
Abstract
Objectives:
miR-200c-3p has been shown to serve as a tumor suppressor in various tumor types. However, the biological function of miR-200c-3p in nephroblastoma remains unknown. This study aims to investigate the biological function and regulatory mechanisms of miR-200c-3p in nephroblastoma development.
Methods:
The expression of miR-200c-3p in nephroblastoma tissues and cells was evaluated by quantitative real-time polymerase chain reaction (qRT-PCR). The effects of miR-200c-3p on the proliferation and cell cycle of SK-NEP-1 nephroblastoma cell line were evaluated by CCK-8 assay, colony formation assay, and flow cytometry. The effects of miR-200c-3p on the migratory and invasive capacities of SK-NEP-1 cells were measured by wound healing assay and transwell assay. The ability of miR-200c-3p to target fibroblast growth factor receptor substrate 2 (FRS2) was detected by quantitative PCR, western blot, and luciferase reporter assay.
Results:
The expression of miR-200c-3p was significantly downregulated in nephroblastoma tissues and cells compared with that in normal renal tissues and cells. miR-200c-3p inhibited the proliferative, migratory, and invasive capacities of nephroblastoma cells by targeting FRS2.
Conclusions:
miR-200c-3p suppresses the malignant behaviors of nephroblastoma cells by downregulating the expression of FRS2.
Introduction
Nephroblastoma (Wilms' tumor [WT]) is one of the most common types of malignant renal tumor, and has a high incidence in children, especially those around 3 years of age.1,2 At present, protocols proposed by the International Society of Paediatric Oncology–Renal Tumour Study Group (SIOP–RSTG) are commonly used in the clinical treatment of WT, including preoperative chemotherapy, surgical resection, and postoperative care.3–5 Although there are studies showing that the development of WT is associated with chromosomal anomalies and mutations,6,7 the molecular etiology and regulatory elements of WT are still far from understood, which largely hinders the further development of WT therapeutic strategies.
microRNAs (miRNAs) are single-stranded RNA molecules that are ∼18–25 nucleotides in length. Although incapable of encoding proteins, miRNAs can regulate diverse kinds of physiological and pathological processes by binding to the 3′-untranslated region (UTR) of target genes, leading to their degradation or translation repression. 8 To date, numerous miRNAs have been found to participate in regulating the development of multiple types of human cancers, including WT. 9 For example, miR-1180-5p inhibited the apoptosis of SK-NEP-1 nephroblastoma cell line by targeting p73. 10 miR-185 suppressed the growth of WT by downregulating the expression of Six1 oncogene. 11 miR-204 regulated the progression of WT through targeting oncogenic transcription factor MEIS1. 12 These findings indicate that miRNAs are involved in the regulation of WT pathology.
miR-200c-3p is a cancer-associated miRNA that belongs to the miR-200 family. Studies have shown that miR-200c-3p participates in regulating diverse kinds of human cancers. It is reported that miR-200c-3p facilitated epithelial-to-mesenchymal transition in human colorectal cancer, thereby induced the liver metastasis of tumor cells. In this study, several epithelial to mesenchymal transition (EMT)-associated genes, such as ZEB1, ETS1, and FLT1, were identified to be miR-200c-3p targets. 13 In addition, miR-200c-3p was found to modulate the expression of cancer stem cells markers in colorectal cancer cells. 14 Moreover, miR-200c-3p suppresses the proliferation and metastasis of glioblastoma cells by decreasing the expression of HMGB3. 15 However, whether and how miR-200c-3p can regulate the progression of WT are largely unknown to date.
Fibroblast growth factor receptor substrate 2 (FRS2) is an oncogenic tyrosine kinase adaptor protein whose abnormal expression was reported in diverse kinds of tumors, such as bladder cancer 16 and ovarian cancer. 17 In WT, miRNA-mediated downregulation of FRS2 was shown to attenuate tumor malignancy, 18 suggesting that identifying miRNAs targeting FRS2 may become a potential strategy for the treatment of WT.
In this study, we explored the role of miR-200c-3p on the proliferative, migratory, and invasive capacities of nephroblastoma cells by modulating the expression of FRS2. Thus, our study identified miR-200c-3p as a potential tumor suppressor in nephroblastoma.
Methods
Human specimens
The specimens in this study were obtained from 32 patients who were all pathologically diagnosed with nephroblastoma at the Gansu Provincial Maternity and Child-Care Hospital. All patients who were receiving chemotherapy, radiotherapy, and/or biotherapy were excluded, and those with other types of malignant tumors were also eliminated. Specimens were collected at surgery, and were frozen immediately in liquid nitrogen then stored at −80°C for RNA extraction. The study was approved by the Hospital's Ethics Committee. Informed consent was obtained from all participants. Clinical characteristics of nephroblastoma patients are given in Table 1.
Clinical Pathological Characteristics of Patients with Wilms' Tumor
FH, favorable histology; NWTS, National Wilms' Tumor Study; UH, unfavorable histology.
Cell culture
The SK-NEP-1 cell line was obtained from ATCC (Manassas, VA) and maintained in McCoy's 5a medium containing 20% fetal bovine serum (FBS) and 1× penicillin/streptomycin. The HEK293 cell line was obtained from ATCC and cultured in Dulbecco's modified Eagle medium containing 10% FBS and 1× penicillin/streptomycin.
Cell transfection
The human miR-200c-3p mimics/inhibitor and their corresponding miRNA controls were synthesized by GenePharma (Shanghai, China). FRS2 expression plasmid was purchased from Origene (Beijing, China). Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA) was used for cell transfection according to the manufacturer's instructions.
CCK-8 assay
The CCK-8 assay was performed using Enhanced Cell Counting Kit-8 (Beyotime, Shanghai, China) following the manufacturer's instructions. In brief, 1 × 104 SK-NEP-1 cells were seeded into 96-well plates and cultured overnight. Then 20 μL CCK-8 was added to each well. Optical density values at different time points were measured at 490 nm on a microplate reader (Thermo Fisher Scientific) after 4 hours of incubation.
Colony formation assay
1 × 102 SK-NEP-1 cells were seeded in six-well plates and cultured for 15 days. Cell colonies were fixed in 70% ice-cold methanol for 10 minutes, stained with 0.5% crystal violet for another 10 minutes. Images were taken and analyzed by microscopy (Olympus, Tokyo, Japan).
Flow cytometry
Cell cycle was evaluated by flow cytometry (BDbiosciences, San Jose, CA). In brief, trypsin-digested SK-NEP-1 cells were fixed in 70% cold ethanol at 4°C overnight, followed by two washes with phosphate-buffered saline (PBS). Cell pellets were resuspended in propidium iodide solution containing RNase A and were incubated at 37°C for 30 minutes in dark. The cell cycle was analyzed on an FACS Calibur. The data were analyzed using FlowJo software (LLC, Ashland, OR).
Wound healing assay
1 × 105 SK-NEP-1 cells were seeded in 24-well plate and were cultured until nearly 100% confluence. Cells were then scraped with pipette tips and were photographed at 0, 24, and 48 hours. Three fields in each well were recorded.
Transwell assay
For migration analysis, SK-NEP-1 cells were seeded in the upper chambers of transwell inserts and were cultured in FBS-free medium, medium containing 10% FBS was added into the lower chambers. Forty-eight hours later, cells in lower chambers were stained with crystal violet and were examined under a microscope (Olympus). For invasive analysis, the upper chambers of Transwell inserts were precoated with Matrigel.
Quantitative real-time polymerase chain reaction
RNA was extracted using Trizol reagent, and complementary DNA was synthesized using the Reverse Transcription System (Kangwei, Beijing, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using SYBR qRT-PCR mixture (Kangwei) with the following parameters: 95°C 10 minutes, followed by 40 cycles of 95°C for 10 seconds and 60°C for 50 seconds. Gene expression was analyzed using the 2−ΔΔCt method. For messenger RNA (mRNA), GAPDH was used as the internal control, and for miRNA, U6 was used as the internal control. Primer sequences are given in Table 2.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Western blot
Total protein extractions from SK-NEP-1 cells were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) followed by western blot analysis as previously described. 19 In brief, SK-NEP-1 cells were washed twice with PBS and lysed by NP-40 lysis buffer. Thirty micrograms of total protein was separated through SDS-PAGE and transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA), which were then blocked with 5% nonfat milk followed by incubation with primary antibodies (1:1000 dilution). Primary antibodies against GAPDH and FRS2 were purchased from Cell Signaling Technology (Danvers, MA) and horseradish peroxidase-conjugated secondary antibodies were purchased from Beyotime.
Dual-luciferase reporter assay
miR-200c-3p, FRS2 3′-UTR luciferase reporter plasmid and control renilla luciferase reporter plasmid were cotransfected into HEK293 cells using Lipofectamine 2000 according to the manufacturer's protocol. Luciferase activities were detected by Dual-Luciferase reporter kit (Promega, Madison, WI).
Statistical analysis
Unpaired Student's t-tests were performed using GraphPad Prism 5. All experiments were conducted at least three times. Data were expressed as the mean ± standard deviation. p < 0.05 was considered to be statistically significant.
Results
Expression miR-200c-3p is downregulated in nephroblastoma tissues and cells
First, qRT-PCR was performed to detect the expression of miR-200c-3p in nephroblastoma patients. The results demonstrated that compared with tumor-adjacent normal tissues, the mRNA level of miR-200c-3p was significantly decreased in the tumor tissues of nephroblastoma patients (Fig. 1A). Furthermore, miR-200c-3p level was significantly lower in patients with advanced stages of nephroblastoma (stage III/IV) than that in patients with early stages of nephroblastoma (stage I/II) (Fig. 1B). On the contrary, miR-200c-3p expression was not associated with lymph node metastasis (Fig. 1C). Consistently, the abundance of miR-200c-3p was significantly lower in human nephroblastoma cell line SK-NEP-1 than in HEK293 cells, which represent human normal kidney cells (Fig. 1D). These results suggest that miR-200c-3p may play an important role in regulating the development of nephroblastoma.
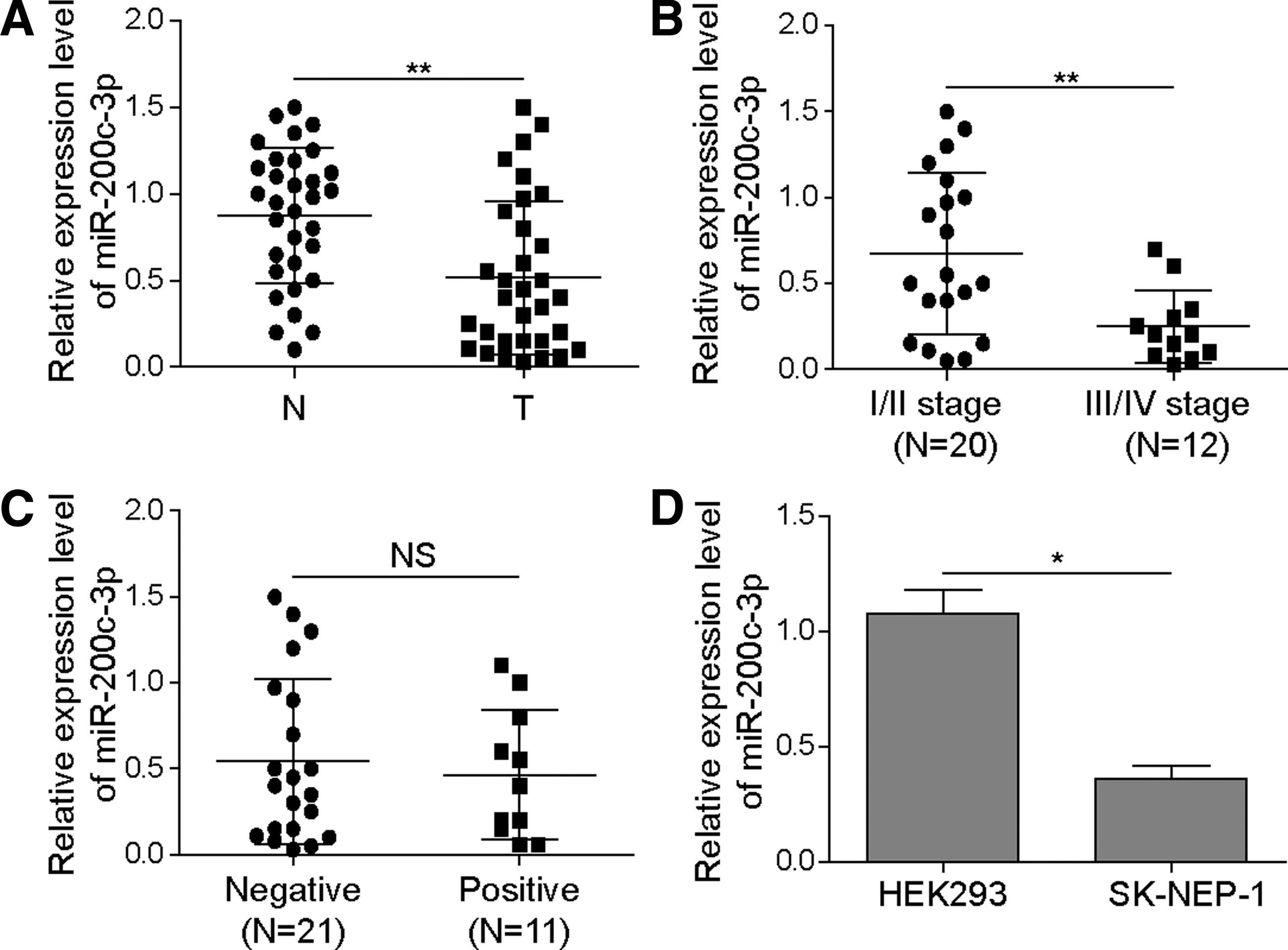
The expression miR-200c-3p is downregulated in nephroblastoma tissues and cells.
miR-200c-3p suppresses the proliferation, migration, and invasion of SK-NEP-1 cells
Next, we transfected SK-NEP-1 cells with miR-200c-3p mimics and miR-200c-3p inhibitor to further explore its possible impact on cellular behaviors of nephroblastoma cells. miR-200c-3p overexpression significantly inhibited the proliferation of SK-NEP-1 cells, which was nevertheless enhanced by miR-200c-3p inhibition, as evidenced by both CCK-8 assay and colony formation assay (Fig. 2A, B). Similarly, overexpression of miR-200c-3p led to significantly increased percentage of G0/G1 phase cells, whereas miR-200c-3p inhibition caused the opposite effect (Fig. 2C), suggesting that miR-200c-3p promoted G0/G1 cell cycle arrest in nephroblastoma cells.
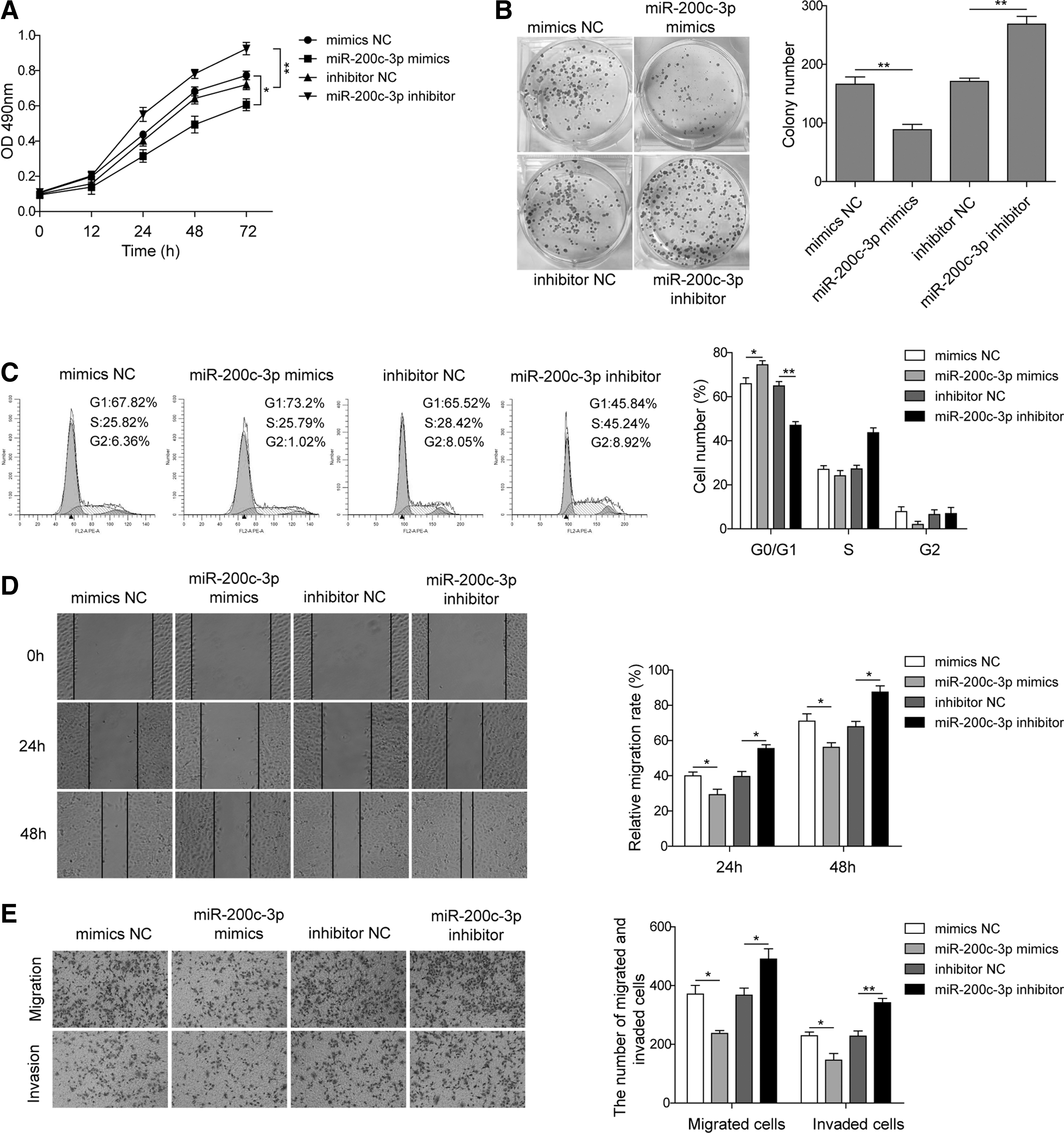
miR-200c-3p suppresses the proliferation, migration, and invasion of SK-NEP-1 cells. SK-NEP-1 cells were transfected with miR-200c-3p mimics, miR-200c-3p inhibitor or control microRNA for 48 hours, the proliferation of SK-NEP-1 cells was examined by CCK-8 assay
In a wound healing experiment, an impaired migratory ability was observed in miR-200c-3p-overexpressed SK-NEP-1 cells. In contrast, miR-200c-3p inhibition accelerated the wound closure (Fig. 2D). The transwell assay also demonstrated that miR-200c-3p impaired both the migratory and invasive capacities of SK-NEP-1 cells (Fig. 2E, F). Taken together, these results suggest that miR-200c-3p restrains multiple malignant behaviors of SK-NEP-1 cells, including cell proliferation, migration, and invasion.
miR-200c-3p regulates the expression of downstream target FRS2
We then sought to determine the molecular basis underlying the regulatory function of miR-200c-3p. Through predicting the putative targets of miR-200c-3p in Targetscan, FRS2 was identified to be potentially targeted by miR-200c-3p (Fig. 3A). This prediction was verified by the results that overexpression of miR-200c-3p significantly decreased, whereas inhibition of miR-200c-3p significantly enhanced the expression of FRS2 at both mRNA and protein levels in SK-NEP-1 cells (Fig. 3B, C). In addition, the expression of FRS2 was increased in nephroblastoma tissues compared with that in matched normal tissues (Fig. 3D). Moreover, FRS2 expression was higher in advanced stages of nephroblastoma than in early stages (Fig. 3E). In contrast, the FRS2 level was irrelevant to lymph node metastasis (Fig. 3F). Of importance, there was a negative correlation between the levels of miR-200c-3p and FRS2 in nephroblastoma tissues (Fig. 3G). Through a dual-luciferase reporter assay, we found that miR-200c-3p mimics significantly dampened the luciferase activity in SK-NEP-1 cells transfected with wild-type FRS2 3′-UTR plasmid, but not in those transfected with mutant FRS2 3′-UTR plasmid, which harbors a seed region incapable of being recognized by miR-200c-3p (Fig. 3H). Hence, miR-200c-3p can downregulate the expression of FRS2 in SK-NEP-1 cells.
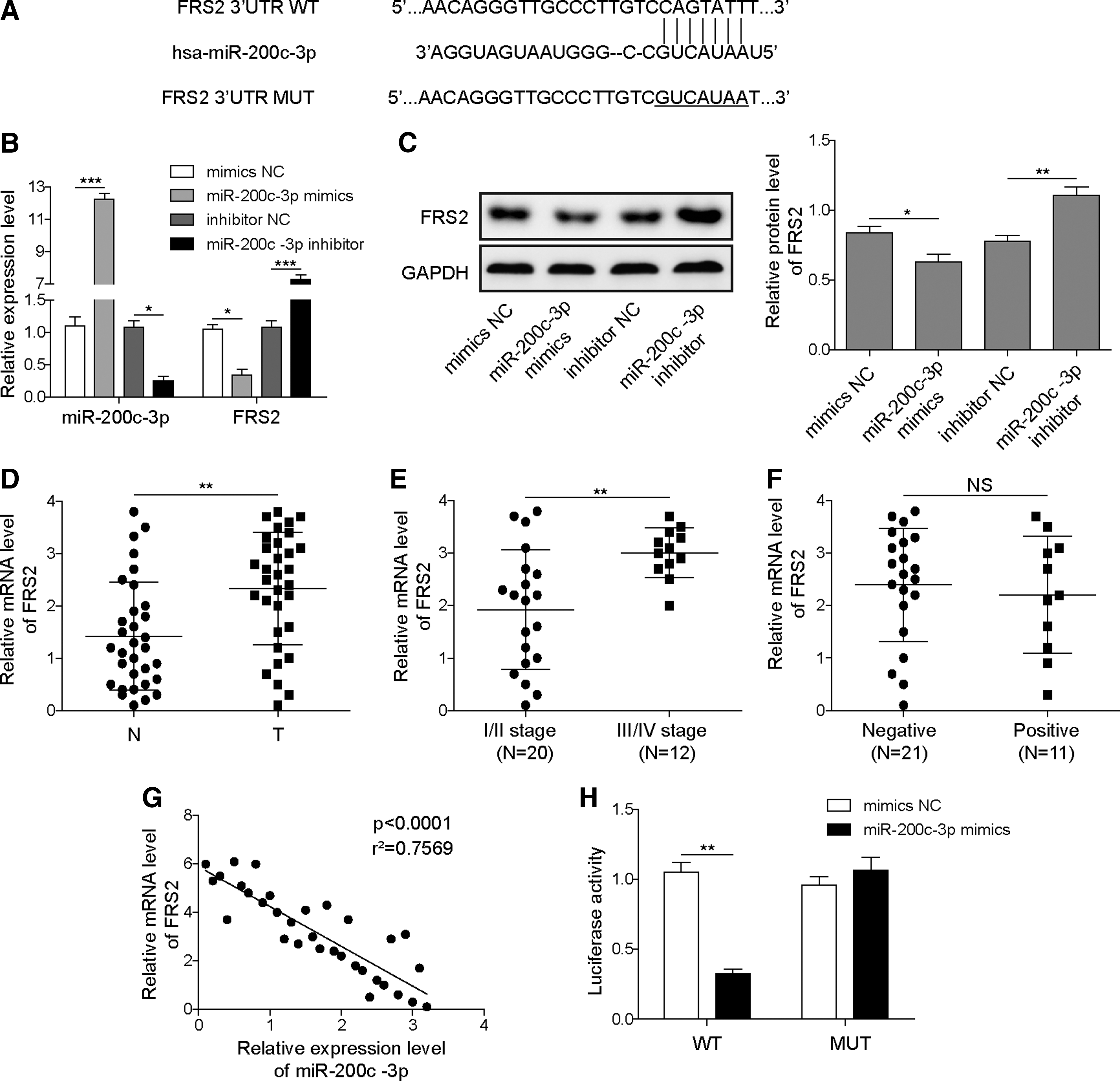
miR-200c-3p regulates the expression of downstream target FRS2.
miR-200c-3p inhibits proliferation, invasion, and migration of SK-NEP-1 cells by targeting FRS2
To further ascertain if miR-200c-3p affects the function of SK-NEP-1 cells by regulating FRS2 expression, we transfected them with miR-200c-3p mimics and pcDNA3.1-FRS2 or shFRS2. We then analyzed the functional relevance of miR-200c-3p-mediated downregulation of FRS2. As assessed by CCK-8 assay and colony formation assay, although miR-200c-3p caused growth inhibition of SK-NEP-1 cells, concomitant overexpression of FRS2 largely abrogated this effect. In contrast, cotransfection with miR-200c-3p mimics and shFRS2 had a synergistic effect in further inhibiting the growth of SK-NEP-1 cells (Fig. 4A, B). Similarly, FRS2 overexpression prevented miR-200c-3p-induced G0/G1 phase cell cycle arrest in SK-NEP-1 cells, whereas FRS2 inhibition by shFRS2 further enhanced the effect of miR-200c-3p (Fig. 4C). Through performing the wound healing assay and transwell assay, we also found that forced expression of FRS2 counteracted the effects of miR-200c-3p in reducing the migratory and invasive abilities of SK-NEP-1 cells, whereas the inhibition of FRS2 caused the opposite effects (Fig. 4D, E). As expected, miR-200c-3p mimics suppressed FRS2 expression, and this effect was reversed by FRS2 overexpression in SK-NEP-1 cells at both mRNA and protein levels (Fig. 4F, G). In summary, miR-200c-3p suppresses the proliferative, migratory, and invasive capacities of nephroblastoma cells by downregulating the expression of FRS2.
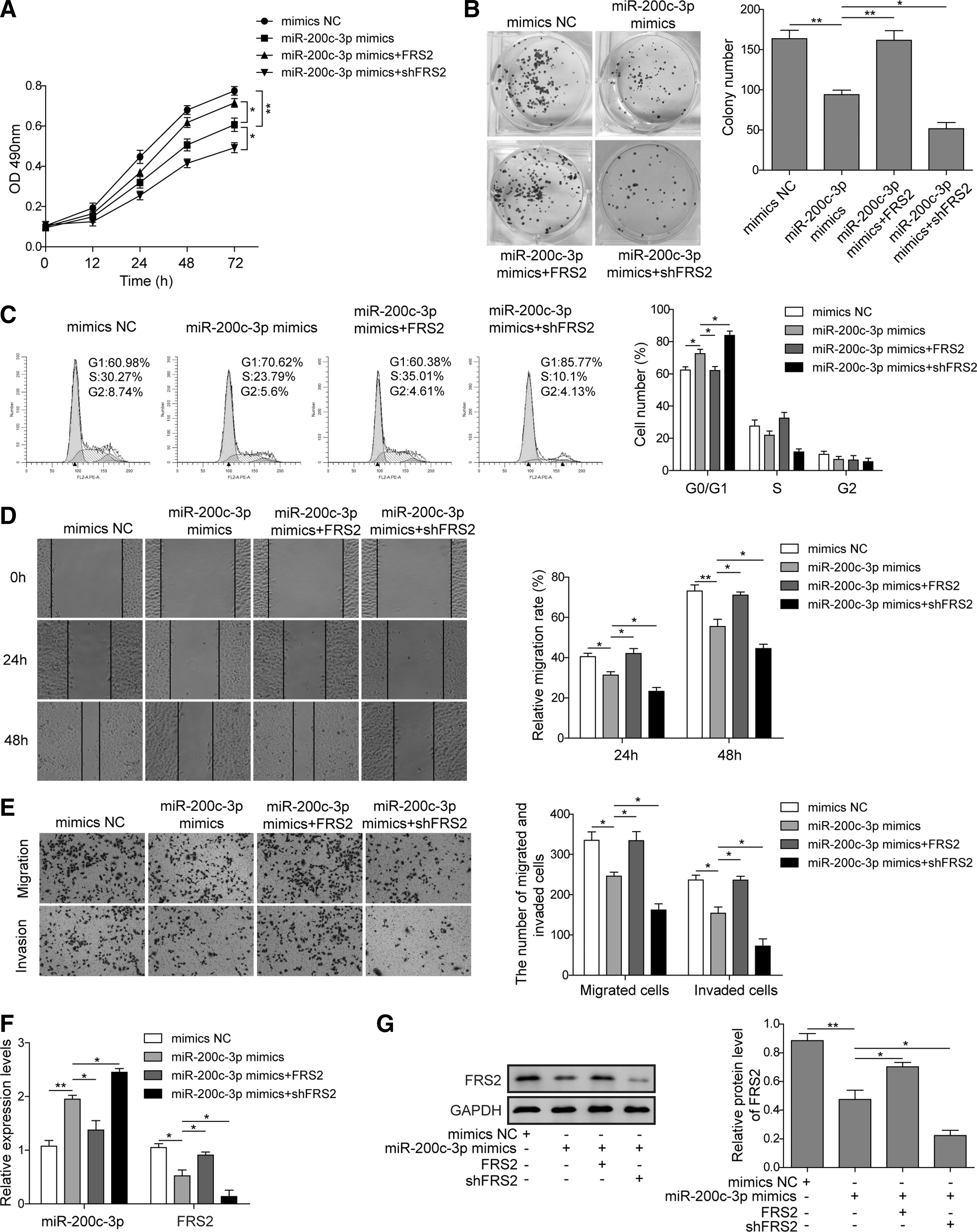
miR-200c-3p inhibits proliferation, invasion, and migration in SK-NEP-1 cells by targeting FRS2. SK-NEP-1 cells were transfected with miR-200c-3p mimics or FRS2-overexpressing plasmid or FRS2-silencing plasmid for 48 hours, either alone or in combination as indicated. The proliferation of SK-NEP-1 cells was examined by CCK-8 assay
Discussion
Although WT has an approximately 90% overall survival rate, a proportion of patients with unfavorable histology display a poor prognosis.20,21 Furthermore, because WT frequently occurs at preschool age,22,23 the standard treatment protocol involving surgical resection and chemotherapy usually causes tremendous physical and mental trauma to children. In addition, the fact that WT usually does not manifest obvious early symptoms poses a great obstacle to the diagnosis of WT.24,25 Therefore, exploring the molecular elements that participate in the regulation of WT progression is of potential significance for the clinical diagnosis or targeted intervention of WT.
miRNAs are reported to function as important regulators in the pathology of WT. In WT patients, Jiang et al. have observed a significant upregulation of miR-1180, which can decrease the expression of a tumor suppressor p73 and inhibit the apoptosis of nephroblastoma cells. 10 Moreover, inhibition of miR-1180 in nude mice restrains the growth of WT xenograft tumors in vivo, suggesting the clinical potential of miRNAs for the targeted intervention of WT. Our work expanded the knowledge toward anti-WT miRNAs by showing that miR-200c-3p reduced multiple malignant behaviors of nephroblastoma cells including cell proliferation, migration, and invasion. The effects of miR-200c-3p are dependent on its ability in downregulating the expression of FRS2.
As a tyrosine kinase adaptor protein, the oncogenic roles of FRS2 have been found in many human cancers.26,27 For example, FRS2 was abnormally amplified in high-grade serous ovarian cancer, leading to the hyperactivation of downstream Ras–MAPK-ERK signaling pathway that facilitated the oncogenic transformation of cancer cells. 17 Amplification of FRS2 was also found in liposarcoma, silencing FRS2 expression prevented the overactivation of FGFR/FRS2 signaling and inhibited the proliferation of FU-DDLS-1 liposarcoma cell line. 28 In non-small cell lung cancer, increased expression of FRS2 promoted the activation of ERK1/2 signaling pathway, thereby enhancing the proliferative and migratory ability of A549 lung cancer cells. 29 Because of the oncogenic properties of FRS2, miRNAs that target FRS2 can be putatively considered as tumor suppressive. Indeed, it is reported that miR-4653-3p could target FRS2 for downregulation in breast cancer cells, through which miR-4653-3p sensitized tumor cells to tamoxifen-induced cell apoptosis. 30 In addition, miR-206-mediated downregulation of FRS2 suppressed the growth and metastasis of gastric cancer cells. 31 In WT, miR-613 targeted FRS2 and decreased the proliferation, invasion, and migration of WT cells. 18 Our study identified miR-200c-3p as a new FRS2-targeting miRNA, and thus provided new insight into the miRNA-mediated regulatory network of nephroblastoma.
In summary, the study revealed the downregulation of miR-200c-3p in nephroblastoma, which played a pivotal role in suppressing the proliferation, invasion, and migration of nephroblastoma cells. Meanwhile, further molecular mechanisms point to the conclusion that miR-200c-3p directly targeted FRS2 and inhibited the development of nephroblastoma.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.