
Abstract
Currently the most popular mouse sperm freezing medium is R18SM3+MTG containing 18% raffinose, 3% skim milk, and 0.5 mM monothioglycerol (MTG), but there is no information available about whether MTG and other antioxidants can cryoprotect mouse sperm DNA integrity. It is also uncertain if sucrose can be used successfully for sperm cryopreservation. In this report we compared the cryoprotective effects of sucrose and raffinose, as well as the antioxidants MTG, reduced glutathione (GSH), and quercetin on sperm motility, DNA integrity, and fertility in the C57BL/6J mouse strain. Results show that: (1) 10% sucrose in the presence of 3% skim milk and 0.5 mM MTG (S10SM3+MTG) was as effective as R18SM3+MTG (p > 0.05) in cryopreserving sperm motility (21.0% ± 4.0% vs. 19.0% ± 3.6%), DNA integrity (8.1% ± 1.5% vs. 9.0% ± 1.5% TUNEL positive), fertilization rate (48.3% ± 7.5% vs. 45.0% ± 7.9%), and pup birth rate (36.7% ± 10.0% vs. 37.7% ± 3.5%); (2) Supplementation of freezing medium with MTG (0.5 mM), GSH (0.5 mM), or quercetin (25 μM) had a significant (p < 0.05) cryoprotective effect on sperm motility recovery rate compared to that of controls (MTG, 32.7% ± 10.7% vs. 19.4% ± 4.2%; GSH, 34.3% ± 3.7% vs. 24.5% ± 1.8%; quercetin, 36.3% ± 3.3% vs. 25.1% ± 3.6%); and (3) Cryopreservation significantly increased sperm DNA fragmentation level (16.4% ± 2.3% in R18SM3 and 14.6% ± 2.5% in S10SM3) compared to that of fresh sperm (3.0% ± 2.0%); however, supplementation with MTG (0.5 mM), GSH (0.5 mM), or quercetin (25 μM) significantly (p < 0.05) decreased the sperm DNA fragmentation level (MTG, 8.1% ± 1.5%; GSH, 9.0% ± 1.5%; and quercetin, 8.3% ± 1.3%). It was concluded that sucrose can be used as effectively as raffinose for mouse sperm cryopreservation, and supplementation of MTG, GSH, or quercetin at an appropriate concentration can help cryoprotect sperm motility and DNA integrity.
Introduction
S
Unfortunately, the R18SM3 freezing medium is not effective for cryopreservation of sperm from many inbred mouse strains, especially the C57BL/6 strain in which the genetic background of most genetically modified mouse lines is developed.5,6 Cryopreservation using R18SM3 not only significantly impairs mouse sperm motility but also damages DNA integrity, which significantly decreases the rate of in vitro fertilization (IVF) and embryo development.7–9
In 2001, a freezing medium containing 0.3 M (10%) sucrose, a common and cheap nonreducing disaccharide, and 3% skim milk (termed S10SM3 in this report) was reported to be as effective as R18SM3 in cryopreserving mouse sperm motility 10 ; however, further research is needed to confirm the results. It is also uncertain if sucrose can be used successfully for cryopreservation of sperm DNA integrity and fertility in mice.
Mechanisms of sperm cryodamage are probably multifactorial, including cold shock, osmotic stress, intracellular ice formation, and excessive production of reactive oxygen species (ROS).11–13 In recent years, numerous studies in humans and domestic animals have demonstrated the beneficial effects of antioxidant supplements on protecting sperm motility, DNA integrity, and fertility from cryodamage related to oxidative stress during cryopreservation, for example, reduced glutathione (GSH),14–16 melatonin,17,18 quercetin,19,20 vitamin E, and its analogs.21–23 The post-thaw fertilization rate of C57BL/6 mouse sperm was also significantly improved using R18SM3 supplemented with antioxidant monothioglycerol (MTG) for sperm cryopreservation.6,24 In addition, R18SM3 supplemented with antioxidant melatonin was reported to cryoprotect sperm motility of an outbred mouse strain, the Kunming mouse. 25 However, there is no information available about whether antioxidants cryoprotect mouse sperm DNA integrity and if other antioxidants are beneficial compared with MTG to mouse sperm cryopreservation.
Thus, the objectives of the present study were to compare the cryoprotective effects of sucrose and raffinose, as well as the antioxidants MTG, GSH, and quercetin on sperm motility, DNA integrity, and fertility in the C57BL/6J mouse strain.
Materials and Methods
Materials
C57BL/6 mice were obtained from the Animal Center, Xinxiang Medical School. The mice were kept at a temperature of 22°C with light cycles of 14 hours light and 10 hours dark. Mice were provided food and water ad libitum and used when they were 12 to 16 weeks old for males and 3 to 5 weeks old for females. Euthanasia was performed by cervical dislocation. The care, use, and disposition of all mice used in this study were reviewed and approved by the Ethics Committee of Xinxiang Medical University, and the animal experimentation was carried out in accordance with the institutional and national guidelines for the care and use of laboratory animals. All chemicals and reagents were purchased from Sigma-Aldrich Co. (Shanghai, China) unless otherwise stated. HTF, M2, and MBCD media were prepared according to published formulas.26–28 A stock solution of quercetin was dissolved in dimethyl sulfoxide (DMSO). GSH and MTG stock solutions were prepared in M2 medium.
Sperm cryopreservation
Sperm freezing media R18SM3 (18%, w/v, raffinose pentahydrate and 3%, w/v, skim milk, 470–490 mOsm/kg) and S10SM3 (10% sucrose and 3% skim milk, 456–465 mOsm/kg) were prepared by dissolving the two components (raffinose or sucrose and skim milk) in cell culture grade water at 60°C followed by centrifugation at 15,000 g for 2 hours and filter sterilization through a 0.2 μm pore-size membrane.
To compare the effects of sucrose and raffinose in their ability to cryopreserve sperm motility and fertility in the experiments performed in Figure 2, one side cauda epididymis of each of three to four male mice was dissected out and pooled together in 1 mL of freshly prepared warm S10SM3+MTG (0.5 mM), and another side cauda epididymis of each of the same male mice was dissected out and pooled in 1 mL of freshly prepared warm R18SM3+MTG (0.5 mM) to diminish individual variation. Each cauda epididymis was cleaned of fat before being placed into the freezing medium. Sperm were collected from the cauda epididymides using a pair of forceps to hold the tissue and a syringe needle (26 G) to gently pierce and press and move against the tissue under a dissecting microscope. After 10 minutes of incubation at room temperature, the sperm suspension in each treatment was mixed and loaded into 1.8 mL cryovials (60 μL each) using large orifice pipet tips; then the sperm samples were cryopreserved by cooling in liquid nitrogen vapor phase (about −120°C) for 10 minutes before being submerged in liquid nitrogen. This sperm cryopreservation procedure using 3 to 4 male mice each was repeated thrice using a total of 11 males to provide enough sperm samples for the experiments. Sperm samples were stored in liquid nitrogen for at least overnight before being thawed at 37°C for 5 minutes in a water bath.
To compare the cryoprotective effects of different antioxidants (MTG, GSH, and quercetin) at different concentrations on sperm motility in the experiments performed in Figures 3–5, sperm from the cauda epididymides of five to six male mice were collected and pooled together in warm S10SM3 freezing medium (0.5 to 1 mL per male, sperm concentration 30–50 million/mL) to diminish individual variation. After incubation at 37°C for 10 minutes, the sperm suspension was mixed thoroughly and divided into 0.5 mL aliquots using a large orifice pipette tip. Immediately, each of the sperm aliquots was supplemented with different concentrations of an antioxidant by adding 2.5 μL of 200 × stock solution of each concentration to be tested to each sperm aliquot and mixing thoroughly. After 10 more minutes of incubation at room temperature, the sperm suspension in each treatment was mixed again and loaded into cryovials (60 μL each) using large orifice pipet tips; then the sperm samples were cryopreserved, stored, and thawed as described above. For testing each of the three antioxidants, the sperm cryopreservation procedure using five to six male mice each was repeated thrice. The concentrations tested for each of the three antioxidants were selected on the basis of previous studies on sperm of humans15,19,20 and mice.6,24
To compare R18SM3 and S10SM3 freezing media in their ability to cryopreserve sperm DNA and to determine if supplementation of MTG, GSH, or quercetin could cryoprotect sperm DNA in the experiments performed in Figure 6, sperm from one side of the cauda epididymides of five to six male mice were collected and pooled in freshly prepared R18SM3 freezing medium, and sperm from another side of the cauda epididymides of the same male mice were collected and pooled in freshly prepared S10SM3 freezing medium as described above. Then the sperm suspension in R18SM3 was divided into two parts, that is, R18SM3 and R18SM3+MTG (0.5 mM), and the sperm suspension in S10SM3 was divided into four parts, that is, S10SM3, S10SM3+MTG (0.5 mM), S10SM3+GSH (0.5 mM), and S10SM3+quercetin (25 μM). Each antioxidant was added by adding 200 × stock solutions and mixing thoroughly. After 10 minutes of incubation at room temperature, the sperm suspension in each treatment was mixed, cryopreserved, stored, and thawed as described above. This sperm cryopreservation procedure using 5 to 6 male mice each was repeated thrice using total 16 males.
Sperm concentration and motility assessments
Sperm concentrations were determined using a hemocytometer. To determine the percentage of motile sperm (motility), the sperm sample was diluted with warm (37°C) M2 medium (1:40) and mixed. On a warm glass slide a 15 μL drop of gently mixed sperm suspension was placed and covered with a warm cover glass (22 × 22 mm) to provide a chamber ∼20 μm deep according to the guidelines of the World Health Organization (2010), and then the slide was placed on a warm stage (37°C) of a phase contrast microscope, and the percentage of motile sperm was evaluated as soon as the contents were no longer drifting at 400× magnification. At least 300 sperm were counted per sample from viewing five or more fields. Motility recovery rate = (post-thaw motility ÷ prefreeze motility) ×100.
Determination of sperm DNA fragmentation
The mouse sperm DNA TUNEL assay was performed using a method published previously with some modifications. 29 Briefly, 30 μL of fresh sperm suspension was mixed with 70 μL of 1% low melting agarose in Ca/Mg-free phosphate-buffered saline (PBS) at 37°C by gently pipetting using a large orifice pipette tip. For frozen-thawed sperm, sperm were washed immediately by dilution in 1 mL M2 medium followed by centrifugation at 400 g for 5 minutes and resuspension in 100 μL of M2 medium. Then, 20 μL of the sperm-agarose mixture was pipetted onto a sterile glass slide precoated with 0.65% normal melting agarose and spread by covering with a 22 × 22 mm sterile coverslip. Loaded slides were placed at 4°C to solidify for 5 minutes, then the coverslip on each slide was gently removed, and the slides were immersed in PBS containing 4% β-mercaptoethanol and 1 mM EDTA for 30 minutes on ice to decondense sperm nuclei. After being washed with ice-cold PBS, the sperm on the slide were fixed in 4% (w/v) paraformaldehyde in PBS for 2 hours on ice, then washed in PBS, and then treated with ice-cold 70% ethanol for 1 hour. Before the TUNEL labeling reaction, sperm on the slide were washed in PBS and permeabilized in 0.5% Triton X-100 in PBS. Single- and double-strand DNA breaks were evaluated by the terminal deoxynucleotidyl transferase fluorescein-dUTP nick end labeling (TUNEL) assay using the in situ Cell Death Detection Kit (Roche, Mannheim, Germany). On each slide, 50 μL of DNA labeling reaction mixture solution was added and spread by covering a sterile cover glass and then incubated in a humidified incubator at 37°C for 1 hour in the dark. Negative controls lacking the TdT enzyme were run in each repetition. Slides were washed in PBS after cover glass was removed from each slide and mounted with VECTASHIELD antifade mounting medium with DAPI (4,6 diamidino-2-phenylindole) (Vector Laboratories, Burlingame, CA) to stain sperm DNA. A minimum of 200 sperm per sample was scored at 1000× magnification under a fluorescence microscope using a standard fluorescence filter set. The total number of sperm per field stained with DAPI (blue) was first counted and then the number of cells emitting green fluorescence (TUNEL positive showing DNA damage) was counted to obtain the percentage of TUNEL positive sperm (Fig. 1).
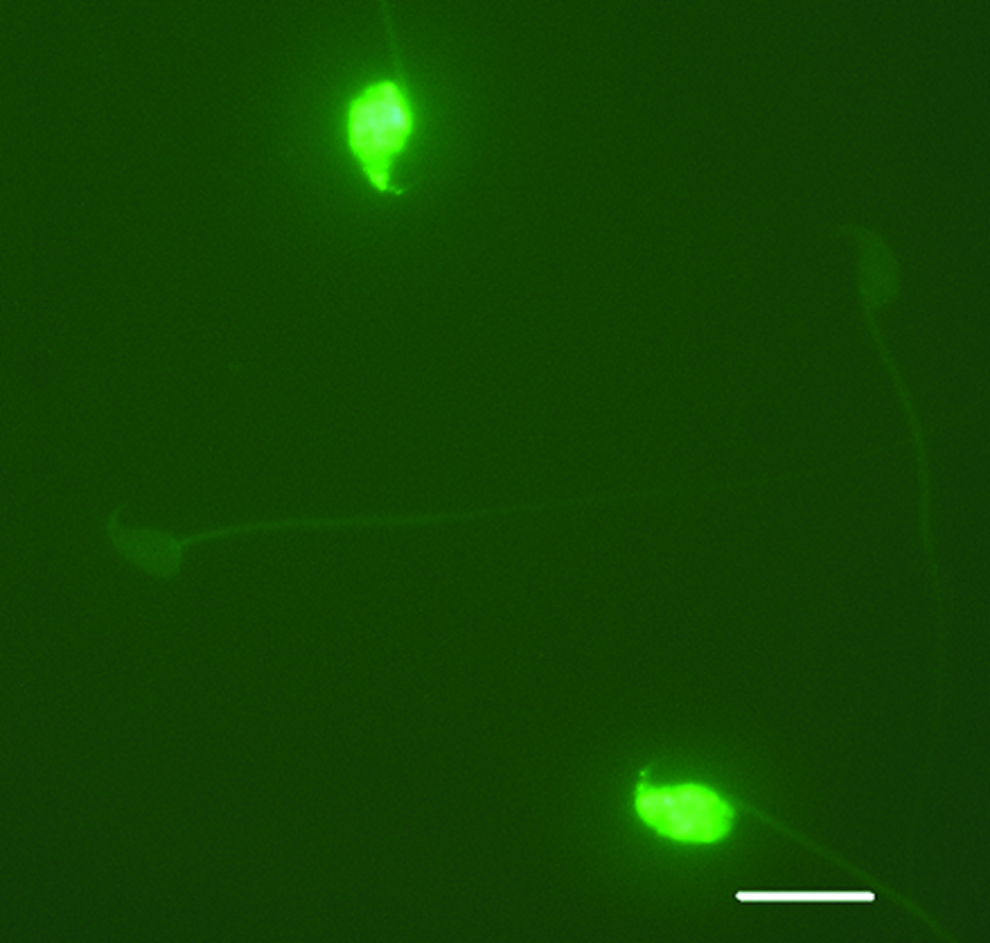
TUNEL positive and negative sperm. Original magnification 1000 × , bar = 10 μm. Color images available online at www.liebertpub.com/bio
IVF and embryo transfer
IVF and embryo transfer procedures were performed to compare the fertilization ability and developmental competence of sperm cryopreserved in S10SM3+MTG compared with that of sperm cryopreserved in R18SM3+MTG. Cumulus–oocyte complexes (COCs) were collected from superovulated C57BL/6J females 14–15 hours post human chorionic gonadotropin (hCG) injection. Superovulation was induced by intraperitoneal injection of 5 IU pregnant mare serum gonadotropin followed by 5 IU hCG 48 hours later. IVF was performed in 5% CO2 in humidified air at 37°C. A modification of a previously published MBCD-GSH IVF method was used for comparing IVF rates of sperm cryopreserved in different freezing media. 28 Briefly, for each IVF procedure sperm cryopreserved in R18SM3+MTG or S10SM3+MTG were thawed and mixed gently and then capacitated by preincubation in three pre-equilibrated 100 μL MBCD medium drops (10 μL sperm per drop) for 60 minutes before insemination. COCs from 12 superovulated females were collected, pooled, and mixed in pre-equilibrated HTF medium and then distributed evenly into two pre-equilibrated 250 μL drops of HTF medium containing 0.5 mM GSH. After sperm had been incubated for 60 minutes in MBCD medium and COCs had been incubated for 15–20 minutes, a total of 45 μL sperm were collected from the peripheral parts of the three MBCD medium drops (15 μL from each drop) for inseminating the COCs in each of the two IVF drops. After 4 hours of coculture, oocytes in each IVF drop were washed and incubated in HTF medium overnight until the morning of the next day. The numbers of 2-cell embryos and 1-cell oocytes were counted, and the IVF rate (2-cell rate) was calculated for each sperm freezing medium.
IVF-derived 2-cell stage embryos were transferred into the oviducts of 0.5 days postcoitum pseudopregnant ICR recipient female mice (10 for each of the 2 oviducts per recipient) anesthetized with 2.5% (w/v) tribromoethanol (Avertin) by intraperitoneal injection at a dose of 250 mg/kg. Recipients were kept warm on a heating pad until fully recovered from anesthesia. All pregnant recipients were allowed to give birth.
Statistical analysis
GraphPad Prism software (GraphPad Software, Inc., San Diego) was used for statistical analysis. Sperm motility, motility recovery rates, percentages of TUNEL positive sperm, fertilization rates, and embryo transfer pup birth rates were arcsine transformed, and then group differences were detected by both one-way ANOVA followed by Tukey HSD tests and paired and one-tailed t-tests, and p < 0.05 was considered significant. Data are expressed as mean (M) ± standard deviation (SD). All experiments were repeated three times (n = 3).
Results
Comparison of sucrose and raffinose in cryoprotecting sperm motility and fertility
To compare the cryoprotective effects of sucrose and raffinose on sperm motility and fertility, sperm from the same males were cryopreserved in S10SM3 and R18SM3, respectively, supplemented with 0.5 mM MTG in each treatment, and then the fertilization ability and embryo developmental potential of frozen–thawed sperm were tested by IVF followed by embryo transfer. The results summarized in Figure 2 indicate that sperm cryopreserved in the two freezing media (S10SM3+MTG and R18SM3+MTG) were not significantly different (p > 0.05) in post-thaw motility (21.0% ± 4.0% vs. 19.0% ± 3.6%), IVF rate (48.3% ± 7.5% vs. 45.0% ± 7.9%), and pup birth rate (36.7% ± 10.0% vs. 37.7% ± 3.5%).
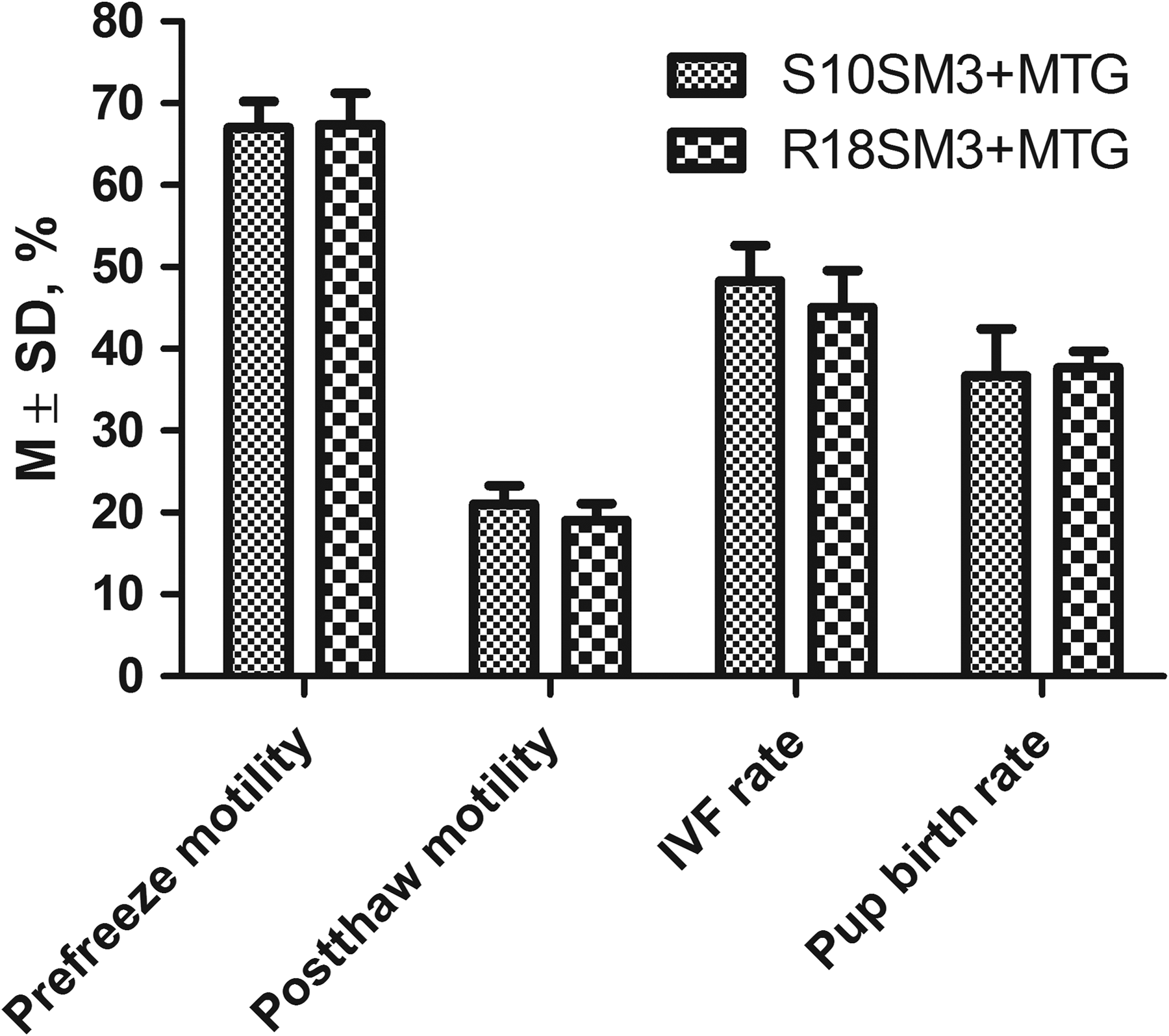
Comparison of S10SM3 and R18SM3 in cryoprotecting sperm motility, IVF rate, and pup birth rate, n = 3. Data within each group of columns are not statistically different (p > 0.05). IVF, in vitro fertilization.
Cryoprotective effects of antioxidants on sperm motility
MTG: As shown in Figure 3, MTG at 0.4 and 0.5 mM (motility recovery rate 27.4% ± 6.8% and 32.7% ± 10.7%, respectively) was significantly effective in protecting sperm motility during cryopreservation (p < 0.05) compared to the control (Ctrl, 19.4% ± 4.2%), although the difference in motility recovery rate between 0.4 and 0.5 mM of MTG was not significantly different (p = 0.096). At 0.75 mM (25.1% ± 7.0%) and 1.0 mM (20.1% ± 2.2%), MTG had no cryoprotective effect on sperm motility (p > 0.05).
GSH: As shown in Figure 4, GSH at 0.5 and 1.0 mM (motility recovery rate 34.3% ± 3.7% and 30.9% ± 2.5%, respectively) significantly protected sperm motility during cryopreservation compared to the control (24.5% ± 1.8%, p < 0.05), and the highest post-thaw sperm motility recovery rate was obtained using GSH at 0.5 mM (0.5 mM vs. 1.0 mM, p = 0.03). The sperm motility recovery rates obtained using GSH at 1.5 mM (25.0% ± 3.1%) and 2.0 mM (19.6% ± 1.5%) were not significantly different from that of the control (p > 0.05).
Quercetin: As shown in Figure 5, quercetin at 25 μM (motility recovery rate 36.3% ± 3.3%) had a significant protective effect on sperm motility compared to the control (25.1% ± 3.6%, p = 0.0001), but higher concentrations 50 μM (16.6% ± 2.6%) and 100 μM (11.42% ± 2.2%) significantly inhibited sperm motility (p < 0.05 compared to the control).

Cryoprotective effects of MTG on sperm motility at different concentrations from 0 (Ctrl) to 1.0 mM, n = 3. *0.4 and 0.5 mM versus Ctrl, p < 0.05. MTG, monothioglycerol.
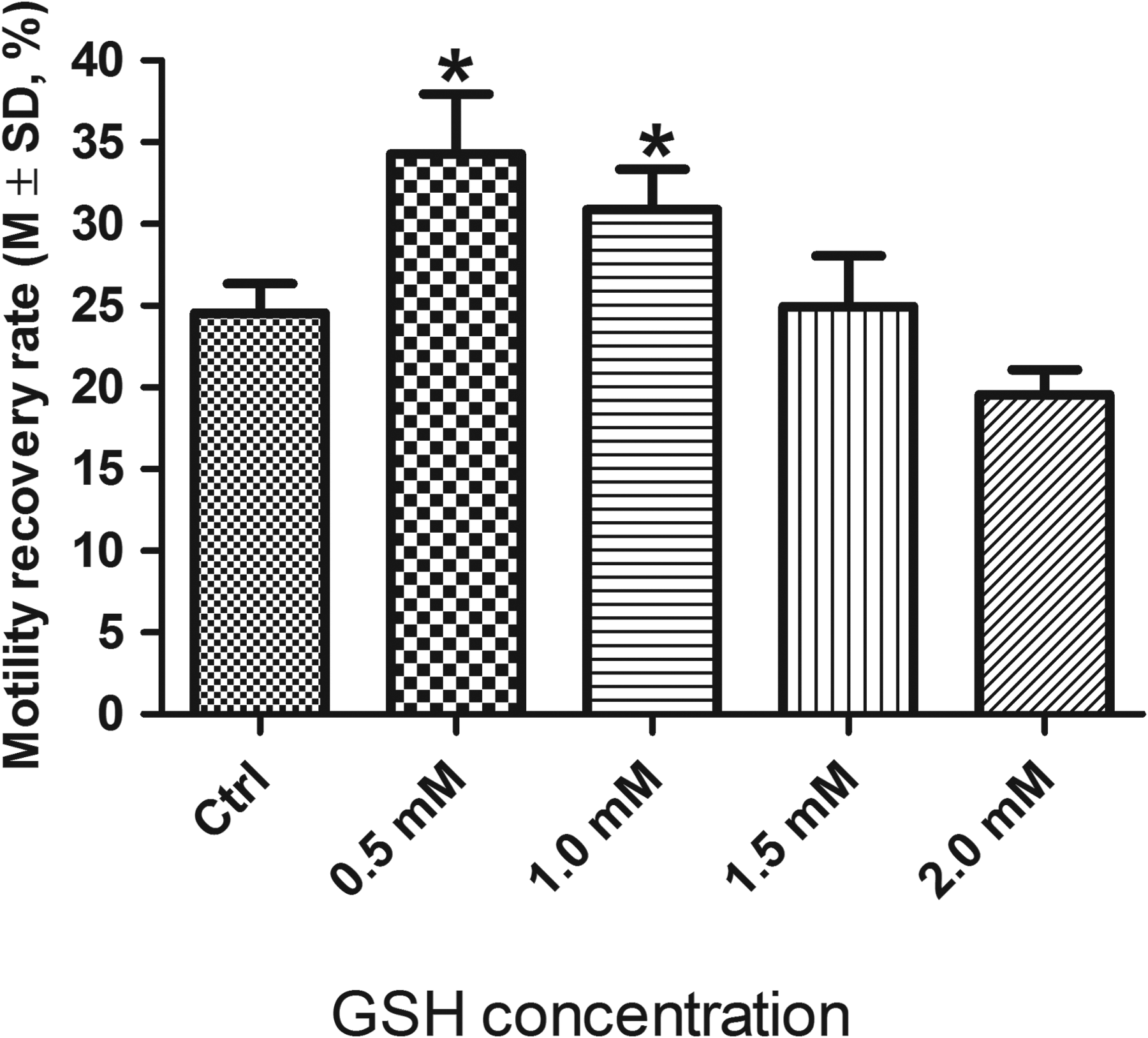
Cryoprotective effects of GSH on sperm motility at different concentrations from 0 (Ctrl) to 2.0 mM, n = 3. *0.5 and 1.0 mM versus Ctrl, p < 0.05; 0.5 mM versus 1.0 mM, p < 0.05. GSH, glutathione.
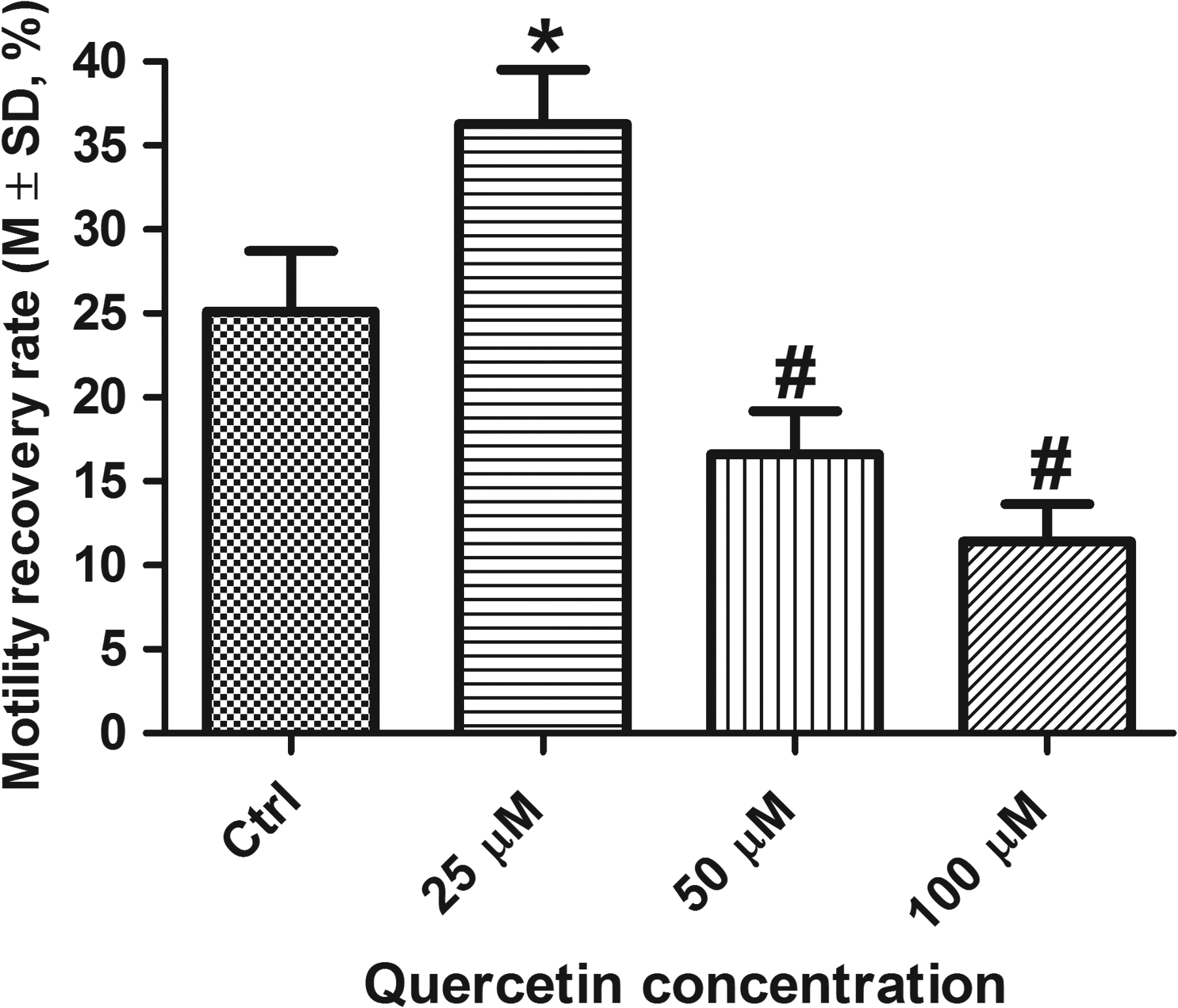
Cryoprotective effects of quercetin on sperm motility at different concentrations from 0 (Ctrl) to 100 μM, n = 3. *Significantly higher compared with Ctrl (p < 0.05); #significantly lower compared with Ctrl (p < 0.05).
Cryoprotective effect of antioxidants on sperm DNA integrity
Sperm cryopreservation experiments using R18SM3 and SM10M3, and these freezing media supplemented with an antioxidant at its optimal concentration obtained above, (MTG 0.5 mM, GSH 0.5 mM, or 25 μM) were performed to compare their cryoprotective effects on sperm DNA integrity. The results summarized in Figure 6 indicate that in fresh sperm there were 3.0% ± 2.0% of sperm with damaged DNA (TUNEL positive). Cryopreservation significantly increased the percentage of sperm with DNA damage to 16.4% ± 2.3% in R18SM3 and 14.6% ± 2.5% in S10SM3 (no significant difference between them, p > 0.05). However, supplementation with MTG (R18SM3+MTG, 9.0% ± 1.5% TUNEL positive; S10SM3+MTG, 8.1% ± 1.5% TUNEL positive no significant difference between them), GSH (9.0% ± 1.5% TUNEL positive), or quercetin (8.3% ± 1.3% TUNEL positive) in the freezing media effectively and equally protected sperm DNA integrity (p < 0.5 compared to R18SM3 or S10SM3), although the percentage of sperm with DNA damage in each of the antioxidant groups was still significantly higher compared with fresh sperm group (p < 0.05).
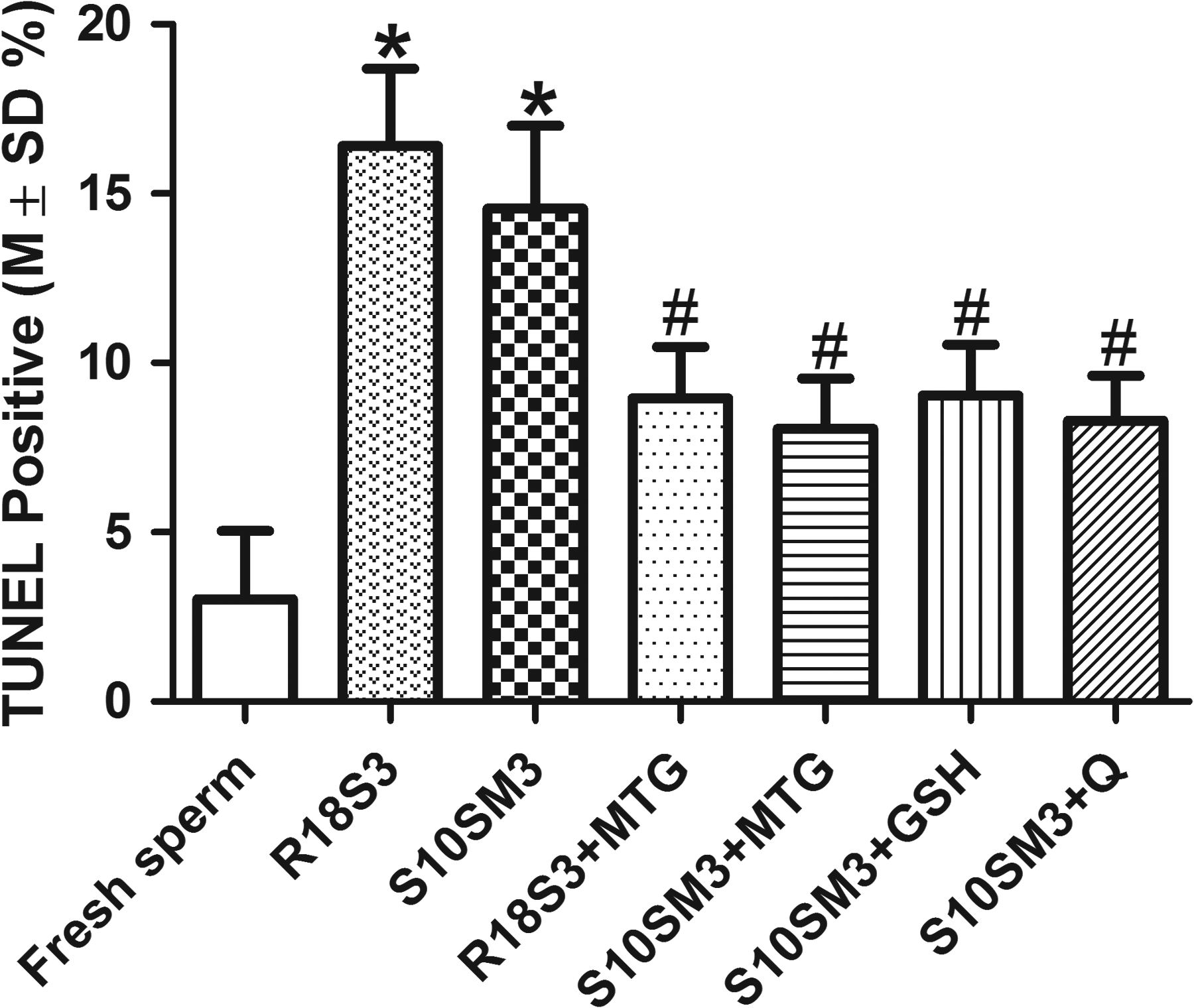
Cryoprotective effects of MTG (0.5 mM), GSH (0.5 mM), and quercetin (Q, 25 μM) on sperm DNA integrity, n = 3. *Significantly higher compared with fresh sperm (p < 0.05); #significantly higher compared with fresh sperm, but lower compared with R18S3 or S10SM3 (p < 0.05). The% TUNEL positive sperm in MTG, GSH, and Q treatments were not significantly different from each other (p > 0.05).
Discussion
Sperm cryopreservation significantly reduces motility, DNA integrity, and fertility in animals and humans. Sperm motility is associated with fertilization, and sperm DNA damage is associated with defective embryonic development and high rates of miscarriage, as well as morbidity in the offspring. 30 Therefore, more research on improving sperm cryoprotective agents and cryopreservation methods is urgently needed. The mechanisms of sperm cryodamage are probably multifactorial, but oxidative stress has been recognized as a significant cause of sperm cryodamage. 11 Sperm cryopreservation has been found to cause a decrease in the antioxidant defense system and excessive production of ROS in humans and domestic animals,12,13,15,16 and the mitochondria are the major source of ROS in sperm. 31 ROS can impair sperm motility and fertilization ability through membrane lipid peroxidation, protein thiol group oxidation, and other mechanisms.32,33 Oxidative stress also induces DNA fragmentation in the sperm nuclear genome.34,35
Numerous studies in humans and domestic animals have demonstrated the beneficial effects of antioxidant supplements, for example, GSH, melatonin, quercetin, and vitamin E, on protecting sperm motility, DNA integrity, and fertility from cryodamage related to oxidative stress during cryopreservation14–23 by reducing sperm ROS levels.15,19 The concentrations of GSH beneficial to cryopreservation of sperm motility in humans were reported to be 1 mM, 15 and the concentration of quercetin beneficial to cryopreservation of sperm motility and DNA integrity in humans was reported to be 10 and 50 μM.19,20 Antioxidant MTG at 0.477 mM (∼0.5 mM) has also been demonstrated to be effective in protecting C57BL/6 mouse sperm motility and fertility.6,24 In the present report, we found for the first time that MTG at 0.5 mM is also effective in protecting mouse sperm nuclear DNA integrity during cryopreservation. Moreover, we found that antioxidants GSH (0.5 mM) and quercetin (25 μM) are as effective as MTG (0.5 mM) in protecting mouse sperm motility and DNA integrity.
ROS have both beneficial and detrimental effects in terms of important regulatory functions, including sperm motility and fertility. The presence of appropriate antioxidant capacity in sperm freezing medium can neutralize the oxidative stress; however excessive antioxidants can stop the normal sperm functions associated with ROS.32,36 This is probably the reason we found in the present report that MTG, GSH, and especially quercetin at high concentrations have no protective effect on sperm motility or even to significantly inhibit sperm motility. Therefore, it is important to use appropriate antioxidant concentration to maintain the natural balance that exists between ROS generation and metabolic activities, which depends on species, sperm quality, and sperm concentration.
Since the first establishment of mouse sperm cryopreservation in 1990 using raffinose and skim milk,2,3 R18SM3 has been the most popular freezing medium for mouse sperm cryopreservation.4,6,10,28 In this report, we confirmed the results reported in 2001 that a combination of 10% sucrose and 3% skim milk can cryopreserve motility and fertility of C57BL/6 mouse sperm as efficiently as raffinose. 10 We also found for the first time that there is no difference in cryoprotecting sperm DNA integrity between 10% sucrose and 18% raffinose.
In conclusion, the present study demonstrated that 10% sucrose is as efficient as 18% raffinose in the presence of 3% skim milk for cryopreserving sperm motility, DNA integrity, and fertility in the C57BL/6J mouse strain. Cryopreservation leads to a significant decrease in sperm motility and a significant increase in sperm DNA damage, but the antioxidant supplements MTG, GSH, or quercetin can effectively cryopreserve sperm motility and DNA integrity.
Footnotes
Acknowledgments
This research was funded by “National Natural Science Foundation of China, Award No. U1604179” and “Program for Training Young and Key Teachers in Colleges and Universities in Henan Province, China, Award No. 2016-GGJS-220.”
Authors' Contributions
All authors have accepted responsibility for the entire content of this submitted article and approved its submission.
Author Disclosure Statement
No competing financial interests exist.