
Abstract
The postwarming recovery culture, as one of the steps in cryopreservation process, is directly correlated with the survival and quality of embryos. Generally, recovery medium includes undefined serum or serum components that may cause the instability of results and other problems. The objective of this study was to evaluate the effect of knockout serum replacement (KSR) as a substitute for serum during recovery culture on the development and quality of vitrified parthenogenetic porcine blastocysts. Fetal bovine serum (FBS) was used as a positive control. The expanded blastocysts on day 5 were vitrified by the Cryotop method, and recovered with 10% (v/v) KSR or 10% (v/v) FBS for 48 hours after warming. Survival and hatching rates of vitrified blastocysts were significantly increased by KSR or FBS supplementation. The vitrified blastocysts recovered in KSR or FBS exhibited significantly decreased percentages of membrane damage and apoptosis, and increased total cells. Addition of KSR or FBS during recovery culture significantly reduced reactive oxygen species levels, and improved mitochondrial activity and adenosine triphosphates content in the vitrified blastocysts. Vitrification did not affect the gene expression of PCNA, CDX2, and CPT1, but significantly increased mRNA levels of POU5F1 and uPA. KSR added to the recovery medium significantly upregulated mRNA levels of PCNA and CPT1, and downregulated POU5F1 mRNA levels. The expression levels of PCNA, CDX2, CPT1, and uPA in vitrified blastocysts were significantly upregulated by addition of FBS to recovery medium. Moreover, the BAX: BCL2L1 ratio was similar between fresh and vitrified blastocysts, and KSR or FBS supplementation had no effect on the value. In conclusion, our data showed that KSR supplementation during recovery culture can improve the development and quality of vitrified parthenogenetic porcine blastocysts. These findings provide a useful reference that KSR could be used to replace FBS as a defined serum supplement for recovery culture of vitrified blastocysts.
Introduction
The cryopreservation of embryos is a powerful biotechnological tool for germplasm cryobanking and utilization of valuable genetic resources. Porcine embryos have been more difficult to cryopreserve as compared to the embryos of other important domestic animals. 1 The major cause of poor cryosurvival is high lipid content in porcine embryos. Compared with conventional slow freezing, vitrification methods have been proven to result in higher embryo survival and reliably produced piglets after transfer.2–4 Furthermore, porcine expanded blastocysts derived from in vivo or in vitro fertilization (IVF) are often used for vitrification due to their reduced cytoplasmic lipids. 5
However, these vitrified embryos usually show observable changes in structure and function, including membrane damage, cytoskeletal fracture, DNA fragmentation, organelle dysfunction, redox disbalance, gene expression abnormality, etc., which contribute to reducing their developmental competence.6,7 The postwarming recovery culture is one of the most important steps in cryopreservation process, which influence the survival and quality of oocytes/embryos.8–10 In general, the recovery medium is supplemented with fetal bovine serum (FBS) at concentrations of 10% or 20% (v/v) for culturing postwarming porcine blastocysts.11–14 However, no report is currently available on how serum supplementation during recovery culture affects blastocyst quality and the underlying mechanisms.
FBS as a serum supplementation is used principally to provide growth factors, energy substrates, pH buffers, free radical scavengers, and heavy-metal chelators.15–17 Nonetheless, serum compositions are not entirely known and may differ among batches. There is a potential risk of disease transmission in utilization of these animal-derived serum supplements. 18 On the contrary, knockout serum replacement (KSR) is a more defined, serum-free medium supplement, which is synthetically produced by small organic molecules, trace elements, and proteins. KSR has been widely used to support the in vitro derivation of pluripotent stem cells.19,20 A recent study reported that KSR supplementation during the late in vitro culture (IVC) phase was beneficial for in vitro production (IVP) of porcine embryos, providing functionalities similar to serum. 21 Therefore, KSR may also be an appropriate supplement to replace FBS for culturing vitrified-warmed porcine blastocysts.
The IVP embryos created by artificial parthenogenesis have the same ability to undergo preimplantation and postimplantation development as in vitro-fertilized embryos. 22 These special embryos offer multiple opportunities in the research of biotechnology and biomedicine. The parthenogenetic embryos are also frequently used as a model to investigate cryopreservation protocols. On the contrary, a bank of parthenogenetic porcine embryos needs to be established for the convenience of different researching purposes and needs. So far, there have been only few reports about vitrification of parthenogenetic porcine blastocysts.23,24
Therefore, we used porcine expanded blastocysts produced by parthenogenetic activation (PA) of oocytes as a model, to evaluate whether the KSR supplementation during recovery culture can improve the ability of embryos to survive cryopreservation. FBS was used as a positive control because it has been often added to recovery medium in a conventional operation. The quality of embryos was assessed based on blastocyst cell number, membrane damage, apoptotic cell, reactive oxygen species (ROS) production, mitochondrial activity, adenosine triphosphates (ATP) content, and mRNA expression levels of genes related to developmental potential.
Methods
Unless otherwise stated, all chemicals used were purchased from Sigma-Aldrich Chemical Company (St. Louis, MO). Tissue culture medium 199 (TCM199), FBS, Dulbecco's phosphate buffered saline (DPBS), 2′,7′-dichlorodihydrofluorescein diacetate, MitoTracker™ Red CMXRos, and BODIPY-ATP were obtained from ThermoFisher Scientific (Shanghai, China).
Porcine oocyte collection and in vitro maturation
Prepubertal porcine ovaries were collected from a local slaughterhouse and transported to the laboratory within 2 hours in 0.9% NaCl (w/v) at 35°C–37°C. Follicles (3–8 mm in diameter) were punctured using a disposable syringe with 18-gauge needle, and follicular contents were allowed to sediment in a 50 mL conical tube. The precipitate was resuspended with Tyrode's lactate-HEPES-polyvinyl alcohol medium (TLH-PVA), 25 and then observed under a stereomicroscope (Olympus, Tokyo, Japan) for cumulus–oocyte complexes (COCs) collection.
Only oocytes with uniform cytoplasm and compact cumulus cells were selected and washed three times in in vitro maturation (IVM) medium. The IVM medium was TCM199 supplemented with 10% (v/v) porcine follicular fluid, 3.05 mM
Production and culture of parthenogenetic porcine embryos
For PA, oocytes were washed three times in activation medium (0.3 M mannitol, 0.1 mM MgCl2, 0.05 mM CaCl2, and 0.1% bovine serum albumin), then placed between two wires of a microslide 0.5 mm fusion chamber (model 450; BTX, SanDiego, CA) covered with activation medium, and subsequently stimulated with a direct current pulse of 1.3 kV/cm for 80 μs using a BLS CF-150/B cell fusion machine (BLS, Budapest, Hungary). After washing three times in porcine zygote medium-3 (PZM-3) 26 supplemented with 5 μg/mL cytochalasin B and 10 μg/mL cycloheximide, oocytes were transferred to the same medium for 4 hours at 39°C in an atmosphere of 5% CO2 with saturated humidity.
Finally, the presumptive PA embryos were washed three times in PZM-3 before IVC in the same medium under conditions described above. The day on which PA was performed was designated as day 0. Generally, most of parthenogenetic porcine embryos can develop into blastocyst stage on day 5. Therefore, on day 50 the expanded blastocysts of excellent or good morphology were used for vitrification.
Vitrification and warming of blastocysts
The expanded blastocysts were subjected to vitrification using the Cryotop method as previously described 27 with some modifications in a laboratory maintained at 25°C ± 1°C. All solutions for vitrification and warming were prepared using DPBS supplemented with 20% (v/v) synthetic serum substitute (Irvine Scientific, Santa Ana, CA) as the basal medium (BM). For vitrification, groups of 5–8 blastocysts were washed in BM for 3 minutes, and then pre-equilibrated with 15% (v/v) ethylene glycol (EG) for 5 minutes. The equilibrated blastocysts were exposed to vitrification solution containing 35% EG (v/v), 0.6 M sucrose, and 50 mg/mL polyvinylpyrrolidone for 20–30 seconds, and were then loaded onto the tip of the Cryotop with the minimum volume of vitrification solution. The Cryotop was immediately plunged into liquid nitrogen (LN2).
For warming, blastocysts on the Cryotop were rapidly dipped into 1.0 M sucrose for 1 minute at 42°C, and then transferred stepwise into 0.5 and 0.25 M sucrose for 2.5 minutes, respectively. After keeping for 5 minutes in BM, these blastocysts were incubated for 48 hours in recovery culture medium according to the experimental design, at 39°C in an atmosphere of 5% CO2 with saturated humidity. Based on observation under a stereomicroscope, the blastocysts with blastocoel volume of more than half of the embryo volume were considered to have survived. Furthermore, the percentages of fully expanded and hatched blastocysts were recorded.
Evaluation of membrane damage
The membrane-damaged cells of blastocysts were evaluated by the differential staining method as described previously. 28 In brief, a total of 45–50 blastocysts in each group were incubated in TLH-PVA supplemented with 10 μg/mL Hoechst 33342 and 30 μg/mL propidium iodide for 20 minutes at 39°C, and then washed three times in TLH-PVA. After being mounted on microscope slides and covered with a coverslip, the stained blastocysts were observed under a fluorescence microscope (Nikon, Tokyo, Japan) using ultraviolent light and green light, respectively. The nuclei of membrane-damaged cells could appear red and blue, respectively; while the nuclei of cells with intact membrane could only appear blue. Total cells and membrane-damaged cells in each blastocyst were counted using NIH ImageJ software.
TUNEL assay
Detection of apoptotic cells was performed using terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL) with an in situ cell death detection kit (Roche, Mannheim, Baden-Württemberg, Germany). In brief, a total of 45–50 blastocysts in each group were fixed in 4% (w/v) paraformaldehyde for 1 hour at room temperature, and then permeabilized in 0.5% (v/v) Triton X-100 for 30 minutes. After washing in DPBS containing 0.1% (w/v) polyvinyl alcohol (DPBS-PVA), the blastocysts were incubated in TUNEL reaction cocktail for 1 hour at 39°C in the dark, and subsequently counterstained with 10 μg/mL Hoechst 33342 for 10 minutes to label all nuclei. Finally, blastocysts were washed three times in DPBS-PVA, mounted on microscope slides, and covered with a coverslip.
The stained blastocysts were examined using a fluorescence microscope first with excitation at ultraviolet light, and then with blue light to obtain two images of each blastocyst, respectively. The nuclei with green fluorescence were considered as apoptotic, and all nuclei were stained blue with Hoechst 33342. The apoptotic rate was calculated as the proportion of apoptotic nuclei/total cells.
Measurement of intracellular ROS levels, mitochondrial activity, and ATP
To determine intracellular ROS levels, a group of ∼5–7 blastocysts was incubated in DPBS-PVA supplemented with 10 μM 2′,7′-dichlorodihydrofluorescein diacetate for 30 minutes at 39°C in 5% CO2 incubator. After washing in DPBS-PVA, the stained blastocysts were imaged using a confocal laser-scanning microscope (Nikon A1, Tokyo, Japan) with excitation at 488 nm. The mitochondrial activity and ATP content in blastocysts were measured using 100 nM MitoTracker Red CMXRos and 500 nM BODIPY-ATP, respectively. The same operational procedures, including incubation, washing, and imaging, were used for blastocysts in all the experimental groups. The excitation wavelength was set at 535 nm to detect mitochondrial activity or 405 nm to detect ATP content.
Each experiment was independently repeated five times. The fluorescence intensities in blastocysts were analyzed with NIS-Elements software (Nikon). The representative images of blastocysts stained with 2′,7′-dichlorodihydrofluorescein diacetate (green), MitoTracker Red CMXRos (red), and BODIPY-ATP (blue) are shown in Figure 1.

The representative images of parthenogenetic porcine expanded blastocysts in different groups stained with 2′,7′-dichlorodihydrofluorescein diacetate (ROS, in green), MitoTracker™ Red CMXRos (mitochondrial activity, in red), and BODIPY-ATP (ATP, in blue). Scale bar = 100 μm. ATP, adenosine triphosphates; ROS, reactive oxygen species; KSR, knockout serum replacement; FBS, fetal bovine serum.
Quantitative real-time reverse transcription polymerase chain reaction
Total RNA was isolated from 10 blastocysts per treatment per replicate using the Eastep® super Total RNA extraction Kit (Promega, Beijing, China) according to manufacturer's protocol. RNA concentration and purity were quantified with a Nanodrop ND1000 Spectrophotometer (Biolab). First, the cDNA strand was synthesized using GoScript™ Reverse Transcription System (Promega, Beijing, China), and these cDNAs were diluted 10 times and stored at −20°C until analysis.
Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was conducted using a CFX Real-Time PCR Detection System (Bio-Rad, Hercules, CA) with a 20 μL reaction mixture containing 2 μL cDNA, 10 μL GoTaq® qPCR Master Mix (Promega, Beijing, China), 0.4 μL forward and reverse primers, and nuclease-free water. The reaction conditions were as follows: 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds, 60°C for 1 minute, and finally a dissociation stage consisting of 95°C for 15 seconds, 60°C for 15 seconds, and 95°C for 15 seconds. Five replicates were performed for each reaction. Relative gene expression was analyzed by the 2−ΔΔCT method after normalization to GAPDH mRNA abundance, an internal control gene. The primers used in this experiment are listed in Table 1.
Primers Used for Gene Expression Analysis
F, primer forward; R, primer reverse.
Experimental design
In our experiments, on day 5 expanded blastocysts were assigned randomly to four treatment groups: fresh blastocysts cultured for 48 hours in PZM-3 (Fresh group); postwarming blastocysts cultured for 48 hours in PZM-3 (Vitrified group); postwarming blastocysts cultured for 48 hours in PZM-3 supplemented with 10% (v/v) KSR (Vitrified+KSR group); and postwarming blastocysts cultured for 48 hours in PZM-3 supplemented with 10% (v/v) FBS (Vitrified+FBS group) as a positive control. The survival, full expansion, and hatching rates were determined under a stereomicroscope, according to morphological criteria. Furthermore, we examined the membrane-damaged cells, apoptotic nuclei, and total cell numbers in blastocysts. The intracellular ROS levels, mitochondrial activity, and ATP content of blastocysts were also determined for each group. Finally, qRT-PCR was performed to analyze the mRNA expression levels of embryo quality-related genes (PCNA, POU5F1, CDX2, CPT1, uPA, and BAX: BCL2L1) in all groups.
Statistical analysis
Data were analyzed using SPSS 20.0 software (SPSS, Inc., Chicago, IL). Blastocysts were randomly distributed among treatment groups. Percentage data were arcsine transformed before analysis to ensure homogeneity of variance. All data were analyzed by analysis of variance, followed by Student–Newman–Keuls’ multiple comparison test. Results were expressed as the least-squares mean ± standard error of the mean, and p < 0.05 was considered significant.
Results
Effects of KSR supplementation during recovery culture on survival and hatching rates of vitrified parthenogenetic porcine blastocysts
As shown in Table 2, the percentages of survived and fully expanded blastocysts after 24 and 48 hours postwarming culture were lower (p < 0.05) in the Vitrified group than in all the other groups. The hatching rate of blastocysts did not differ between Fresh and Vitrified groups, and the two values were lower (p < 0.05) than those of the Vitrified+KSR and Vitrified+FBS groups. Moreover, there was a higher (p < 0.05) hatching rate in the Vitrified+FBS group than in the Vitrified+KSR group.
Effects of Knockout Serum Replacement Supplementation During Recovery Culture on Survival and Hatching Rates of Vitrified Parthenogenetic Porcine Blastocysts
Fresh: fresh blastocysts, Vitrified: vitrified blastocysts, Vitrified+KSR: vitrified blastocysts recovered with KSR, and Vitrified+FBS: vitrified blastocysts recovered with FBS. Data are expressed as mean ± SEM values.
Different superscripts in the same column indicate significant differences (p < 0.05).
FBS, fetal bovine serum; KSR, knockout serum replacement; SEM, standard error of the mean.
Effects of KSR supplementation during recovery culture on membrane damage, apoptosis, and total cell number of vitrified parthenogenetic porcine blastocysts
As shown in Figure 2A, percentage of membrane-damaged cells in blastocysts was the highest (p < 0.05) in the Vitrified group than in all the other groups. Moreover, the membrane damage rate was similar between Fresh and Vitrified+KSR groups but higher (p < 0.05) than that of the Vitrified+FBS group. The apoptotic rate in blastocysts did not differ among Fresh, Vitrified+KSR, and Vitrified+FBS groups, and the value was significantly lower than that in the Vitrified group (Fig. 2B). There were no significant differences in the total cell number of blastocysts between Fresh and Vitrified groups. However, these values were lower (p < 0.05) than those of the Vitrified+KSR and Vitrified+FBS groups (Fig. 2C).
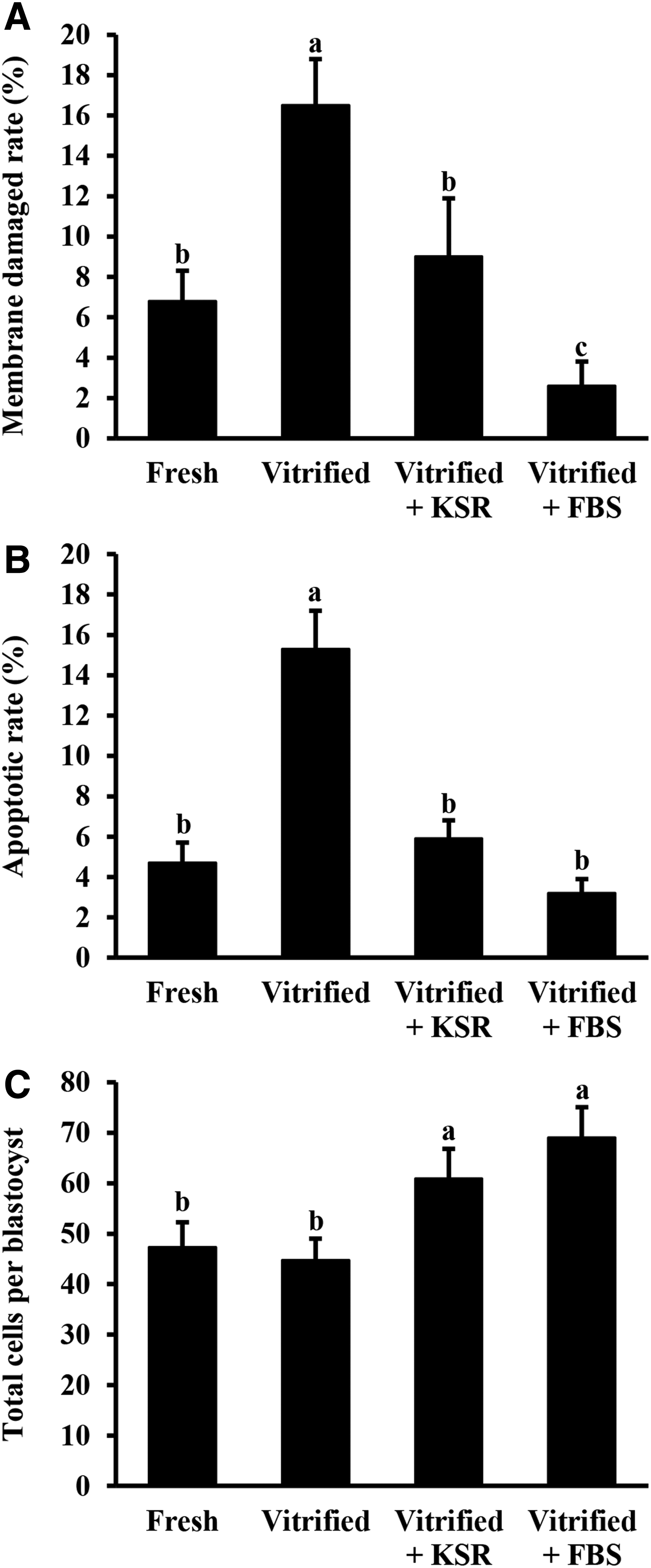
Effects of KSR supplementation during recovery culture on membrane damage
Effects of KSR supplementation during recovery culture on intracellular ROS levels, mitochondrial activity, and ATP content of vitrified parthenogenetic porcine blastocysts
There were higher (p < 0.05) intracellular ROS levels in the Vitrified group than in all the other groups. Furthermore, the ROS levels of blastocysts were similar between Vitrified+KSR and Vitrified+FBS groups, and were lower (p < 0.05) than those of the Fresh group (Fig. 3A). The mitochondrial activity of blastocysts was significantly lower in the Vitrified group compared with all the other groups (Fig. 3B). The ATP content did not differ between Fresh and Vitrified groups, and the two values were lower (p < 0.05) than those of Vitrified+KSR and Vitrified+FBS groups. Moreover, ATP content in the Vitrified+FBS group was significantly higher than that in the Vitrified+KSR group (Fig. 3C).
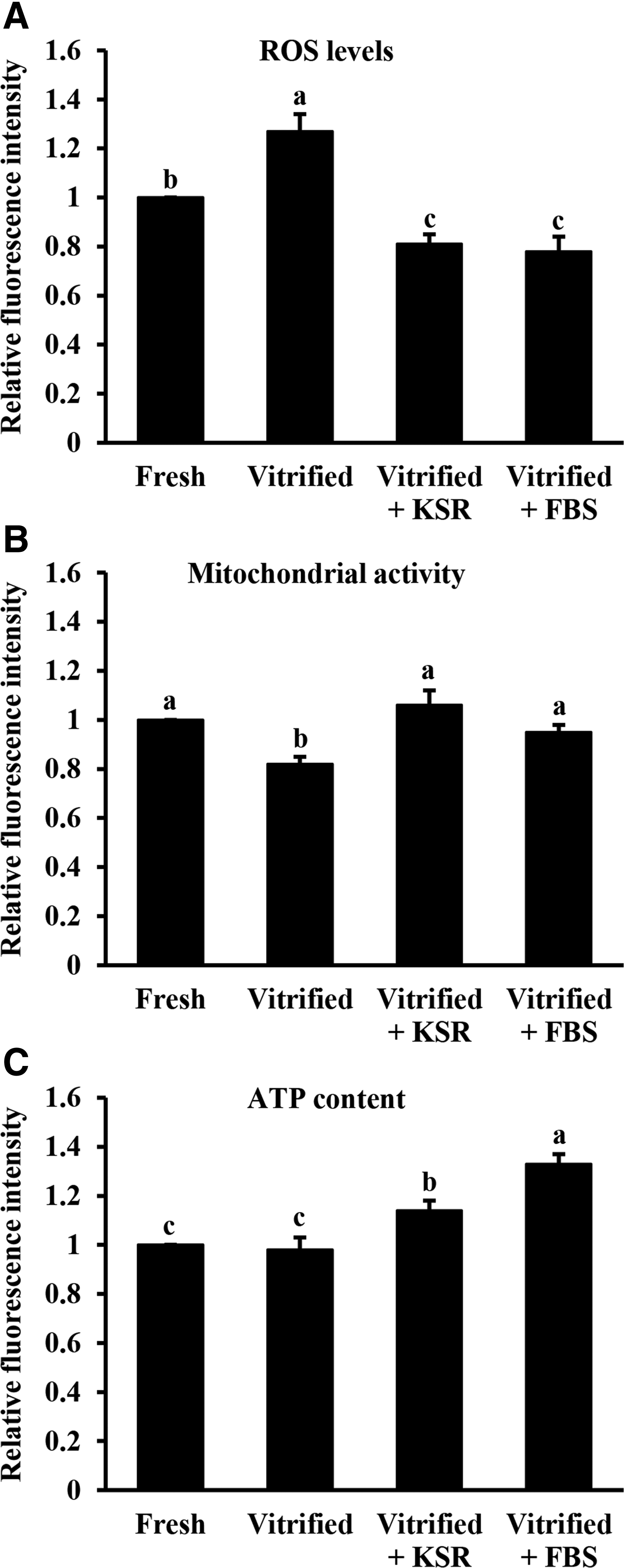
Effects of KSR supplementation during recovery culture on intracellular ROS levels
Effects of KSR supplementation during recovery culture on gene expression of vitrified parthenogenetic porcine blastocysts
As shown in Figure 4, PCNA mRNA levels were similar between Fresh and Vitrified groups, and were significantly lower than those of the Vitrified+KSR and Vitrified+FBS groups. POU5F1 mRNA levels in both Vitrified and Vitrified+FBS groups were significantly higher than those in both Fresh and Vitrified+KSR groups (Fig. 4). The mRNA levels of CDX2 did not differ among Fresh, Vitrified, and Vitrified+KSR groups, and all were lower (p < 0.05) than those in the Vitrified+FBS group (Fig. 4). The expression trend of the CPT1 gene in each group was similar to that of PCNA gene (Fig. 4). The mRNA levels of uPA were similar between the Vitrified and Vitrified+KSR groups. The values were higher (p < 0.05) than those in the Fresh group but lower (p < 0.05) than those in the Vitrified+FBS group (Fig. 4). Moreover, the BAX: BCL2L1 ratio was not different among all groups (Fig. 4).
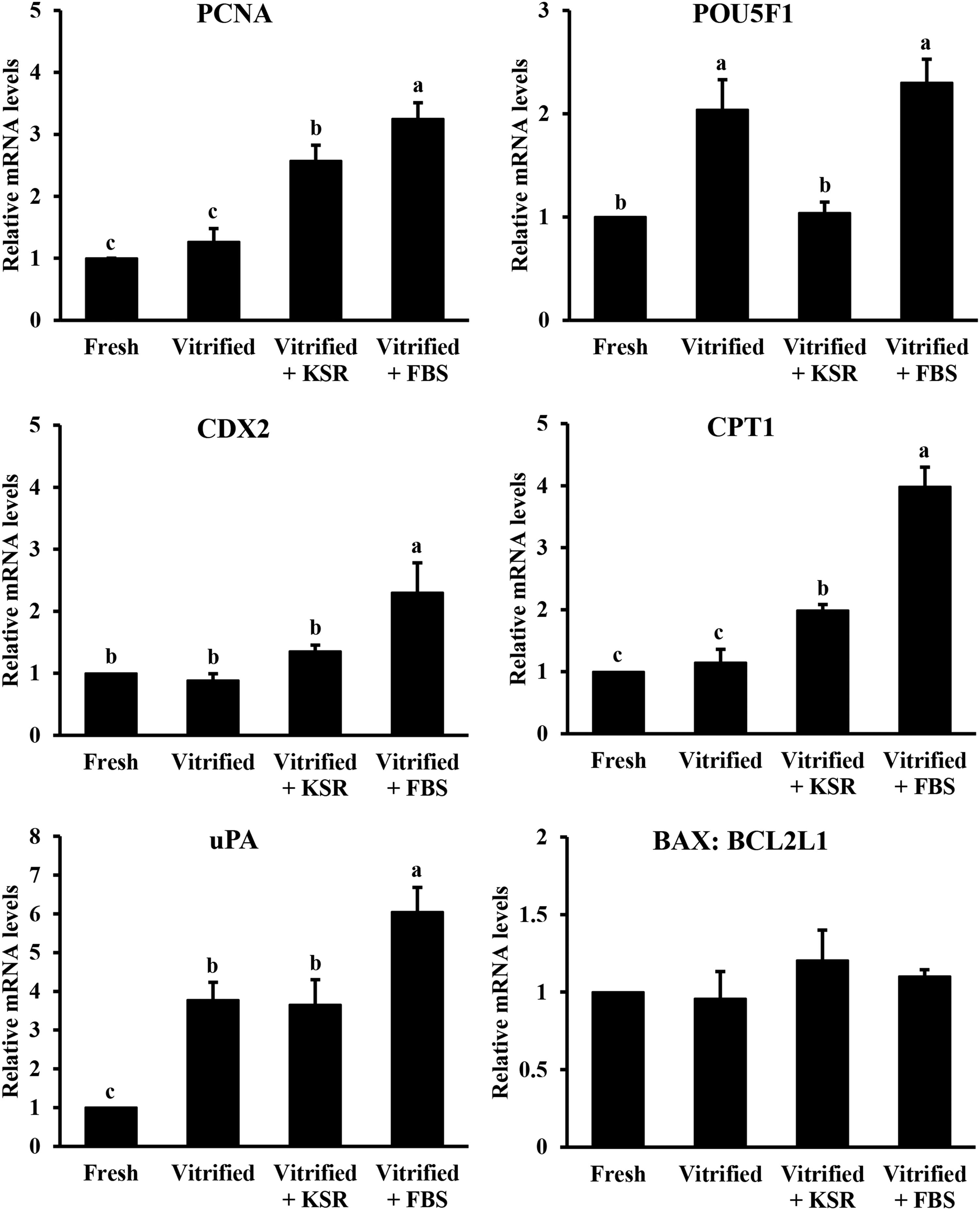
Effects of KSR supplementation during recovery culture on gene expression of vitrified parthenogenetic porcine blastocysts. Fresh: fresh blastocysts, Vitrified: vitrified blastocysts, Vitrified+KSR: vitrified blastocysts recovered with KSR, and Vitrified+FBS: vitrified blastocysts recovered with FBS. Values are mean ± SEM. Different superscripts indicate significant differences (p < 0.05).
Discussion
The Cryotop vitrification method has been recognized as one of the most efficient means for the cryopreservation of porcine oocytes and embryos, because of both high cooling (−23,000°C min−1) and warming (40,000°C min−1) rates. 27 However, the vitrification can inevitably inflict damage on blastocysts, resulting in decreased embryo survival and quality. The results of this study demonstrated the beneficial effects of replacing the FBS during recovery culture with KSR on the vitrified parthenogenetic porcine blastocysts and its probable mechanisms involved. This suggested that the defined KSR could be used for recovery of postwarming blastocysts, avoiding the interference of many uncertainties of the FBS.
In this study, KSR or FBS supplementation during postwarming culture resulted in a higher proportion of surviving blastocysts. It was reported that the porcine embryos cultured in KSR- or FBS-contained medium during the late IVC phase were beneficial for supporting their successful development, based on blastocyst yield, total cell numbers, and hatching rate.21,29,30 In this study, recovery culture of vitrified porcine blastocysts with KSR or FBS also increased the total cell numbers, and enhanced the hatching ability.
The loss of plasma membrane integrity after cryopreservation of embryos has been observed in several studies.24,28,31,32 We found that the integrity of embryo cells was more easily impaired when the vitrified blastocysts were recovered in the normal PZM-3 medium. The addition of KSR during postwarming blastocyst culture could reduce the percentage of membrane-damaged cells, and FBS supplementation played a better role in maintaining the membrane integrity. On the contrary, the incidence of apoptosis is an important parameter to reflect the embryo quality. A large number of research studies have proven that cryopreservation could induce cell apoptosis in embryos.33–36 This study showed a decrease in the number of apoptotic cells in vitrified blastocysts when KSR or FBS was added to recovery culture medium. Similarly, the presence of FBS in postwarming recovery medium also exhibited a protective effect against cell apoptosis in vitrified ovine embryos. 9
It has been reported that cryopreservation may cause a disturbance in the redox status by increasing intracellular ROS production, resulting in lower embryo quality.37–39 To overcome this, a frequent strategy was to utilize exogenous antioxidants to enhance antioxidant capacity of the vitrified embryos.40–42 In this study, the supplementing KSR or FBS in postwarming culture medium was found to reduce intracellular ROS levels of parthenogenetic porcine blastocysts. The antioxidant effect of FBS was also confirmed when it was added during the late IVC phase of fresh porcine embryos. 30
A loss of mitochondrial activity was observed in cryopreserved sheep embryos. 43 It has been reported that mitochondrial activity did not change upon vitrification in morulae, whereas it was reduced in blastocysts. 37 We also found a decrease of mitochondrial activity in the vitrified blastocysts, and supplementation of KSR or FBS to recovery medium was beneficial to improve the parameter. ATP production and accumulation in the cytoplasm is necessary for oocyte maturation and early embryo development. 44 Vitrification of oocytes and embryos has been reported to decrease their ATP content and sequential developmental competence.45,46 In our study, the vitrified blastocysts could maintain normal ATP content after 48 hours of warming. Furthermore, KSR or FBS supplementation during recovery culture was found to increase the ATP content in vitrified blastocysts.
Currently, several studies have evaluated the influence of cryopreservation on the expression pattern of embryo development-related genes in some species.35,47–50 PCNA is essential for the DNA replication and repair machinery. 51 The proportion of PCNA-positive nuclei was found to be similar between vitrified and fresh porcine blastocysts, using immunocytochemical detection. 52 In this study, the mRNA levels of PCNA in vitrified blastocysts were increased by adding KSR or FBS to recovery culture medium. POU5F1 gene has a role in pluripotency and development of preimplantation embryos. 53 We found that vitrification significantly increased the expression of POU5F1 mRNA in parthenogenetic porcine blastocysts. A similar result was also observed in vitrified mouse blastocysts.49,54 In contrast, there have been reports of a decrease in POU5F1 expression in porcine or rabbit embryos after vitrification.35,55 In addition, our study showed that KSR supplementation during recovery culture could decrease POU5F1 mRNA levels in vitrified blastocysts, but FBS did not affect this gene expression. Kazemi et al. observed that the expression of CDX2 gene involved in trophectoderm formation was increased significantly in vitrified mouse blastocysts, 56 but Zhao et al. reported no change in CDX2 expression. 57 We also found that vitrification had no effect on the mRNA expression of CDX2 in parthenogenetic porcine blastocysts. Only FBS supplementation could increase this gene expression.
In this study, the mRNA levels of lipid metabolism-related gene CPT1 in parthenogenetic porcine blastocysts were found to be unaffected by vitrification, and supplementing KSR or FBS to culture medium considerably stimulated the mRNA expression of this gene. The uPA as a candidate for zona lysis of embryos has been reported with higher mRNA levels in porcine blastocysts cultured in KSR. 21 Our results revealed that mRNA expression of uPA significantly increased in vitrified blastocysts recovered with KSR or without. The gene expression of proapoptotic BAX and antiapoptotic BCL2L1 involved in regulation of cell apoptosis during embryo development, and the BAX: BCL2L1 ratio has also been demonstrated to control the fate of cell between survival and death. 58 Currently, inconsistent results about the expression of apoptosis-related genes in vitrified embryos have been obtained in different studies.35,58–60 In this study, we found that vitrification did not affect the ratio of BAX to BCL2L1 in parthenogenetic porcine blastocysts.
In conclusion, our data showed that the supplementation of KSR similar to FBS could increase survival, hatching ability, and total cell number of vitrified parthenogenetic porcine blastocysts during recovery culture. Moreover, the quality of vitrified blastocysts was ameliorated by adding KSR or FBS to recovery medium, as evidenced by reducing membrane damage, apoptosis, and intracellular ROS levels, increasing mitochondrial activity and ATP content, and modifying some gene expression. These findings provide a useful reference that KSR could be used to replace FBS as a defined serum supplement for recovery culture of vitrified blastocysts. Still, further research is needed to confirm the beneficial effects of KSR on the vitrified embryos derived from in vivo or IVF.
Footnotes
Acknowledgments
This study was funded by the National Natural Science Foundation of China (Grant No. 31660661 and 31560636), Yunnan Applied Basic Research Projects (Grant No. 2015FD018), and the Key Laboratory Foundation of Animal Nutrition and Feed Science of Yunnan Province (Grant No. DYKF2015005).
Author Disclosure Statement
No conflicting financial interests exist.