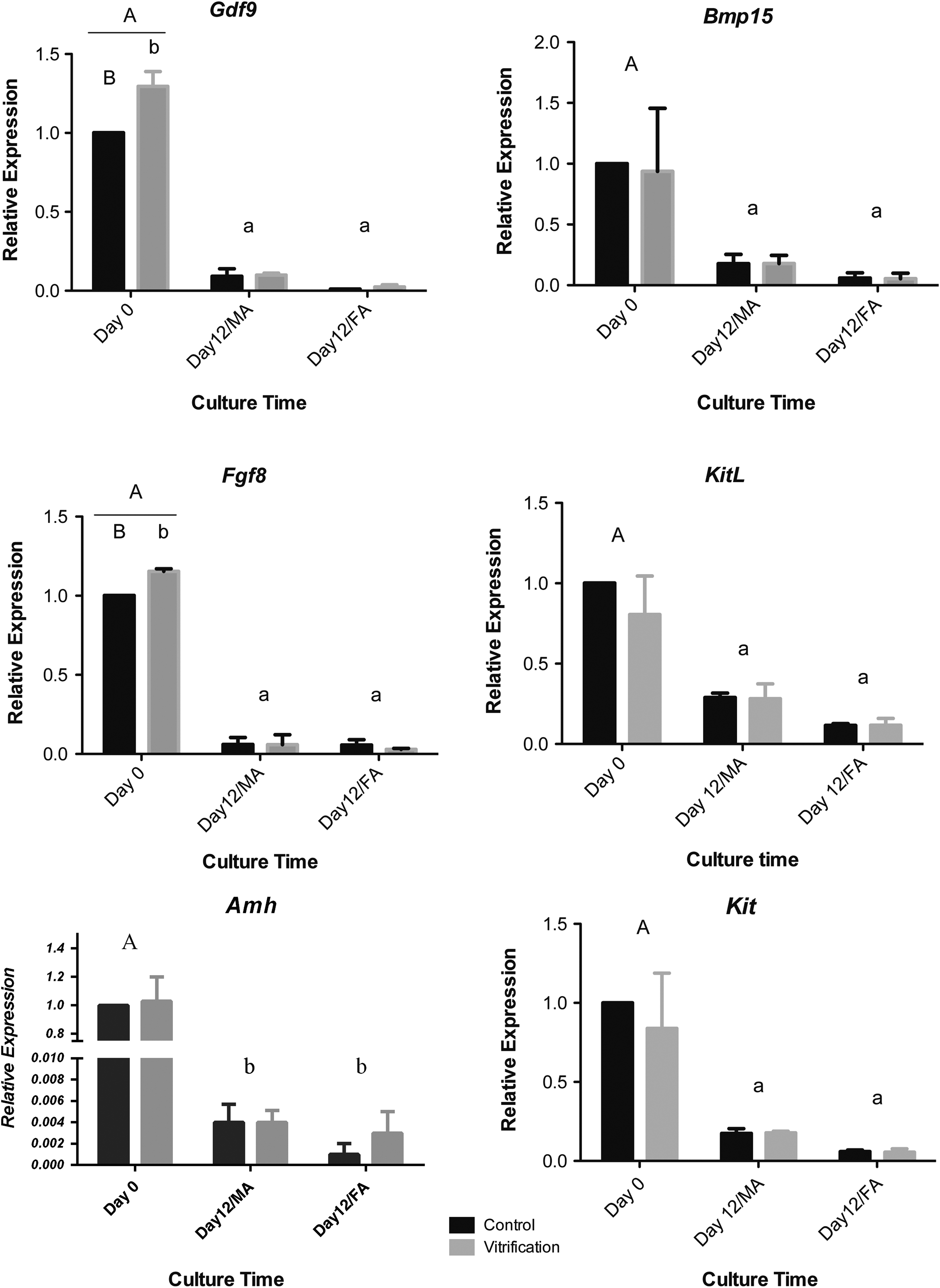
Relative genes expression of in vitro cultured follicles on day 0 and 12 of culture in all experimental groups. Same capital versus small letters (A vs. a, B vs. b) show significant differences. p < 0.05.
Protein expression
The expressions of BMP15 and GDF9 proteins were poorly detected in the cytoplasm of oocytes of preantral follicles in immature mouse ovarian tissue and clearly detected on the 12th day in vitro cultured follicles (antral stage) (Fig. 4).

Immunohistochemichal staining of mouse ovarian tissue (OT) and 12th day cultured follicles in all experimental groups for
Discussion
Different scaffolds must be evaluated for applicability in producing mature oocytes through 3D follicle in vitro culturing following vitrification. In our research, two important matrices (FA and MA) have been used according to previous studies.15,17,24 After 12 days of culture, follicles survival rate, antrum formation, growth rate, oocyte maturation genes (Gdf9, Bmp15, Fgf8, Kit, KitL, and Amh), and proteins (GDF-9 and BMP-15) expression in each group were evaluated and compared between experimental groups.
Although follicle survival rates were higher in control groups than vitrification groups, approximately similar antrum formation and maturation rates were observed in both groups. Accordingly, it seems follicles that could survive during the culture period following vitrification showed a suitable capability for developing toward antral stage. 23
Higher survival, growth, and maturation rates were observed in both MA groups. This could be because of the rich content of ECM in matrigel, the proteins playing a critical role in communicating with the intracellular cytoskeleton. The growth factors present in material also signal transmission. Besides, the interaction of activin A in culture medium with matrigel improves the quality of oocytes. 25
There was better mimicking of the in vivo condition for the MA groups and thus it was a better support for the follicle culture. MA therefore appears to be a better environment for follicle development. In the next step, we compared our experimental groups at the molecular level to achieve a better understanding of the in vitro folliculogenesis process. Growth differentiation factor 9 (Gdf9), bone morphogenetic protein 15 (Bmp15), and fibroblast growth factor 8 (Fgf8) are essential factors secreted by the oocyte for folliculogenesis and female fertility in a species-specific manner. In this study, the expression pattern of these growth factors was found to decrease during the culture period in all experimental groups, which is consistent with previous in vitro and in vivo studies.10,22,24,26 This shows the proper developmental procedure in all experimental groups. Higher expression of Gdf9 and Fgf8 genes on day 0 in the vitrification group compared with the control group may be due to the temperature reduction during the vitrification procedure and despite these initial changes, their expression was similar to the control groups at the end of the culture period in both Vit/MA and Vit/FA scaffolds. It appears that the expression increase at the beginning of the culture recovered during the culture period and further in vitro follicle development was not affected. Such changes in genes expression have been observed in earlier studies as well. 24 Amh is expressed in ovaries from the granulosa cells of secondary to early antral follicles; it inhibits initiation of follicle growth and sustains the arrested state of primordial follicles. It also plays a role in adjustment of the sensitivity to FSH in growing follicles.21,27 High expression of Amh was observed in isolated preantral follicles on day 0 and its level reduced in antral stage follicles (day 12) in all experimental groups. Previous studies showed the highest Amh expression in preantral and early antral follicles and very low Amh expression in the antral stage follicles in mouse ovary. 28 Expression of KitL has been shown in the granulosa cells of mouse, rat, ovine, and human follicles and it is required for oocyte survival and growth. KitL binds to its receptor c-Kit, a member of the tyrosine kinase receptor family and influences target cells. c-Kit and KitL expressions were high in preantral stage follicles and reduced in 12-day cultured antral follicles. It is indicated that expression of KitL increased in preantral follicles and remained high in the early antral stage but decreased in late antral follicles. 22 The expressions of GDF9 and BMP15 proteins were clearly seen in the oocyte of in vitro developed antral follicles through immunohistochemical staining. This provides good evidence for the observed significant decrease of Gdf9 and bmp15 expressions on day 12 of culture in all experimental groups and could demonstrate the translation of transcripts to proteins.
In vitro follicle culture following ovarian tissue vitrification could broaden the scope of fertility preservation. In fact, the possibility of transforming malignant cells after transplantation of cryopreserved ovarian tissue and probable cancer recurrence enhances the utility of this technique. One of the most important criteria for achieving oocyte maturity during cultivation is the selection of the scaffold that supports the follicle thoroughly.
Conclusion
The better survival and maturation rates of follicles observed in the MA compared to FA scaffold shows that the MA matrix, being rich in ECM components, mimics the ovarian condition better and provides a more conducive environment for follicle development.
Footnotes
Acknowledgments
This research was financially supported by the Royan Institute. There is no conflict of interest with this research.
Author Disclosure Statement
No conflicting financial interests exist.