
Abstract
The cryotolerance of farm animal spermatozoa varies according to their specific features, such as size, shape, and lipid composition. Thus, it is impossible to develop a standardized freezing procedure for different kinds of livestock species. The establishment of an efficient semen cryopreservation procedure will facilitate long-term conservation of small ruminant genetic resources and extension of artificial insemination in daily production. Different from sheep, goat seminal plasma contains a phospholipase, which can affect spermatozoa viability through interaction with milk or egg yolk. Currently, soybean lecithin is a viable alternative that replaces the components of animal origin in freezing extenders for goat semen. In addition, vitrification or freeze-drying may act as another alternative to replace traditional cryopreservation. However, these two methods, especially freeze-drying, may require the aid of the intracytoplasmic spermatozoa injection technology. Furthermore, the cryoinjury mechanism of mammalian spermatozoa has remained unclear until now. The emergence of proteomics and transcriptomics may provide some inspiration concerning this problem. In this review, we summarize the state of art relating to small ruminant semen cryopreservation, mainly focusing on the current status of the freezing procedures. In the meantime, some highlights such as protectants, vitrification, and freeze-drying are also reviewed. Finally, the future perspectives in the field of small ruminant spermatozoa preservation are discussed.
Introduction
As an important part of modern livestock industry, small ruminants, including sheep and goat, can provide meat, wool, skin, or milk for our society. However, with the fast development of modern intensified agriculture, the genetic diversity of farm animal species is rapidly being reduced in many regions worldwide. 1 According to the Second Report on the State of the World's “Animal Genetic Resources for Food and Agriculture” (2000–2014), ∼99 farm animal species have disappeared from the planet forever. About 17% of livestock breeds are on the brink of extinction. 2 Currently, it is believed that small ruminant species are facing more serious risk of extinction, mainly due to the commercial extension of high productive breeds derived from high-intensity artificial selection and breeding. Therefore, to conserve the genetic diversity of small ruminant is becoming a necessity.
At present, the establishment of a living conservation herd is a routine strategy for protecting the genetic resources of domestic animal species. 3 However, this method requires a significant investment of money and labor. To resolve these problems, some researchers have attempted to preserve male or female genetic information through cryopreservation of spermatozoa or oocytes. However, a standardized freezing procedure for mammalian oocytes has not been established because they are sensitive to cryoinjuries caused by the freezing and thawing process. By contrast, semen cryopreservation has been applied in the daily livestock industry, especially in cows. Currently, semen cryopreservation has been applied as an important strategy for establishing the genetic resource bank. Semen cryopreservation does not need to raise animals, largely reducing the cost. Furthermore, this technology can contribute to the extension of artificial insemination (AI), which is a landmark technology in the modern livestock industry. Semen cryopreservation results in AI being free of the limits of operating time or site. Now, this method has been widely used for conservation of the biological diversity of livestock.
In 1937, the early experiments on freezing of sheep semen were performed by Bernstein and Petropavlovsky. In their studies, 9.2% glycerol was used as the permeable cryoprotectant to freeze semen at −21°C. 4 Then in 1949, a milestone breakthrough in the cryobiological history occurred when Polge et al. confirmed the cryoprotective effect of glycerol on poultry semen. 5 Inspired by this finding, the effects of glycerol on goat semen were first assessed during the 1950s. 6 Another critical transition in semen cryopreservation during the 1950s is that dry ice (−79°C) was replaced by liquid nitrogen (−196°C). Spermatozoa viability at −196°C can be maintained indefinitely. However, the metabolic activity of spermatozoa does not completely halt at −79°C, which consequently injures the fertility of spermatozoa. 7 Since then, numerous studies have attempted to mitigate cryoinjuries on mammalian spermatozoa via the selection of cryoprotectants and optimization of the freezing/thawing processes. However, regardless of these discoveries, up to 50% of spermatozoa lose their viability after freezing and thawing. 8 In sheep, 40%–60% of post-thaw ram spermatozoa remain motile. However, only 20%–30% are biologically functional. 9 In addition, after cervical AI using post-thaw semen, the occurrence of embryonic mortality and abnormal spermatozoa transporting patterns through the female reproductive tract demonstrates compromised functions of frozen spermatozoa.10–12
The current cryopreservation procedure for small ruminant semen is modified from that of other farm animals, such as cattle. Interestingly, the freezing effect on goat spermatozoa may be more successful than that of ram spermatozoa. AI with post-thaw goat semen can result in satisfactory fertility if semen is injected into or through the cervix. However, the same method is difficult to obtain similar results in sheep, which may be due to the difference in the cervix anatomy between goat and sheep. 7 During freezing and thawing, spermatozoa have to face various types of stresses, such as ice formation, cold shock, chemical toxicity, osmotic stress, and oxidative stress. These stresses primarily influence the plasma membrane, consequently leading to a lower fertility.13,14 Currently, low pregnancy rates can be achieved when cervical AI with frozen/thawed semen is performed. 15 However, intrauterine AI can obtain better fertility in comparison with cervical AI, especially that a lower number of spermatozoa are required for intrauterine AI than for cervical AI.8,14 However, this method requires a specialized instrument and qualified technicians. In sheep, intrauterine AI is far more effective than cervical AI. According to the study of Maxwell in 1986, only 20 million motile post-thaw sheep spermatozoa were required to achieve fertility of more than 50%. By contrast, cervical AI is believed to need 10 times that dose. 16
The cryoinjury mechanism of spermatozoa is complicated and unclear. In addition, although the semen cryopreservation procedures have been applied in the livestock industry, their progress is tardigrade compared with embryos or oocytes. In this review, the current status of small ruminant semen cryopreservation research is summarized. Some technological highlights, which may give us some inspiration to improve current freezing procedures, are reviewed.
Current Status of Small Ruminant Semen Cryopreservation
Cryopreservation greatly prolongs the storage time of mammalian spermatozoa in vitro, primarily due to complete inhibition of metabolic activity in a frozen state. According to the report of Salamon et al., after cryopreservation for 35 years, ram spermatozoa still have a capacity to fertilize in vivo with intrauterine AI. 17 The mainstream procedures for cryopreservation of small ruminant semen are demonstrated in Figure 1.

The general procedures used for cryopreservation of small ruminant semen. The gray arrows show that the negative effects of seminal plasma should be removed during cryopreservation of goat semen. The dashed arrows represent the procedure of the one-step method.
Semen collection
Semen can be collected using artificial vagina or electric stimulation. However, it should be noted that electric stimulation may not be effective for the goat because it can alter the components of seminal plasma, consequently reducing the capability of spermatozoa to tolerate cryoinjury. 18 To guarantee the quality of collected semen, some stresses, such as cold shock or urine contamination, should be considered. Generally, the semen quality is assessed immediately after collection. The general parameters used for assessment of semen quality include spermatozoa concentration, motility, and morphological normality. In addition, the other tests, such as the thermal resistance test, acrosome and membrane integrity, hypo-osmotic swelling test (HOST), or in vitro fertilization, can be used for assessment of semen quality. 19
It is highly valuable to discover a parameter that can directly reflect the real fertility of spermatozoa. As a simple and efficient approach with good reliability and repeatability, HOST has been used to predict spermatozoa fertility. 20 However, Kasimanickam et al. hypothesized that spermatozoa DNA, mitochondrial membrane potential, and motility may be more reliable than the other parameters for assessment of spermatozoa fertility and the subsequent embryonic development. 21 In addition, some studies demonstrated a potential correlation between spermatozoa motility and fertility after AI.22,23 However, despite these attempts, it is difficult to determine spermatozoa fertility based on a single parameter at present. Except for the effects of spermatozoa quality, their fertilizing capability is also influenced by several factors, such as female fertility, estrus treatment (natural or hormone-manipulated), season of AI, and insemination site (intrauterine or cervical). 19 Considering the great value of finding a parameter to determine spermatozoa fertility, this topic is still attractive.
The general semen cryopreservation procedures
After the assessment of semen quality, sheep semen can be directly diluted using the freezing extenders based on Tris, fructose or glucose, citric acid, glycerol, antibiotic, and egg yolk. In this study, the dilution process should be prudently performed to avoid a “dilution effect.” In previous studies, the dilution rate may vary from 11- to 26-fold. 24 However, the two- to fivefold dilution rates are generally accepted for freezing sheep semen according to supplemented components in freezing extenders.4,24 In terms of the two-step method, sheep semen is first diluted by the freezing extenders containing no glycerol. Then, the diluted semen is further extended by the freezing extenders containing glycerol. By contrast, when the one-step method is used, semen is directly diluted using the freezing extenders containing glycerol before equilibration at a low temperature. The concentration generally used for is ∼4%–6%. 4
After dilution, sheep semen should be slowly cooled to 5°C during a minimum time of 1.5–2 hours, depending on the cooling rate that is applied.19,25,26 Then, chilled semen is further equilibrated at this temperature for 2–4 hours. During equilibration, spermatozoa have to adapt to a reduced metabolism. Moreover, the equilibration process makes cryoprotectants (mainly glycerol) enter cells, consequently reaching an equilibrium status between intracellular and extracellular concentrations of glycerol or other osmotically active components. 24
After equilibration, diluted semen, in pellet form (0.1–0.2 mL), can be prefrozen for 3–4 minutes on the surface of dry ice (−79°C). In addition, chilled semen can also be frozen in straws using an automatic freezing machine with a freezing rate of −8°C/min or a chilled rack. When a chilled rack is used, straws are generally prefrozen for 7–10 minutes in liquid nitrogen vapor (between −75°C and −125°C) (4–6 cm above the liquid nitrogen surface). Finally, prefrozen semen is directly plunged into liquid nitrogen.4,19 Straws are generally thawed for 30–60 seconds in a 35°C–40°C water bath. However, semen pellets can be thawed in either prewarmed thawing solution (wet thawing) or dry glass tubes (dry thawing).19,25,27
For goat semen cryopreservation, glycerol can be supplemented in one or two steps, as indicated in Figure 1. The final concentration of glycerol is generally 6%–7%. 19 Except for removal of the negative effects caused by seminal plasma, the other operations are similar to sheep semen cryopreservation. With a spermatozoa concentration ranging from 80 to 500 × 106 cells/mL, acceptable fertility can be achieved for frozen goat spermatozoa.28–30
The negative effects of egg yolk-coagulating enzyme
Different from sheep, goat seminal plasma contains a special egg yolk-coagulating enzyme (EYCE), which can compromise the viability of spermatozoa in the presence of milk or egg yolk. Now, EYCE has been identified as phospholipase A secretion from the bulbourethral gland, which can coagulate egg yolk and hydrolyze lecithin to fatty acids and spermicidal lysolecithins. 31 These hydrolysates also induce acrosomal reaction and chromatin decondensation. 30 Similarly, a 55–60 kDa glycoprotein lipase (SBUIII) was identified from goat bulbourethral gland, which can induce acrosomal reaction, subsequently injuring survival of goat spermatozoa frozen in milk-based extenders. 32 In addition, both triolein and milk triglycerides can be hydrolyzed by SBUIII to free fatty acids, which strongly damage spermatozoa motility and plasma membrane. 32 Some researchers think that EYCE and SBUIII may be the same protein.18,33 Therefore, the negative interaction between EYCE or SBUIII and egg yolk or milk should be considered during goat semen cryopreservation.
The traditional operation is to completely remove seminal plasma through centrifugation before dilution using freezing extenders.30,33 In general, goat spermatozoa are washed by centrifuging at 550–950 g for 10–15 minutes.34,35 However, although removal of seminal plasma enhances the cryosurvival of goat spermatozoa, some components naturally included in seminal plasma are also lost. It is well known that the biochemical composition of seminal plasma is complex. Some studies have demonstrated a nutritive/protective function of seminal plasma for spermatozoa. 36 Some components in seminal plasma are essential for spermatozoa metabolism, function, survival, and movement in the female reproductive tract. 36 Furthermore, the proteins presented in seminal plasma are engaged in membrane stability, 37 motility, 38 capacitation,39,40 and spermatozoa/egg fertilization.40,41 In addition, seminal plasma proteins may be involved in the extrinsic and intrinsic apoptotic pathways. 42 According to the report of Maxwell et al., the presence of seminal plasma in freezing extenders can improve membrane integrity and motility of post-thaw spermatozoa. Low-molecular-weight proteins (15–25 kDa) are likely to be associated with these improvements. Particularly, adhesin may play an important role for the functions induced by seminal plasma. 43
Another way to avoid the negative effects of EYCE or SBUIII is to use skim milk-based freezing extenders. 30 Recent studies indicate that reducing the concentration of egg yolk to 2.5% cannot damage post-thaw viability of goat spermatozoa. Therefore, freezing extenders containing low concentrations of egg yolk may be used for goat semen cryopreservation.44,45 However, the controversial results still exist. According to the report of Anand et al., 3% egg yolk presence in the freezing extender cannot improve the quality of frozen goat spermatozoa in comparison with 20% egg yolk. 46 Therefore, whether this method is valuable or not still needs further research.
The phospholipids in egg yolk, such as phosphatidylcholine, are important for cryoprotection of spermatozoa plasma membrane. 47 Therefore, some similar components such as soybean lecithin may replace egg yolk to be used for goat spermatozoa cryopreservation.48,49 Currently, commercial extenders containing soybean lecithin have been used for goat semen cryopreservation.48,49 According to the report of Vidal et al., soybean lecithin, with a concentration ranging from 0.04% to 0.16%, benefited the cryosurvival of goat spermatozoa. 50 There are few reports related to the negative effects caused by soybean lecithin at present. According to the report of Valle et al., 51 soybean lecithin may interfere with mitochondrial function in post-thaw spermatozoa. In addition, although soybean lecithin resolves the problems caused by egg yolk, such as contamination, standardization, and agglutination, it can act as the substrate for lipid peroxidation because soybean lecithin contains higher proportions of arachidonic and docosahexaenoic acids and more of the C18 unsaturated fatty acids. 52
Some Highlights Inspiring Small Ruminant Semen Cryopreservation
To optimize the cryopreservation process, some measures have been used to reduce cryoinjuries of small ruminant spermatozoa. Among these attempts, selection of cryoprotectants is one of the important aspects, including antioxidants,53–59 oligosaccharides,60–67 and natural or synthetic ice blockers.68–71 In addition, vitrification and freeze-drying have emerged as potential approaches for long-term storage of spermatozoa. The relative research may give us some inspiration on how to improve the current preservation procedure of small ruminant semen.
The myth of trehalose
The research related to the functional roles of trehalose (α-
Different from monosaccharide, trehalose cannot permeate plasma membranes. So it functions primarily as an extracellular cryoprotectant. According to the report of Crowe et al., to obtain the optimal protective effect, trehalose should be presented on both sides of the plasma membrane. 82 However, how to load trehalose into cells is a great challenge. Some technologies, such as thermotropic lipid-phase transition, 83 transfection to express trehalose in mammalian cells, 84 or microinjection of trehalose into cells, 85 have been developed to resolve this problem. However, whether these technologies can be used for introduction of trehalose into spermatozoa still needs research.
Currently, the cryoprotective mechanism of trehalose remains largely undetermined. At first, the glass transition temperature of trehalose (−30°C) is much higher than that of traditional cryoprotectants, such as ethylene glycol (−85°C) and glycerol (−65°C). 72 Therefore, trehalose may contribute to extracellular vitrification formation and reduce ice crystal production. Second, according to the “water replacement hypothesis,” trehalose may replace the water shell of macromolecules through the hydrogen bond linkage, consequently reducing cryoinjury. 82 Third, trehalose may enhance fluidity of plasma membrane and mitigate cryoinjuries caused by the freezing process. 65 Furthermore, trehalose is presumed to insert into plasma membrane and limit excessive cell dehydration, decreasing physical damage induced by acute alteration of cell volume. 86 Interestingly, some studies have demonstrated the potentially antioxidative roles of trehalose at lower concentrations during spermatozoa cryopreservation.67,87 In addition, trehalose is reported to reduce osmotic sensitivity and block acrosome reaction induced by lysophosphatidylcholine in ram spermatozoa. 88
Can synthetic ice blockers replace antifreeze proteins?
How to prevent ice crystal formation during the freezing and thawing process is an essential challenge that cryobiologists have to face. Reducing injuries caused by ice formation is critical for cryosurvival of mammalian cells. In nature, some particular proteins accumulate in some low-grade organisms and plants in extremely cold environments. Subsequent studies demonstrated that these proteins promote the tolerance of these organisms to coldness or freezing.
In 1969, DeVries and Wohlschlag discovered some interesting proteins in the blood of Antarctic fish. With the aid of these proteins, Antarctic fish can survive through some extreme cold environments. 89 Generally, these proteins were called antifreeze proteins (AFPs), which influence the process of ice formation during cooling below the bulk melting point and protect mammalian tissues or cells in various ways, such as modifying ice crystal formation,90,91 prohibiting recrystallization,92,93 and interacting with the plasma membrane at low temperature.94,95
At present, AFPs have been used for cryopreservation of spermatozoa collected from sheep, 68 bovine, 96 mouse, 97 and chimpanzee. 98 Moreover, AFPs have shown a capability to improve the post-thaw motility of chimpanzee spermatozoa 98 and acrosome integrity of sheep spermatozoa. 68 However, the disputes still exist. In mouse, AFP cannot decrease cryoinjuries in spermatozoa caused by the freezing process. 97
Currently, only three types of AFPs (AFP I, AFP III, and antifreeze glycoprotein) are commercially available. 96 Owing to the high cost and difficult availability of natural AFPs, some investigators have attempted to discover or design new ice-inhibiting cryoprotectants to substitute AFPs.99–103 Chou first designed some inhibitors of ice crystal formation. However, he chose to slightly modify existing natural AFPs and did not plan to discover or synthesize nonprotein antifreeze molecules. 99 Wowk et al. reported the enhancement of vitrification solutions by the addition of modified polyvinyl alcohol.101,103 In addition, the molecular modeling technique has been used to identify molecular conformations of some chemicals, which may complement the atomic spacing of hydrogen-bonding sites on the prism face of ice crystal and change the ice crystal shape by lattice matching with available sites on the basal plane surface of an ice crystal. 104 According to the report of Taylor et al., 1,3,5-cyclohexanetriol (1,3,5-CHD), 1,3-cyclohexanediol (1,3-CHD), and 1,4-cyclohexanediol (1,4-CHD) possess the required bond angles and distances, which may be used to change ice growth pattern. 104 Cyclohexanetriol has been found to inhibit apoptosis in sheep spermatozoa induced by the freezing and thawing process. 69 Furthermore, the inhibitory capability of 1,3-CHD may be superior to that of 1,4-CHD on ice formation. 104
The inspiration from vitrification
Although traditional cryopreservation techniques have been extensively applied for storage of small ruminant semen, they cannot completely prohibit ice crystal formation, which leads to extensive cell shrinkage and structural damage.13,105 To avoid the negative effects induced by ice crystal formation, vitrification has been recommended as an alternative method. Different from traditional freezing processes, vitrification involves a direct phase transition of aqueous solutions from the liquid state to the glassy state, not experiencing the stage of ice crystal formation.
The main features of vitrification include high cryoprotectant concentration and fast freezing/warming velocity. The combination of the two factors quickly increases the viscosity of vitrification solutions and consequently blocks ice crystal formation. Vitrification has been successfully used for mammalian embryo cryopreservation. However, direct usage of embryo vitrification procedures in spermatozoa cryopreservation is not suitable, mainly due to the high sensitivity of spermatozoa to toxic and osmotic stress. 106 In addition, embryos have a multicellular structure. Therefore, even if some cells in embryos are damaged because of cryoinjuries, the remaining live cells still can proliferate to substitute for those dead cells. 107 However, spermatozoa are single cells that lack the capability of transcription and translation, and so, they cannot recover from cryoinjury. However, considering the functional role of spermatozoa as a vehicle of male DNA, the effects of membrane and acrosome integrity may be neglected when the intracytoplasmic spermatozoa injection (ICSI) technology is used for fertilization. Actually, during ICSI, spermatozoa tails are usually cut by hand to make the performance more convenient. Therefore, the structural integrity of spermatozoa is not critical for the success of ICSI.
In comparison with traditional cryopreservation procedures, vitrification may be more advantageous in some aspects. 108 Vitrification can prevent intracellular ice formation and mitigate the detrimental effects induced by high solute concentrations during the freezing and warming processes. Another advantage is that the vitrification process is fast and only takes a few seconds. Moreover, vitrification of mammalian spermatozoa may not require the addition of egg yolk. It is well known that the main limitation of egg yolk is its undefined components, which may be the primary reason leading to variable results among different research groups. Egg yolk may also bring potential bacterial contamination and disease transmission. Removing egg yolk is more meaningful for goat semen, due to the toxic interaction between phospholipase A in seminal plasma and egg yolk. 30 In addition, glycerol may be unnecessary when vitrification is used for spermatozoa storage.109–111 Although glycerol enhances the cryotolerance of spermatozoa, it also produces potential toxic and osmotic stress on spermatozoa.
However, there are a few limits that may influence the efficiency of spermatozoa vitrification. At first, the glassy state of vitrified samples is rather fragile and easily lost. Therefore, when warming, spermatozoa may face a great risk of devitrification. Recent studies have demonstrated that damages caused by the warming process may be more severe than the freezing process.112–114 In addition, vitrified spermatozoa generally lose their motility, due to their high sensitivity to osmotic and chemically toxic stresses, and so, conventional AI may be impossible. 113
Spermatozoa vitrification is not a novel concept. The successful vitrification of frog spermatozoa was reported in 1938. 115 However, early experiments on spermatozoa vitrification only resulted in low or no cryosurvival. 116 Actually, the opinion that vitrification is not suitable for cryopreservation of mammalian spermatozoa was popular for a long time. In 2008, Isachenko et al. found that small (30 μL) droplets of human spermatozoa survived through vitrification and warming after dilution using a freezing medium containing human tubal fluid, 1% human serum, and 0.25 M sucrose. Sixty-five percent progressive motility was obtained and mitochondrial function was maintained in post-thaw spermatozoa. 116 A similar phenomenon was also found in fish semen. 117 These studies are meaningful and encouraging because they imply that semen vitrification may be possible. With the emergence and extension of the ICSI technology, spermatozoa vitrification was revived and came into favor with researchers again.
Currently, vitrification of human spermatozoa has achieved some inspiring results.109,118–121 Some researchers have attempted to vitrify pig122,123 or sheep spermatozoa.108,111 However, limited information related to vitrification of small ruminant semen is available at present. According to the report of Arando et al., although the quality of vitrified sheep spermatozoa was significantly less than that of fresh spermatozoa, sucrose can improve total motility, viability, and membrane functionality, when spermatozoa were first equilibrated at 5°C before vitrification. The authors presumed that sucrose may act as a potential cryoprotectant to vitrify sheep spermatozoa. In addition, equilibration at 5°C can mitigate the cryoinjuries caused by vitrification on sheep spermatozoa. 111 In another study, Jiménez-Rabadán et al. found that vitrification using combinations of sucrose and glycerol significantly reduced the quality of frozen sheep spermatozoa in the absence of egg yolk. On the contrary, spermatozoa vitrified with combinations of egg yolk and glycerol at the lowest concentration demonstrated acceptable viability, acrosome integrity, and DNA fragmentation index, although vitrification still severely damaged spermatozoa motility, demonstrating that egg yolk may benefit the survival of vitrified sheep spermatozoa. 108 The main limitation in these two studies is that egg yolk has proved to be helpful for cryosurvival of vitrified sheep spermatozoa. However, it is known that one of the benefits brought by spermatozoa vitrification is to avoid the negative effects caused by egg yolk.
In addition, it should be noted that vitrification may not be suitable for daily production because a small volume is a necessary condition to obtain a fast freezing/warming rate and ensure a glassy state. However, as a way to conserve farm animal genetic resources, vitrification is valuable and has great application prospects. Even though the viability of vitrified spermatozoa is poor or completely lost, an oocyte still can be fertilized with a sperm with the aid of the ICSI technology. Therefore, the studies associated with vitrification of small ruminant spermatozoa still are meaningful.
Can freeze-drying be used for storage of small ruminant semen?
The idea of preserving mammalian spermatozoa at room temperature or in a refrigerator is always attractive. Liquid nitrogen is not required for storage of freeze-dried spermatozoa, which can bring some advantages, including reduction of the storage or shipping costs and avoiding viral contamination.124,125 In comparison with cryopreservation, the freeze-drying process is more complicated, due to the additional drying or dehydration process. As indicated in Figure 2, the freeze-drying process generally includes primary and secondary drying and two phase transitions. 126 During the primary drying process, samples are first transformed from the liquid phase into ice crystals by freezing to below their eutectic temperature. Then, frozen water is evaporated as water vapor in a vacuum environment without experiencing the intermediate liquid phase. After primary drying, there is an ∼8%–10% moisture remaining in samples based on their composition. To ensure sample stability at a relatively higher temperature, such as room temperature, the remaining unfrozen bound water has to be further removed by desorption through secondary drying. At this stage, samples are heated in a lowest vacuum environment to make bound water form water vapor. Following the primary and secondary drying processes, dried samples may be stored at ambient temperature or in a refrigerator.
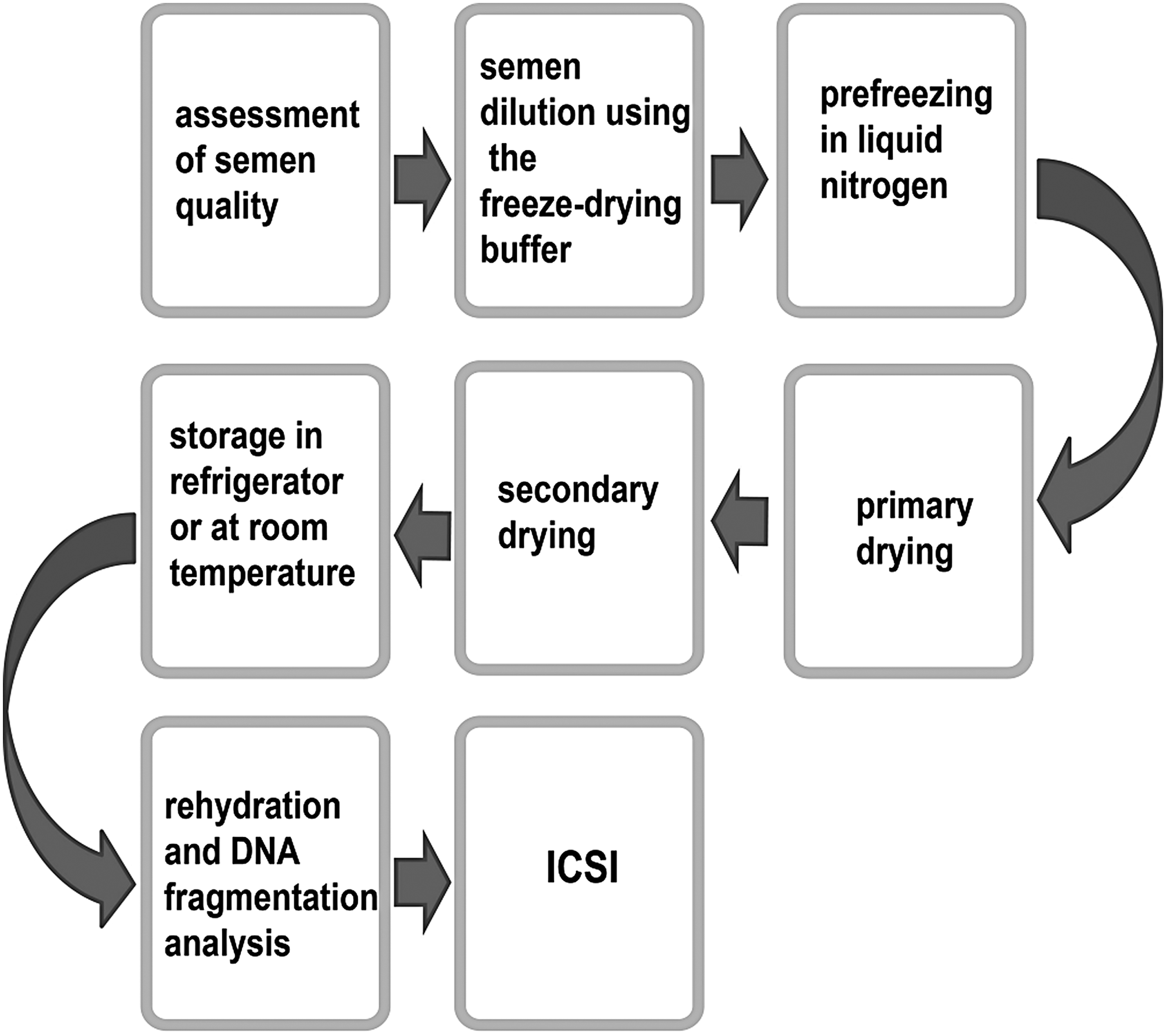
The present procedure used for freeze-drying of mammalian spermatozoa. “ICSI” is the abbreviation of the intracytoplasmic spermatozoa injection technology.
Although early studies on freeze-drying of spermatozoa showed some encouraging results,127–129 the failure to repeat these experiments made the research on freeze-drying of spermatozoa quiet for a long time. A breakthrough emerged in 1998 when Wakayama and Yanagimachi obtained healthy mice through fertilization of oocytes with freeze-dried spermatozoa. 130 The key point of their success is that the ICSI technology was used to load freeze-dried spermatozoa into an oocyte, due to the fact that all rehydrated spermatozoa were dead and lost their motility. Furthermore, the authors proved that DNA integrity and oocyte-activating factors of freeze-drying spermatozoa can be preserved for 3 months at 4°C and for several weeks at room temperature. 130 Their study demonstrated that the possession of intact genetic information is a necessary condition ensuring the success of the ICSI technology and critical for maintaining the full-term developmental capacity of freeze-dried mammalian spermatozoa.
Encouraged by the study of Wakayama and Yanagimachi, some researchers attempted to freeze-dry spermatozoa of various farm animal species, such as bovine, 131 pig,132,133 equine, 134 rabbit, 135 and sheep.2,136 In small ruminants, currently there are sporadic reports related to freeze-drying of spermatozoa. Olaciregui et al. assessed the effects of rosmarinic acid and storage temperature on DNA integrity of freeze-dried sheep spermatozoa. Meanwhile, the developmental capability of oocytes microinjected with freeze-dried spermatozoa was also tested. 136 These results demonstrated that freeze-dried sheep spermatozoa can be preserved at 4°C and room temperature for 12 months. Furthermore, rosmarinic acid can mitigate DNA damage of spermatozoa induced by the freeze-drying process. No differences were found between freeze-dried spermatozoa and frozen spermatozoa with respect to blastocyst formation. 136 Recently, Anzalone et al. further confirmed that freeze-dried sheep spermatozoa supported blastocyst development after ICSI, implying that freeze-drying is an alternative, low-cost storage strategy for biodiversity conservation. In their study, after ICSI with freeze-dried sperm, oocytes were chemically activated by ionomycin to start embryonic development. 2 The sperm-oocyte-activating-factor (SOAF), locating at plasma membrane, can activate oocytes by stimulating a Ca2+ release from ooplasmic stores. 137 Therefore, spermatozoa with damaged membrane may lose their capacity to activate oocytes after ICSI, due to lack of SOAF. 138
The resurrection of freeze-drying of spermatozoa is coupled with the development of the ISCI technology. Because freeze-dried spermatozoa completely lose their moving ability, they need the ISCI technology to realize the fertilization with oocytes. Currently, the quality of freeze-dried spermatozoa has great room for improvement. Without a doubt, the damage induced by the freeze-drying process on spermatozoa is more severe than that caused by cryopreservation, due to additional drying treatments.
Spermatozoa DNA may be degraded by DNAases or oxidative stress during the freeze-drying process. The endogenous nucleases released from freeze-dried spermatozoa may be the main reason causing chromosome structural abnormality. 139 Some measures have been used to reduce the negative effects of freeze-drying on spermatozoa DNA.139–141 Kusakabe et al. found that the chromosome integrity of mouse spermatozoa could be maintained when they were freeze-dried in a Tris-HCl-buffered solution containing 50 mM EGTA and 50 mM NaCl. 139 In addition, the stability of freeze-dried mouse spermatozoa can be improved by using a simplified Tris-buffered solution supplemented with calcium chelators, such as EGTA or EDTA.139,140 Therefore, the chelation of calcium seems to play an important role for inhibition of endonuclease activity.139,141 Furthermore, the effect of a slightly alkaline solution (pH = 8) has been proven to be superior to that of neutral or acidic (pH = 7.4–6.0) solutions for maintaining chromosomal integrity and subsequent embryonic development of freeze-dried spermatozoa. 142
Disaccharides, especially trehalose, may be helpful for chromosome stability and DNA integrity of freeze-dried spermatozoa.143–145 Some studies have demonstrated that spermatozoa freeze-dried in the presence of trehalose can fertilize with oocytes.143,145 In addition, oxidative stress can lead to spermatozoa DNA fragmentation during freeze-drying and rehydration.139,146 Exposure of DNA to reactive oxygen species leads to various chemical alterations, such as crosslinking, base modification, and DNA strand breakage. 147 Since DNA integrity can be influenced by oxidative stress, whether antioxidants can reduce oxidative stress on freeze-dried spermatozoa may be an attractive topic for research. 148
The primary purpose of spermatozoa preservation is to conserve complete male genetic information. Considering this aspect, freeze-drying may act as a potentially alternative scheme of traditional cryopreservation processes. Cryopreservation requires liquid nitrogen to block spermatozoa metabolism. The continuous addition of liquid nitrogen and mechanical maintenance of freezing equipment are critical for long-term cryopreservation. Once liquid nitrogen provision is interrupted, frozen spermatozoa will lose their viability, especially during some unexpected disasters, such as earthquakes or typhoons. 149 Different from cryopreservation, the storage requirements of freeze-drying are simple and easily controlled. However, the storage stability of freeze-dried spermatozoa may be shorter than that of cryopreservation, mainly due to the higher storage temperature.
Perspectives
Although the research on cryopreservation of small ruminant semen has lasted for about 60 years, using current methods 50% of post-thaw spermatozoa still lose their viability. A standardized cryopreservation process has not been established although some measures, including specific cryoprotectants, have been applied to improve the survival and fertility of frozen small ruminant spermatozoa.
Factors leading to spermatozoa cryoinjury are complicated and have not been determined so far. The extensive and severe cryoinjuries on the structure and physiological function of small ruminant spermatozoa have been confirmed. According to the results obtained by transmission electron microscopy, the plasma membrane is extremely sensitive to cryodamage, followed by acrosome. Currently, the cryoinjury mechanism of mammalian spermatozoa is a hot topic in the field of cryobiology and reproductive science. The technology of either cytology or thermophysics has been used to explore this problem. However, these methods still cannot explain the nature of spermatozoa cryoinjury. The functional role of spermatozoa is regulated by proteins and nucleic acid. Therefore, a molecular explanation may be necessary. At present, the fast development of the high-throughput sequencing technology has provided an opportunity for us to answer this problem based on RNA or proteins. In addition, some potential biomarkers obtained by omics analysis can be used for assessment of spermatozoa quality.
As alternative strategies of traditional cryopreservation, vitrification and freeze-drying have been used for storage of small ruminant spermatozoa. These two methods have some advantages compared with conventional cryopreservation, such as no ice crystal formation and storage at room temperature. However, their shortages are also evident. For example, vitrified or freeze-dried spermatozoa completely lose their mobility, so they need the ICSI technology to help them complete fertilization with oocytes. In this case, how to protect the structural and functional integrity of male DNA is essential.
Research related to cryopreservation of small ruminant semen has progressed greatly in recent years. However, significant room for improvement still exists. The current technological advances, such as vitrification, freeze-drying, and high-throughput sequencing technologies, can provide a new visual angle to improve the conservation efficiency of small ruminant spermatozoa.
Footnotes
Acknowledgments
This study was funded by the National Natural Science Foundation of China (Grant No. 31560635), Applied Basic Research Project of Yunnan Province (Grant No. 2016FB042), the National Wool Caprine Industrial Technology System (Grant No. CARS-39), and Middle and Young Academic Leaders of Yunnan Province (Grant No. 2014HB051). We sincerely thank Dr. Wei Shen from Qingdao Agricultural University and Dr. Allai Larbi from Universitad Autonoma de Barcelona for their careful revision of this article and precious suggestions.
Author Disclosure Statement
No conflicting financial interests exist.