
Abstract
This study investigates the possible causes for low development of blastocysts in vitrified immature oocytes by evaluating the changes of mitochondrial membrane potential and reactive oxygen species (ROS) production and finds a recovery mechanism for these conditions in vitrified immature oocytes. To recover from the cryoinjury, we cultured vitrified immature oocytes in milrinone containing medium for 1, 3, and 5 hours and then extended the culture for oocyte maturation. There was no difference in in vitro maturation and fertilization rate between fresh and vitrified/warmed oocytes. However, the development rate of blastocysts in vitrified/warmed oocytes was significantly lower than that in fresh oocytes (p < 0.05). The development rate of blastocysts was recovered if these oocytes were cultured for 3 hours in milrinone. Vitrified/warmed oocytes incubated in milrinone for 0 and 1 hour showed a significantly higher level of ROS (p < 0.05) and a significantly lower mitochondrial membrane potential (p < 0.05) than fresh oocytes. However, there was no significant difference (p > 0.05) between vitrified oocytes incubated in milrinone for 3 hours and fresh oocytes in terms of ROS level and mitochondrial membrane potential. In conclusion, alteration of highly polarized mitochondria distribution in vitrified oocytes may have an effect on mitochondrial activity, including ROS production during fertilization and further development. Preincubation in milrinone before in vitro maturation of immature vitrified/warmed oocytes may help the redistribution of highly polarized mitochondrial inner membrane potential and in reducing ROS and enhance the further embryonic development after fertilization.
Introduction
M
GV-immature oocytes punctured out of antral follicles are capable of spontaneous maturation to the MII stage and may achieve normal fertilization but cannot fully develop to blastocysts. Poor embryonic development of these oocytes may be due to asynchronous nuclear and cytoplasmic maturation during in vitro culture. 5 According to a recent report, treatment with phosphodiesterase inhibitors in prematuration in vitro (pre-IVM) culture suppressed meiotic resumption of bovine oocytes in vitro and improved development of blastocysts.6,7
Cryopreservation is a promising storage tool for surplus eggs, embryos, and sperm in assisted reproductive technology. The cryopreservation method of vitrification excludes the formation of ice crystals, which cause major cryodamage. Vitrification of MII mature oocytes has been successfully used. 8 However, this technique causes possible damage to subcellular structures, such as the meiotic spindle, and is related to abnormal fertilization with chromosomal abnormalities, mitochondrial function, and cortical granules. Immature oocyte cryopreservation may present an alternative to use MII structures for freezing. Oocytes arrested at prophase I have an intact GV that contains transcriptionally active decondensed chromatin. Cryopreservation of these GV oocytes may be more practicable, as they do not contain intact spindles. Cumulus and granulosa cells are tightly connected to eggs via the zona pellucida. Moreover, the plasma membrane has reduced cholesterol and fatty acid content, and therefore, its permeability to cryoprotectants is significantly higher compared with that of MII oocytes. 9 Immature oocytes can be collected without ovarian stimulation in cancer-related patients and may correspondingly present an opportunity for fertility preservation. 10 However, this procedure is not widely used owing to low success rates, which may be attributable to difficulties in preventing damage occurring during the freezing process or poor in vitro maturation results. Immature oocytes are connected tightly with cumulus cells through transzonal projections. Diffusion of cryoprotectants may therefore alter cumulus or granulosa cell connections with oocytes. 11 The survival rates of vitrified GV immature oocytes and MII oocytes are not significantly different. However, the maturation rates of vitrified oocytes are lower compared with those of fresh oocytes. 12 Despite the considerable advances in vitrification technology, vitrified mouse immature oocytes show reduced developmental rates 13 with altered and fragmented mitochondrial structures. 14
Mitochondria are specific organelles that generate adenosine trisphosphate (ATP), which contributes to energy metabolism for egg activation and further embryonic development. Mitochondrial membrane potential (Δψm) reflects differences in mitochondrial respiratory activity and is the key indicator of bioenergy synthesis in cells. 15 Increased Δψm is associated with oocyte cytoplasmic quality.
Oocyte cryopreservation is well established in human-assisted reproductive technology (ART) with mature MII oocytes. However, immature oocyte vitrification is currently marginal in ART, not necessarily as an effect of poor outcome derived from the double intervention of cryopreservation/IVM (in whatever order), but because of the scarcity of material and applicable cases. Here, we investigated a new method to recover from cryodamage in the immature oocytes vitrification process.
The main objective of this study was to investigate whether prematuration culture of immature oocytes in the presence of the phosphodiesterase inhibitor, milrinone, can wait to recover from cryodamage to mitochondrial function, before meiotic resumption, and to improve maturation of oocytes and further embryonic development.
Materials and Methods
Animals
All animal handling experiments were approved by the Institutional Animal Care and Use Committee of the CHA University (Approval No. IACUC 140027). Three to four weeks old B6D2F1 female mice (Samtako, Seoul, Korea) were maintained in temperature- and humidity-controlled conditions under light (12 hours) and dark (12 hours) cycles. For immature oocyte-enclosed cumulus cell (cumulus oocyte complex [COC]) collection, female mice were intraperitoneally injected with 5 IU pregnant mare serum gonadotropin (PMSG; Sigma-Aldrich, St, Louis, MO). At 44–45 hours after PMSG, immature oocytes were collected from the ovary of antral follicle by puncturing with a 29 gauge needle in Quinn's advantage medium with HEPES (Quinn's-HEPES; Sage In vitro Fertilization, Inc., Trumbull, CT) containing 10% serum protein substitute (SPS; Sage) and 5 μM milrinone, phosphodiesterase 3 inhibitor (Sigma-Aldrich).
Vitrification and warming process
Vitrification and warming processes were performed according to Kim et al. 16 For basic medium, all vitrification and warming processes used Quinn's-HEPES with 20% fetal bovine serum (GIBCO, Grand Island, NY). As a cryoprotectant (CPA), a combination of ethylene glycol (EG) and dimethyl sulfoxide (DMSO; Sigma-Aldrich) was used. COC was preequilibrated in 1.34 M EG +1.05 M DMSO for 2.5 minutes, followed by 2.68 M EG +2.1 M DMSO and 0.5 M sucrose for 20 seconds. CPA-equilibrated 10 COCs were loaded on an electron microscope (EM) copper grid and plunged into slush nitrogen (SN2) using a Vit-Master (IMT, Ness Ziona, Israel) and then stored in liquid nitrogen (LN2). 17 The warming process was performed with 1.0, 0.5, 0.25, 0.125, and 0 M sucrose for 2.5 minutes in each step on a 37°C slide warmer. Vitrified/warmed immature oocytes were cultured for 1, 3, and 5 hours in TCM-199 medium containing 100 μM isobutylmethylxanthine (IBMX) or 5 μM milrinone for recovery of cryodamage of cytoplasm, in a process called prematuration culture. Fresh COCs were used as controls.
Prematuration culture of COC was examined in an inner mitochondrial membrane potential test or measurement of reactive oxygen species (ROS) or in vitro maturation. For in in vitro maturation, COCs were incubated in TCM-199 medium containing 10% fetal bovine serum (FBS), 0.075 IU/mL recombinant follicle-stimulating hormone (rFSH, Gonal F; Serono, Modugno Bari, Italy), 0.5 IU/mL human chorionic gonadotropin (hCG, Ovidrel; Serono), and 1.0 μg/mL estradiol (Sigma-Aldrich) for 16 hours. Cumulus cells from mature oocytes were removed with 0.1% hyaluronidase (Sigma-Aldrich). Successful oocyte maturation was evaluated by the first polar body (first PB) extrusions. Oocyte maturation status was divided into germinal vesicle intact oocytes (GV), metaphase I without first polar body (MI oocytes), and MII with first polar body (MII oocytes). Denuded oocytes were subjected to fertilization in vitro.
Sperm was collected from the cauda epididymis of 10–12 weeks old male mice capacitated in Quinn's Advantage Fertilization medium (Sage) for 30 minutes. Capacitated spermatozoa were added (1–2 × 106/mL) into a mature oocyte culture dish and incubated for 6 hours. The eggs with two pronuclei were evaluated for fertilized (2PN) or two-cell formation at 24 hours postfertilization. Fertilized eggs were cultured for 5 days to facilitate embryonic development to the blastocyst stage (BL, Fig. 1).

Successful oocyte maturation, fertilization, and blastocyst formation.
Measurement of mitochondrial inner membrane potential and ROS of oocytes
Precultured or fresh oocytes were incubated in 2 μM mitochondrial membrane potential indicator, MitoProbe™ JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide; Molecular Probes, Grand Island, NY), in TCM-199 medium for 30 minutes at 37°C and followed by immediate live imaging by confocal laser scanning microscope (LSM 510Meta; Carl Zeiss, Germany). To confirm that JC-1 response is sensitive to changes in membrane potential, oocytes were incubated with mitochondrial membrane potential disrupter, 50 μM CCCP (carbonyl cyanide 3-chlorophenylhydrazone), for 5 minutes before with JC-1 incubation. The approximate excitation peak of JC-1 is 488 nm, the monomeric emission peak of low membrane potential is green, and J-aggregate forms an orange-red emission of high membrane potential. The ROS indicator, Image-IT® LIVE Green Reactive Oxygen Species Detection Kit (5-(and-6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate (carboxy-H2DCFDA); Molecular Probes), was used for ROS measurement. Precultured oocytes were incubated in TCM-199 medium containing 25 μM carboxy-H2DCFDA for 30 minutes and followed by 1 μM Hoechst 33342 for 5 minutes. Gently washed oocytes were observed on the CLSM or epifluorescence microscope (Axio Imager 2; Carl Zeiss) with 488 nm excitation and 495/529 nm emission and for Hoechst 33343, 350/461 nm excitation and DAPI filter. As a positive control, oocytes were incubated in 100 μM inducer of ROS production (tert-butyl hydroperoxide [TBHP]) for 60 minutes and measured ROS of each oocytes image.
Relative intensities of JC-1 or ROS of each oocyte were quantified using AxioVision software 4.8.1 (Zeiss, Germany).
Statistical analyses
All statistical analyses were performed with ANOVA using Tukey's post hoc test or t-test using GraphPad Prism 7 (GraphPad Software, Inc., La Jolla, CA). p Values <0.05 were considered statistically significant.
Results
Effects of prematuration culture with phosphodiesterase inhibitors on oocyte maturation and embryonic development in vitrified/warmed oocytes
Two different phosphodiesterase inhibitors were tested to determine the optimal conditions for prematuration of immature oocytes. Freshly collected COCs were cultured in 100 μM IBMX or 5 μM milrinone-containing TCM-199 for 1, 3, 5 and 12 hours. All oocytes maintained GV arrest in prematuration culture. After 16 hours culture in phosphodiesterase inhibitor-free medium for in vitro maturation, we observed no differences in oocyte maturation to second meiotic metaphase oocytes (MII) among the groups. Cleavage rates after in vitro fertilization were increased at 3 hours prematuration compared with 0 and 1 hour prematuration (p < 0.05). However, 12 hours prematuration oocytes showed very low cleavage and blastocyst formation rates (p < 0.05). Moreover, each prematuration stage in the presence of milrinone was more advanced, indicating higher embryonic development, compared with IBMX, although it is not statistically significant. Based on these results, we used 5 μM milrinone-containing TCM-199 for further experiments (Fig. 2). To evaluate oocyte maturation and embryonic development in vitrified/thawed eggs after prematuration culture, eggs were cultured in 10 μM milrinone for 1, 3, and 5 hours after thawing. In fresh eggs, the maturation rate to MII was 94.8% (±5.0), the fertilization rate was 73.6% (±9.3), and blastocyst formation from the two-cell stage was 54.6% (±7.9). In vitrified/thawed eggs, maturation, fertilization, and blastocyst formation rates were significantly decreased compared with fresh eggs. Notably, eggs subjected to 3 hours prematuration in milrinone recovered from cryodamage due to vitrification in terms of oocyte maturation, fertilization, and embryonic development (Table 1; 82.9% ± 8.4%, 71.1% ± 5.5%, and 48.1% ± 2.4%, respectively, p < 0.05).
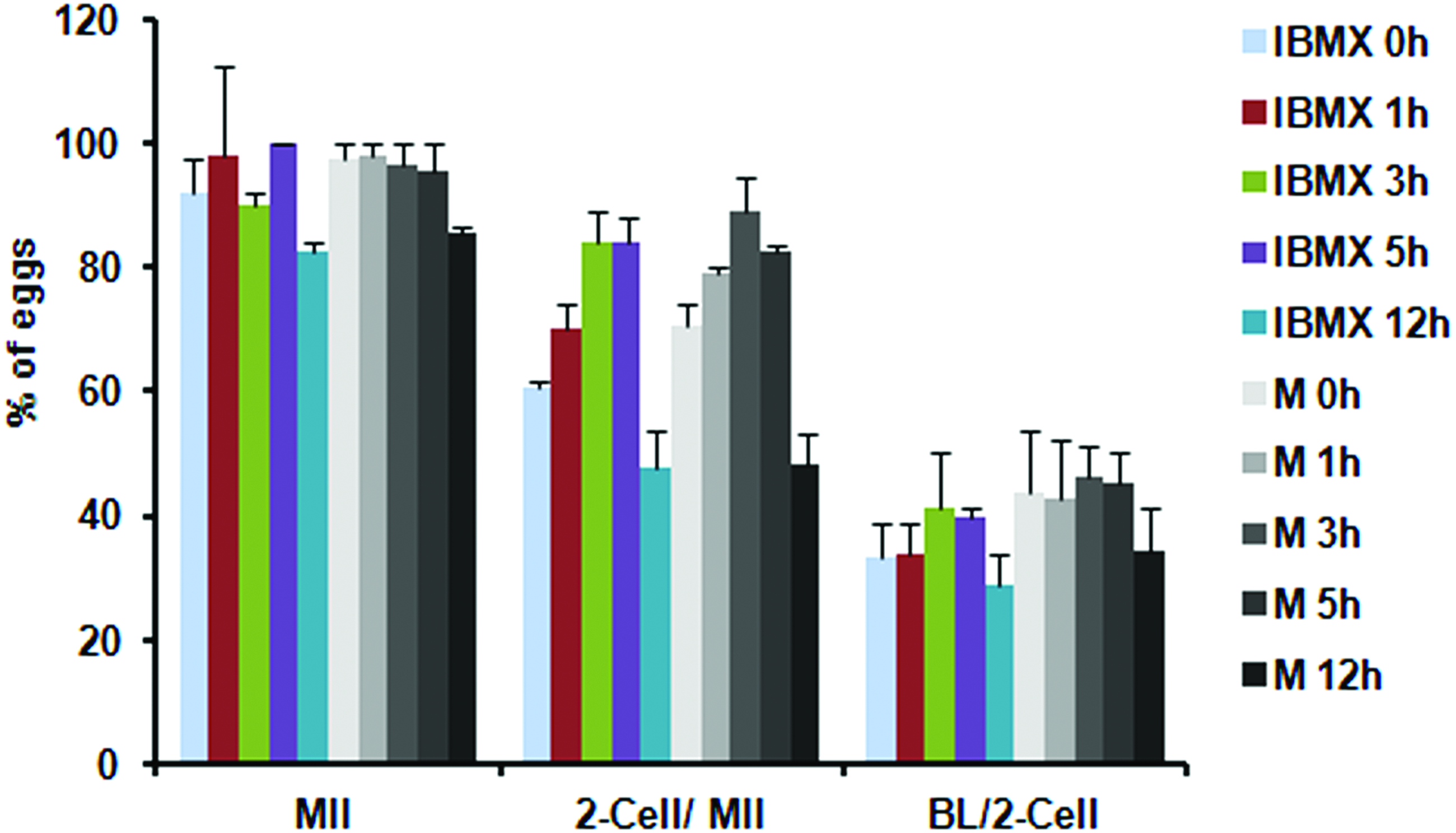
Oocyte maturation, fertilization, and embryonic development of precultured oocyte in 100 μM IBMX (I) or 5 μM milrinone (M) for 1, 3, 5, and 12 hours. COCs were subjected to prematuration culture in two different phosphodiesterase inhibitors, IBMX or milrinone (M), containing TCM-199 medium followed by phosphodiesterase inhibitors free medium for 16 hours to evaluate oocyte maturation (MII oocytes). Two-cell embryos (two-cell) were observed at 24 hours, and blastocysts (BL) were observed at 96 hours postfertilization. COCs, cumulus oocyte complexes; IBMX, isobutylmethylxanthine. Color images available online at www.liebertpub.com/bio
Data are represented as average (%) ± SEM for three replications.
Represents significant difference to fresh eggs (p < 0.05).
Represents significant difference to 0 hour prematuration culture (p < 0.005).
BL, blastocyst; GV, germinal vesicle oocytes; MI, metaphase I oocytes; MII, metaphase II oocytes.
Effects of prematuration culture with phosphodiesterase inhibitors on mitochondrial inner membrane potential and ROS production in vitrified/warmed eggs
To investigate the mechanisms underlying higher maturation, fertilization, and embryonic development of vitrified/warmed eggs subjected to prematuration culture with milrinone, we examined the mitochondrial inner membrane potential with JC-1 (Fig. 3). The majority of fresh eggs (7 of 12) showed a pattern of pericortical J-aggregate red fluorescence with high mitochondrial inner membrane potential (high Δψm), similar to a previous report. 15 The percentage of eggs with pericortical red fluorescence and high Δψm increased with prematuration culture periods in the fresh egg group. However, vitrified/warmed eggs from 1 hour prematuration culture displayed low Δψm with green fluorescence in the central cytoplasmic area, and the percentage of oocytes with high Δψm was significantly low (p < 0.05, Fig. 3 B). Moreover, these eggs did not show high Δψm with red fluorescence in the pericortical area. As the prematuration period was extended to 3 or 5 hours, high Δψm was recovered with red fluorescence. It showed a similar percentage of oocytes compared with fresh oocytes (Fig. 3B). The sensitivity of JC-1 response to changes in membrane potential was further examined. Pretreatment with 50 μM CCCP, a mitochondrial membrane potential disrupter, led to generation of the green signal only in the 5 hours prematuration group of fresh mature oocytes (Fig. 3A).
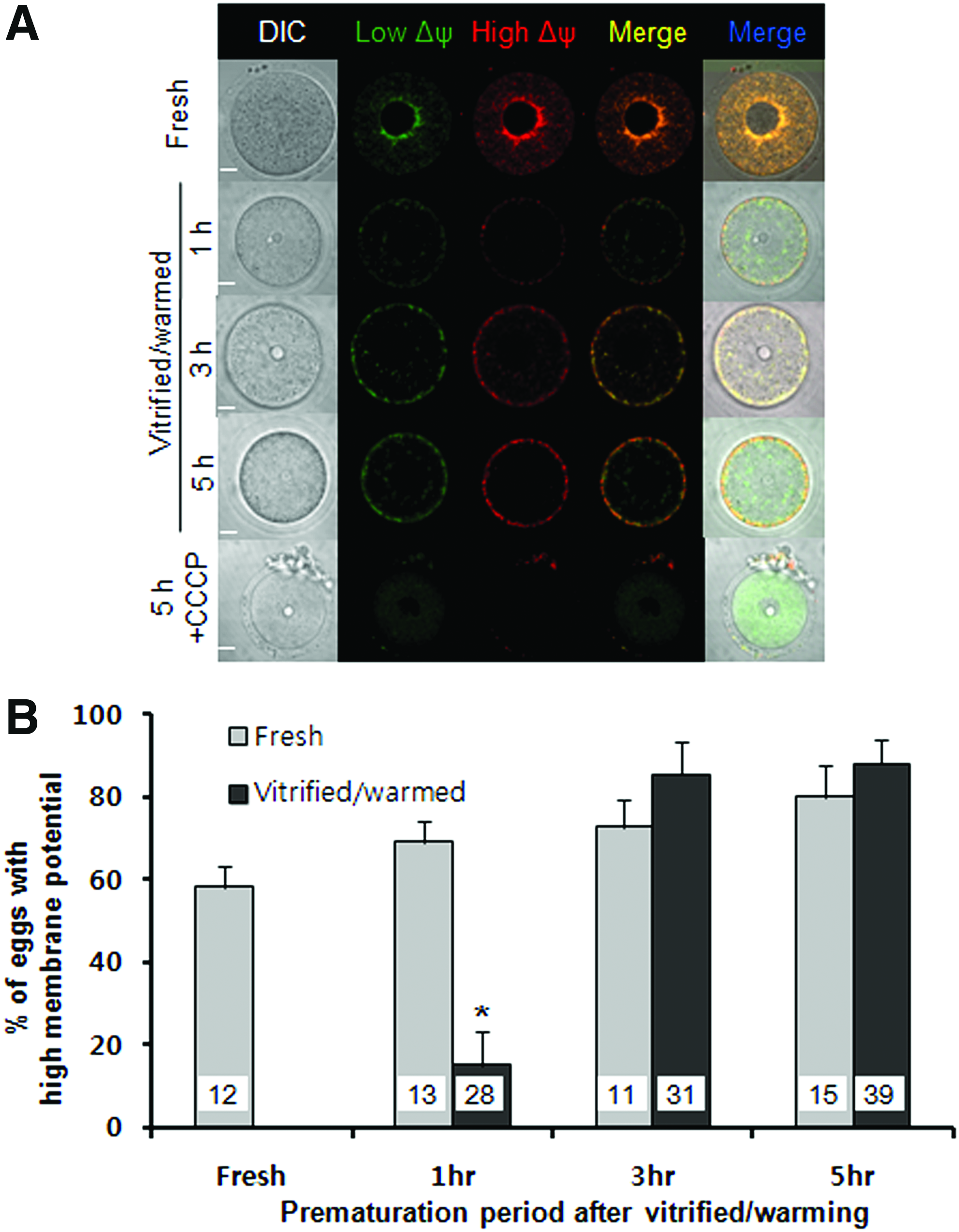
Representative confocal laser scanning microscope images of mouse fresh and vitrified GV oocytes stained with MitoProbe™ JC-1 (JC-1).
Vitrification induces damage to the endogenous antioxidant systems of eggs and increases the ROS in oocytes.18,19 Accordingly, we investigated the changes in ROS levels after prematuration of vitrified immature oocytes. Relative fluorescence intensities of ROS were compared between fresh and vitrified oocytes for each prematuration period. Immediately after thawing or 1 hour prematuration, the ROS level in vitrified oocytes was significantly higher than that in fresh oocytes (p < 0.05). Notably, the ROS level of vitrified oocytes from 3 hours culture after thawing was reduced to the same extent as that of fresh oocytes from 3 hours prematuration (Fig. 4).
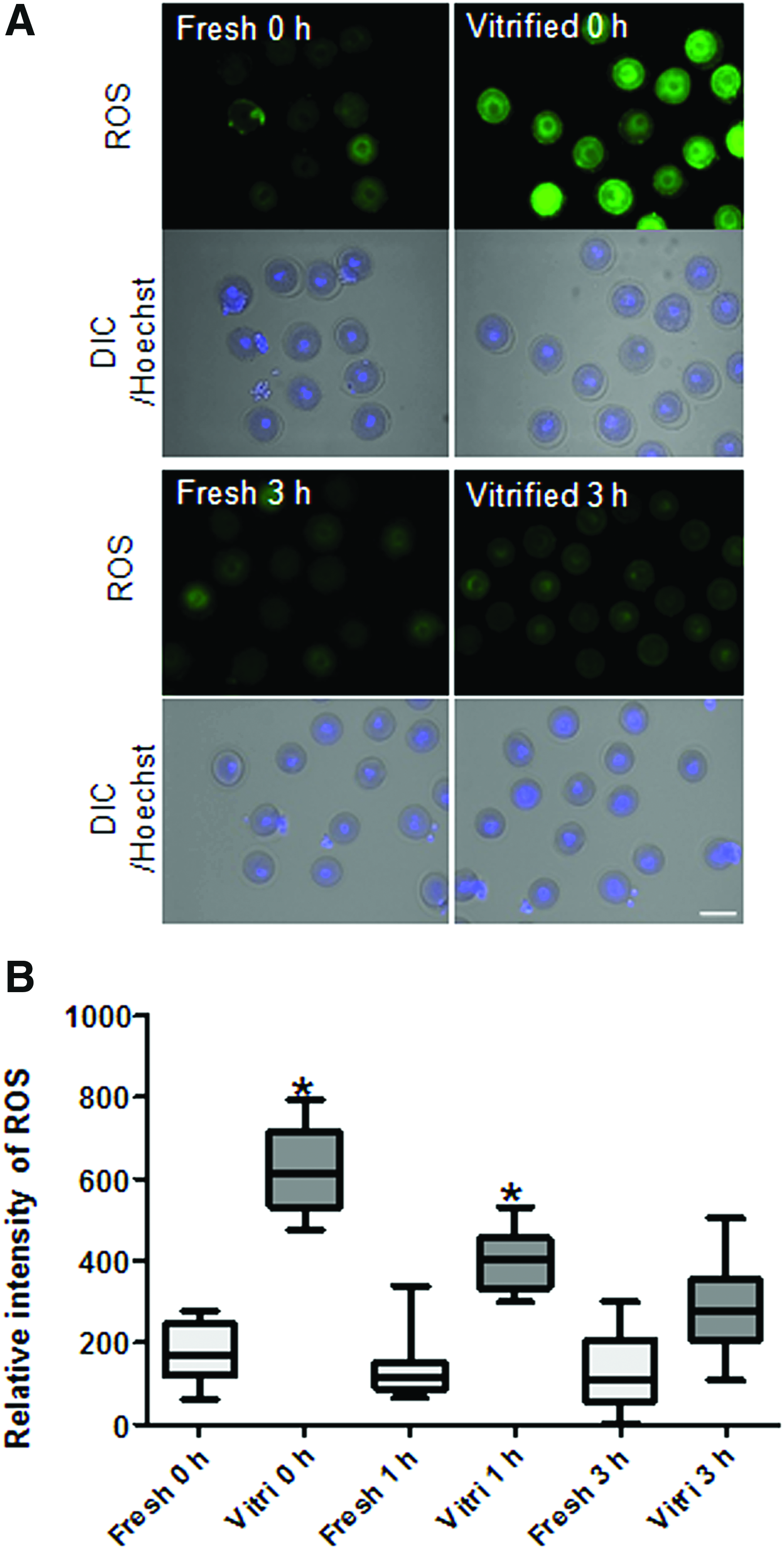
Representative images of mouse fresh and vitrified GV oocytes stained with ROS measurement.
Discussion
Cryopreservation is a critical option in reproductive medicine to preserve sperm, oocytes, embryos, and tissues, including ovarian and testicular tissues, for fertility treatment. Cancer patients undergoing chemotherapy, patients with premature ovarian insufficiency, and women who want to postpone childbearing are additional beneficiaries of oocyte cryopreservation. The first successful pregnancy using a frozen-thawed oocyte was reported by Chen in 1988, 20 although oocyte cryopreservation protocols were still under development. Several factors contribute to the difficulty of oocyte cryopreservation. Large water content may be the main cause of intracellular ice formation, osmotic damage, and chilling injury. The main injuries caused by mature oocyte cryopreservation include cytoplasmic alterations through disruption of the subcortical actin network 16 or spindle alterations through microtubule injuries. 21 Alternative immature oocyte cryopreservation protocols have been developed to resolve these difficulties.22–26 However, several concerns exist with regard to cryopreservation of immature oocytes. After vitrification and thawing, immature oocytes may initiate meiosis to MII before recovery from cytoplasmic cryodamage. Here, we focused on the effects of prematuration culture supplemented with milrinone as the phosphodiesterase inhibitor in vitro on immature mouse oocyte development after vitrification and thawing.
Two different phosphodiesterase inhibitors were examined with the aim of preventing spontaneous meiotic maturation through increased levels of intracellular cyclic adenosine monophosphate (cAMP). 4 According to recent reports, prematuration in the presence of IBMX modulates cumulus cell expansion and oocyte metabolism and enhances the developmental competence of in vitro matured mouse oocytes27,28 and bovine oocytes. 7 However, IBMX is a nonselective adenosine receptor antagonist, and its safety has not been evaluated and it has yet to be approved for clinical use. We additionally examined a novel phosphodiesterase 3 inhibitor, milrinone, used to treat patients with heart failure, which acts by suppressing degradation of cAMP and thus increasing phosphorylation levels of many components in the heart. 29 Prematuration of oocytes for 0, 1, 3, 5, and 12 hours with IBMX or milrinone was compared (Fig. 2).
The optimal duration period of prematuration is under investigation. Prematuration with either IBMX or milrinone for 12 hours led to low maturation, fertilization, and embryonic development to blastocysts, compared to the other groups. Female phosphodiesterase-3 A (PDE3A)-KO mice are completely sterile due to increased cAMP levels in oocytes, resulting in arrest at prophase I of the first meiotic division. 30 Hence, 12 hours prematuration culture with a PDE3 inhibitor may result in a nonreversible meiotic arrest. The optimal duration of prematuration with a PDE3 inhibitor was considered to be 3 hours since both fresh and vitrified oocytes in this group showed higher maturation, fertilization, and blastocyst formation rates than the other 1 or 3 hours prematuration groups.
Vitrified and thawed immature oocytes were subjected to prematuration with milrinone for 0, 1, 3, and 5 hours before in vitro maturation (Table 1). Damage of the cytoskeleton, including microtubules and microfilaments, during vitrification is the main cause of abnormal spindle arrangement and alterations in cytoplasmic organelles, such as endoplasmic reticulum and mitochondria. Vitrification-induced alterations in the inositol triphosphate receptor (IP3R) on the endoplasmic reticulum membrane lead to calcium oscillatory activity and affect mature egg quality and further embryonic development. 16 Several reports to date have shown that vitrification induces abnormal mitochondrial distribution 31 and decreased mitochondrial membrane potential.32,33 Moreover, high levels of ROS and low ATP production with reduced ATP synthase activities have been detected in vitrified/warmed oocytes 34 or embryos.18,19 These negative effects of vitrification on mitochondrial function contribute to decreased embryonic developmental competence. In the current study, vitrified/warmed oocytes showed very low mitochondrial inner membrane potential (Fig. 3). Notably, however, a 3 hours prematuration culture period in milrinone was sufficient for recovery of low mitochondrial activity. Generally, vitrified mature oocytes with the MII spindle require at least 2 hours recovery time after the warming process.35–37 In this case, researchers may worry about the aging effect of mature oocytes. Following ovulation, MII mature oocyte aging, a process of deterioration, occurs over time, both in vivo and in intro. Oocyte aging is associated with deleterious effects on premature cortical granule exocytosis and zona pellucida hardening 38 in critical cell cycle factor, MPF, and mitogen-activating protein kinase. 39 However, prematuration of immature oocytes is not affected by these factors. Data from our study suggest that prematuration of vitrified/warming immature oocytes in the presence of milrinone can promote recovery of mitochondrial inner membrane potential.
Finally, ROS production was decreased in prematuration cultures of immature oocytes. However, vitrified/warmed oocytes showed higher levels of ROS compared with fresh oocytes. ROS formation occurs during the intermediate steps of oxygen reduction. Mitochondrial damage during the vitrification and warming procedure may induce ROS production. Intracellular signaling controls in postovulatory oocyte aging have not been determined as yet. In many cells, oxidative stress is a major mediator of aging and disease. 40 Levels of ROS are reported to increase with oocyte aging 41 and cryopreservation.18,42 Increased ROS can lead to decreased oocyte fertilizing ability and embryonic development and promote the incidence of apoptosis in oocytes and embryos. 33 According to previous reports, recovery of ROS to physiological levels before oocyte maturation may facilitate normal oocyte maturation and further embryonic development. 33 In this study, 3 hours prematuration in milrinone after vitrification/warming successfully led to recovery of oocytes from cytoplasmic injury, including normalization of ROS and mitochondrial functional activity (Figs. 2 and 3). After recovery from cryoinjury, oocytes can commence nuclear maturation followed by cytoplasmic maturation.
In summary, we have demonstrated that vitrification of immature oocytes triggers injury in the cytoplasm, but recovery is possible by prematuration in the presence of milrinone, a phosphodiesterase inhibitor, for 3 hours. This improvement may be attributable to stabilization of mitochondrial functional activity and oxidative stress in the oocyte cytoplasm, which, in turn, assist nucleocytoplasmic maturation in vitro. Our results suggest a means to improve clinical-grade immature oocyte cryopreservation protocols that should additionally benefit cancer, polycystic ovarian syndrome, and premature ovarian insufficient patients. However, the maturation rate of vitrified/warmed immature oocytes clearly remains lower than that of nonvitrified oocytes, and therefore, further studies are required for optimization of the in vitro maturation protocol.
Footnotes
Acknowledgments
This research was supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health and Welfare, Republic of Korea (grant number: HI12C0055). and National Research Foundation of Korea grants (NRF-2017R1D1A1B03028155).
Author Disclosure Statement
No conflicting financial interests exist.