
Abstract
Dinoflagellates of the genus Symbiodinium form symbiotic relationships with corals, other marine invertebrates, and protists; thus, they are considered as important species in coral reef ecosystems. If Symbiodinium could be successfully cryopreserved, the cell bank generated could prove to be a valuable resource for researchers interested in basic biological research of Symbiodinium–invertebrate symbioses. Herein, successful cryopreservation of clade D Symbiodinium was achieved using a two-step freezing protocol. Symbiodinium cells were exposed to cryoprotectants (CPAs) for 30 minutes before being vapor frozen for 20 minutes in liquid nitrogen (LN2); afterward, cells were immediately immersed in LN2 for 2 hours or 10 days. The initial experiment was conducted with the following CPAs at 1, 2, and 3 M concentrations: methanol, dimethyl sulfoxide, glycerol, ethylene glycol (EG), and propylene glycol (PG). It was found that infiltration with 2 M EG and PG yielded cells with the highest percentage viability. Upon thawing, culture of these Symbiodinium was carried out for 2 months in a growth chamber, and cells continued to grow and proliferate over this period. This represents successful cryopreservation of a dominant reef coral symbiont, a feat that will ideally aid in future research of this important lineage of dinoflagellate.
Introduction
The populations of many coral species are rapidly declining after numerous unprecedented mass coral bleaching and degradation events, which are caused by major environmental changes and various anthropogenic activities such as fishing, coastal development, and shipping.1–3 The reef crisis not only affects the local community in terms of food security and social economy, but it also has a profound impact on the entire marine food chain. 4 Although coral reefs are capable of self-recovery, the process requires several years under pristine conditions with controlled management. 1 This has led to various scientific studies on the ex situ culture of corals to ensure that coral species are maintained and preserved for future propagation in their natural environment.5,6 The increasing number of coral species that require conservation, and the associated resource and space requirements needed for coral cultivation, present a substantial challenge.
To coordinate efforts for ex situ coral conservation, coral fragments, gametes, and larvae have been cryopreserved and stored in coral cryobanks for future propagation and research. In addition to coral cells, corals' symbiotic dinoflagellates, Symbiodinium, have been cryopreserved. 7 This is critical because most reef-building coral species depend on their Symbiodinium for nutrients and calcification, which result in a faster growth rate, better gamete quality, and robustness.7–9 Symbiodinium are of particular interest because they are thermosensitive and may leave, or be expelled from, their coral hosts when temperatures increase; given the reliance of corals on photosynthate translocated from Symbiodinium, this “bleaching” phenomenon can lead to coral death and subsequent reef degradation.10,11 To preserve certain clades of Symbiodinium from extinction, it may be necessary to consider cryopreservation.
Despite playing a prominent role in coral physiology and conservation, only a handful of cryopreservation studies, with varying levels of post-thaw viability, have been conducted on Symbiodinium (e.g., Symbiodinium isolated from Pseudopterogorgia elisabethae, 12 Juncella fragilis, 13 and Symbiodinium from three species of coral, including Fungia scutaria, Porites compressa, and Pocillopora damicornis 14 ). These foundational studies have provided useful information for algae cryopreservation, and have highlighted that with the optimum use of cryoprotectants (CPAs) and suitable freezing and culture protocols, cryopreservation of Symbiodinium cells is possible. With this information in hand, the direction for future investigation has many options, but first establishing which genus of Symbiodinium merits the greatest attention for cryopreservation is of interest. Although it might be advantageous to attempt to cryopreserve thermosensitive Symbiodinium strains, such as many of those from clade C, we sought herein to focus on one lineage known to be particularly resilient to elevated temperatures: clade D. The primary reason for this choice is that as corals continue to undergo catastrophic bleaching on an annual basis, there could come a time when it is pertinent to reseed reefs with corals possessing thermotolerant Symbiodinium. Therefore, we hypothesized herein that we could develop a protocol for successful cryopreservation of clade D Symbiodinium.
Methods
Symbiodinium culture and maintenance
clade D Symbiodinium (CCMP2556) cultures were purchased from the Provasoli-Guillard National Center for the Culture of Marine Phytoplankton at Bigelow Laboratory for Ocean Sciences, East Boothbay, Maine. Before the cryopreservation experiment, Symbiodinium cells were cultured in f/2 medium (Sigma-Aldrich, St. Louis, MO) at 25°C ± 1°C with light (100 μmol·m−2·s−1), under a 12-hour light:dark cycle at a density of 105 cells·mL−1. Culture medium was replenished every 10 days with fresh f/2 medium containing antibiotics (10 μg·mL−1 streptomycin and 10 U·mL−1 penicillin; Invitrogen, Carlsbad, CA).
Identification of Symbiodinium clades
DNA from Symbiodinium cells were extracted using ZR Plant/Seed DNA MiniPrep™ (Zymo Research, Irvine, CA), according to the manufacturer's protocol. The 23sDNA product was amplified with 10 × AccuPrime polymerase chain reaction (PCR) buffer, 200 nM of each primer, and 5 U AccuPrime Pfx (Invitrogen) in a 50 μL reaction mix. The forward and reverse primers used for the 23rDNA amplification were as follows: 5′CACGACGTTGTAAAACGACGGCTGTAACTATAACGGTCC3′ and 5′GGATAACAATTTCACACAGGCCATCGTATTGAACCCAGC3′, respectively. 15 Conditions for the PCR amplification were as follows: initial denaturation was performed at 94°C for 5 minutes, followed by 30 cycles at 94°C for 30 seconds, annealing at 50°C for 30 seconds, and extension at 72°C for 60 seconds. The obtained PCR products were resolved through gel electrophoresis on a 1% agarose gel stained in ethidium bromide. The resolved PCR products were further purified using the Axygen DNA Gel Extraction Kit (Axygen Biosciences, Union, CA) and cloned using the PCR-Blunt II TOPO Cloning Kit (Invitrogen). Four clones were sequenced using ABI 3100 (Applied Biosystems, Grand Island, NY).
Separation of Symbiodinium
Before cryopreservation, fresh Symbiodinium cells were cultured in a 160 mL cell-culture T-flask at 25°C in a 12-hour light:dark cycle growth chamber. To collect the Symbiodinium cells, the culture flask was shaken vigorously to detach the Symbiodinium cells that had adhered to the container wall. Aliquots were then poured into 50-mL centrifuge tubes and centrifuged at 2000 rpm for 3 minutes. The culture medium supernatant was discarded, and filtered seawater (FSW; salinity: 35 ppt) was added to resuspend the algae pellet. FSW used for this experiment was prepared by filtering natural seawater through 0.2 μM membrane filters (ADVANTEC, Japan) by using a vacuum pump (Rocker 300; Rocker Scientific, Taiwan). The aliquot was recentrifuged at 2400 rpm for 3 minutes to wash the algae. The supernatant was discarded, and FSW was added to resuspend the algae pellet. During the first observation of the algae sample, the Symbiodinium cells had aggregated into a large mass. Thus, a simple separation, which was performed using a mechanical method with a syringe needle (23 G × 1/2′′ syringe with a 0.45 × 13 mm needle; TERUMO, Philippines), was tested. When the large aggregated cells were drawn through a syringe needle with a small diameter, the cells separated due to the suction force. This motion was repeated until all the aggregated cells were separated. The draw/release motion was tested 3, 5, 8, and 10 times to determine the number of sequences needed to effectively separate the Symbiodinium cells. The observation of aggregation percentage of Symbiodinium according to the number of passes through the needle was conducted using a light microscope (C31, magnification: 400 × ; Olympus, Japan) with a camera system and a software (Micrometrics SE3, Taiwan). Aggregation percentage was calculated by the following equation: aggregation percentage (%) = aggregated area/total area × 100.
Symbiodinium density
After the initial Symbiodinium collection, the density of the Symbiodinium cells was ascertained using a hemocytometer. First, 10 μL of the algae aliquot was pipetted onto a Neubauer counting chamber and covered with a cover glass. The density of the algae was then adjusted through dilution with FSW to obtain ∼1.5 × 106 cells/mL for algae cryopreservation. 11
Symbiodinium cryopreservation
Symbiodinium cells were equilibrated with methanol (MeOH), dimethyl sulfoxide (DMSO), glycerol (Gly), ethylene glycol (EG), and propylene glycol (PG) CPAs at 25°C to the final concentrations of 1, 2, and 3 M for 30 minutes. The conditions chosen were mainly based on a previous publication. 11 All the CPAs were prepared using FSW. EG, PG, and Gly were purchased from JT Baker (Phillipsburg, NJ), DMSO from Sigma (St. Louis, MO), and MeOH was acquired from Merck (Darmstadt, Germany). During the equilibration period, Symbiodinium cells were drawn into a 250 μL cryostraw. After equilibration, the cryostraws were placed on top of a cooling device (Taiwan patent No.: M394447) 11 at a distance of 5 cm from liquid nitrogen (LN2) and cooled with the LN2 for 20 minutes. The straws (0.25 mL) were then immediately plunged into LN2, and stored for 2 hours in a polystyrene box and 10 days in a dewar. Thawing was conducted by retrieving the straws and introducing them into a warm water bath at 37°C for 10 seconds. The straws were then cut and gently agitated to discharge the aliquot.
Adenosine triphosphate viability assay
The viability of the Symbiodinium was validated using a cell viability assay kit (ApoSENSOR ATP Cell Viability Bioluminescence Assay Kit) purchased from BioVision (Milpitas, CA). The procedures were conducted according to the methods described by Chong et al. 13 In brief, 10 μL of the post-thaw algae was incubated with 100 μL of the nucleotide-releasing buffer for 3 minutes in a clear plastic test tube. Adenosine triphosphate (ATP) monitoring enzyme (5 μL) was then added to the algae–buffer composition, incubated for 1 minute, and assessed using a luminometer (Lumat 9507; Berthold Technologies, Germany).
Symbiodinium culture
The culture medium was prepared by mixing 32 g of Coralife Salt Water Mix (WI) with 1 L of distilled water to create 32 ppt artificial salt water (ASW). ASW was then autoclaved, and 970 mL of ASW was added to 30 mL of Guillard's f/2 medium (Sigma-Aldrich) and 1 mL of antibiotic solution (penicillin: 100 U/mL; streptomycin: 10 mg/mL). The solution was then filtered through a 0.45-μM filter (Nalgene Rapid Flow; Thermo Scientific). After the ATP bioassay, the post-thaw samples were centrifuged (800 rpm for 3 minutes at 25°C) to remove the CPA. The resuspended algae solution was then added to a 20 mL cell-culture T-flask with a final volume of 20 mL of culture medium, and this flask was placed in a growth chamber under a 12-hour light:dark cycle. The culture medium was changed every 7 days. After the medium exchange, the flask was shaken vigorously to resuspend the Symbiodinium. Aliquots of 10 μL of the culture were retrieved to determine the cell density through the hemocytometer method.
Negative control
To estimate the contribution of inadvertently cocultured bacteria to ATP content, we allowed Symbiodinium to settle in the tubes (the cells are relatively dense), and then analyzed the ATP concentration in the supernatant. We routinely measured ATP levels of <50 relative units of light (RUL), ∼200-fold less than values obtained with dinoflagellate cells. We therefore estimate the bacterial contribution to the ATP contribution is ∼0.5%, and this contribution did not vary between control and cryopreservation treatments (p < 0.05).
Statistics
Symbiodinium from each treatment were used to obtain three biological replicates, each of which was analyzed three times. The one-sample Kolmogorov–Smirnov test was performed to determine if the data were normally distributed. Statistical data were then analyzed using one-way analysis of variance with Tukey's post hoc tests (SPSS, version 17.0) to test for significant differences between the mean values from experimental treatments and to identify specific differences between groups. The results of the study are presented as mean ± standard error.
Results
Cooling rate
Samples were positioned 5 cm from the LN2 and cooled at a rate of −60°C per minute, until reaching a final temperature of −123.9°C, and then were plunged into LN2.
Mechanical method for separating Symbiodinium cells
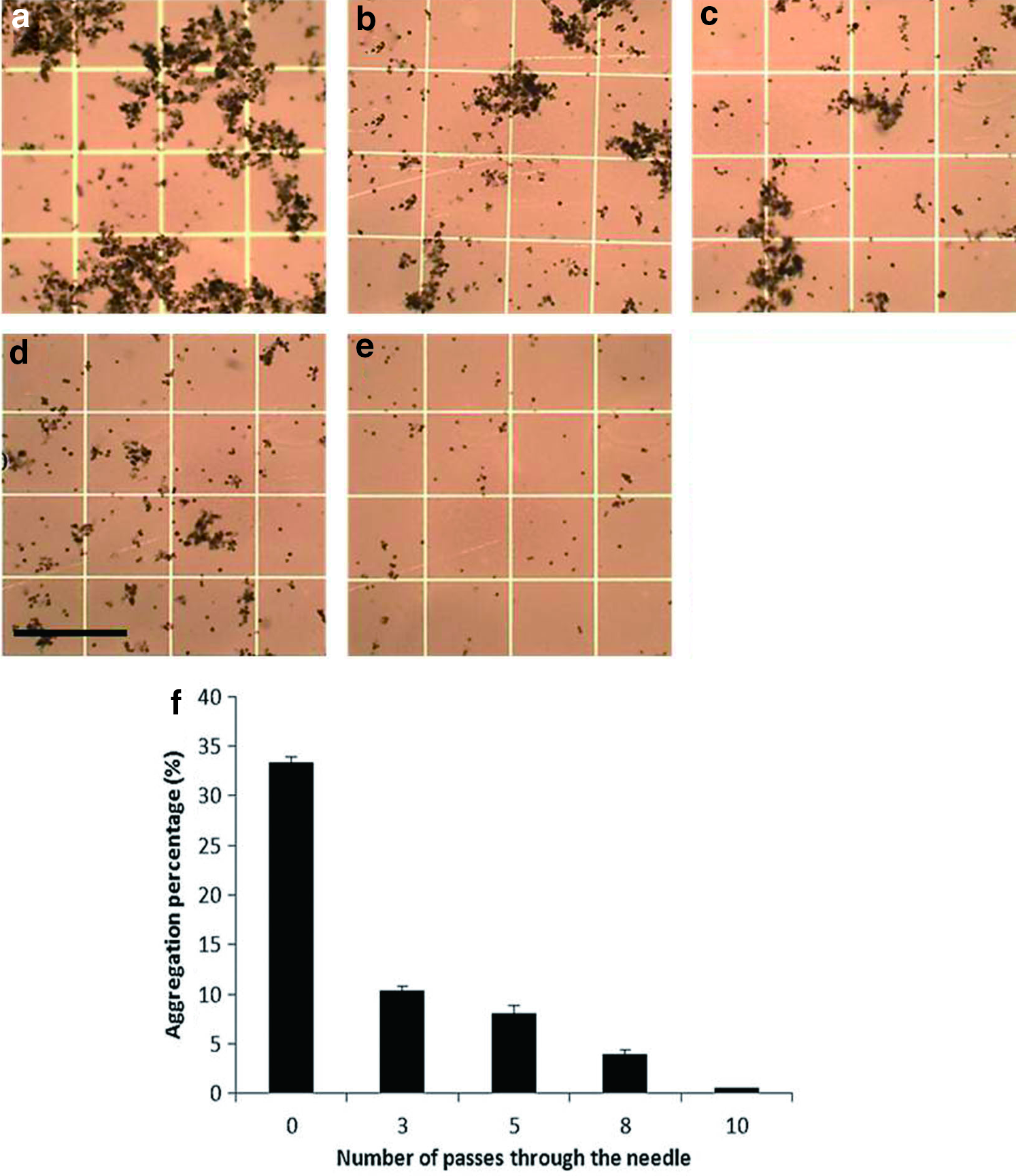
Symbiodinium cells exhibit aggregation behavior. Due to this characteristic, the clumped outer coat of the cells may affect CPA penetration, post-thaw procedures, and ATP results during cryopreservation. Four different mechanical separation treatments (3, 5, 8, and 10 draw/release times through a syringe needle) were tested in our experiments. The aggregated condition before the mechanical treatment of the cells (i.e., the control) can be seen in Figure 1a. In Figure 1b–e, the cell separation caused by repeated pipetting using a needle is visible; separation increased with draw/release frequency. The aggregation percentages of Symbiodinium cells in the control and after each treatment are shown in Figure 1f. Aggregation decreased from 33% to 0.5% based on the number of passes through the needle. The best cell aggregation percentage (0.5%) was obtained after 10 needle passes. The normalized percentages of separated Symbiodinium cells in the control and after each treatment are shown in Supplementary Figure S1. The control shows a 0% separation of algae cells; however, after ≥3 passes through the needle, the separation percentage slightly increases. The separation percentage of cells reached 98% after 10 passes through the needle. The following experiments on Symbiodinium were possible because of the successful separation of the clumped cells.
Cryopreservation
During the 2-hour transition within LN2, various CPAs with different concentrations (1, 2, and 3 M) were added to protect the Symbiodinium cells (Fig. 2a). The most suitable CPA was determined to be EG because each concentration tested achieved an ATP concentration >100%. PG was deemed to be the second best CPA, but only at a concentration of 1 M. MeOH was the least suitable CPA, and DMSO and Gly were also found to be poor CPAs because their ATP concentrations were consistently lower than that of the control.
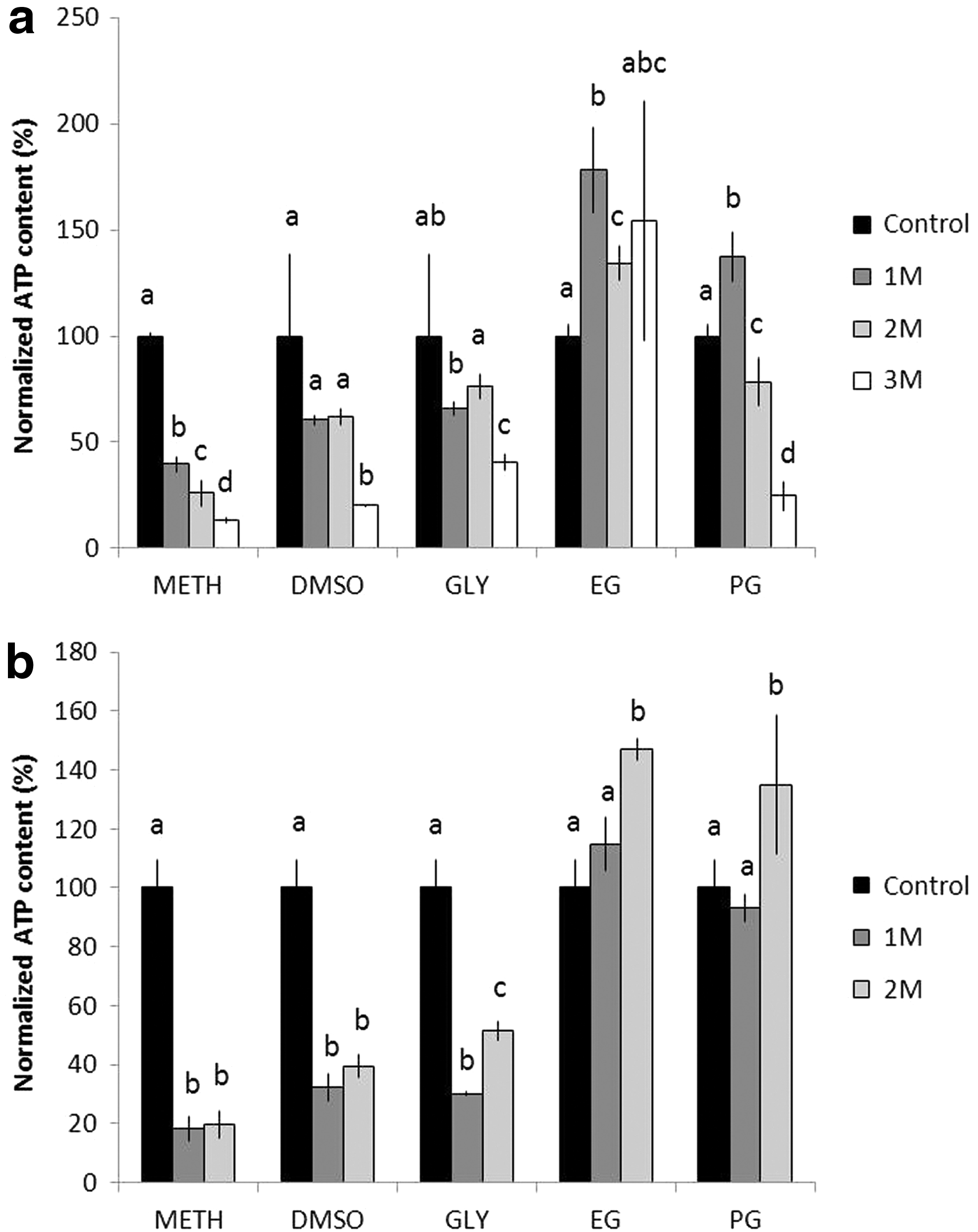
Effect of the various CPAs on the intracellular ATP concentrations after freezing through
The effectiveness of the various CPAs was further assessed using an ATP bioassay post 10 days of dinoflagellate freezing (Fig. 2b). Figure 2b illustrates that both the 2 M EG (150%) and 2 M PG (135%) exhibited higher ATP concentrations than their 1 M counterparts. Accordingly, EG and PG were still considered the optimal CPAs, DMSO and Gly remained poor, and MeOH was the least suitable for cryopreservation due to its extremely low ATP concentration compared with that of the control. After identifying the top CPA concentrations, only the effective 1 M and 2 M CPA values were compared; Figure 3 compares the ATP concentrations after 2-hour and 10-day freezing periods in LN2. In general, the viability of the cells after 10 days was significantly lower (p < 0.05) than that found after 2 hours of freezing, with the exception of the 2 M EG and 2 M PG where the cell viability was higher after 10 days of freezing.

Cryopreservation of the Symbiodinium by using various CPAs at various concentrations, which were compared between 2 hours and 10 days of freezing in LN2. The experiment was conducted with concentrations of 1 and 2 M for each CPA. Different letters represent significant difference between CPA concentrations and freezing periods for Symbiodinium treated with the same CPA (p < 0.05).
Culture of Symbiodinium cells
Symbiodinium cells cryopreserved with 1 and 2 M of EG and PG were thawed and cultured to assess their long-term cell viability. Symbiodinium cultures, obtained from both EG and PG as well as Symbiodinium (noncryopreserved), revealed similar trends in both ATP concentration and cell density during the 56-day culture period (Figs. 4 and 5). ATP concentrations of four frozen-thawed culture groups decreased significantly during the initial 14 days of culture (p < 0.05); however, as the culture period progressed, the ATP concentrations of all the culture groups recovered significantly. After 56 days of culture, the Symbiodinium cells in 1-M EG reached the same ATP concentration as the control (day 0) (Fig. 4a). The ATP concentrations of Symbiodinium cryopreserved in 2 M EG, 1 M PG, and 2 M PG reached 1.5, 0.8, and ∼1.2 RUL, respectively (Fig. 4a, b). Symbiodinium (noncryopreserved) obtained consistent ATP concentrations throughout the 56-day culture period (Fig. 5). Interestingly, although ATP concentration of Symbiodinium cryopreserved in both EG and PG decreased during the initial culture period, cell density steadily increased throughout the 56-day culture period (Fig. 4). At day 0 of culture, all Symbiodinium cell densities were <100,000 cells/mL, and after 56 days of culture, all cell densities apparently increased to ∼200,000 cells/mL (Figs. 4 and 5). Although no significant differences in the viability of cultured Symbiodinium cells were observed between 1 and 2 M of EG and PG (Fig. 4a, b), the 2 M concentrations of both CPAs appeared to perform better than the 1 M concentrations.

Cell density (lines and left y-axis) and normalized ATP concentrations (columns and right y-axis; an indicator of viability) of the post-thaw Symbiodinium cells, cryopreserved in 1 M and 2 M
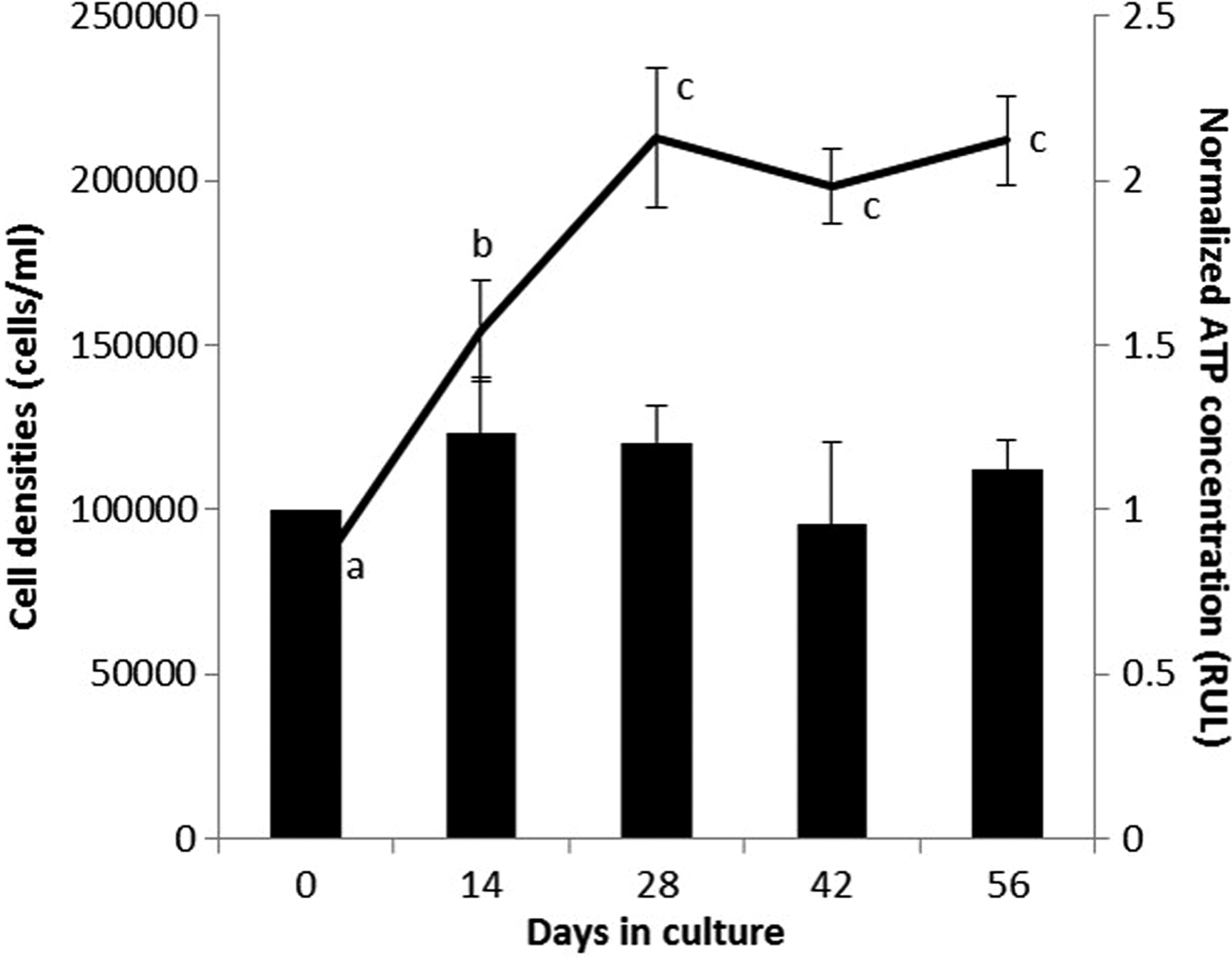
Cell density (lines and left y-axis) and normalized ATP concentrations (columns and right y-axis; an indicator of viability) of the Symbiodinium cells (noncryopreserved) cultured for 56 days. The letters over the lines and columns indicate the differences (p < 0.05) over time for the cell density and ATP concentration data, respectively.
Discussion
clade D Symbiodinium is commonly found in corals situated in near-shore areas with high turbidity.16–19 In this study, we cultured clade D Symbiodinium as an alternative to Symbiodinium isolated from wild corals. Before the cryopreservation procedure, Symbiodinium cells were passed through a syringe needle to ensure that the aggregated cells were well separated into single cells, similar to the method described by Chong et al. 13 This was performed because cell aggregation reduces the cell surface area for CPA permeation, thereby decreasing the cell dehydration efficiency and causing intracellular ice crystallization.20,21 Mechanical dissociation of the aggregated Symbiodinium cells by using a 23-gauge syringe needle was effective, and passing the sample through the syringe needle 10 times was optimal.
The effectiveness of each CPA is highly species specific, with each CPA exhibiting varying effectiveness at different concentrations for each species and cell.22,23 This species specificity was also demonstrated in our present experiment. We tested the five most frequently used CPAs for Symbiodinium, namely MeOH, DMSO, Gly, EG, and PG, at three concentrations.13,24 Our results indicated that more than one CPA could be utilized for Symbiodinium cryopreservation. Both 1 and 2 M of EG and PG provided excellent cryoprotection. This was an interesting finding because, in previous cryopreservation of clade G Symbiodinium, neither EG nor PG exhibited satisfactory post-thaw viability (30% of the normalized ATP concentration). 13 Moreover, the application of EG and PG in marine microalgae studies is slightly limited compared with their use in other types of microorganisms. 25 This may be attributed to EG being harmful for cells with high polysaccharide content because they may react and dissolve the cell wall and membrane. 26 Nonetheless, in other studies of marine algae, EG was effective only when combined with other CPAs, possibly to reduce its toxicity. In previous successful algae experiments with EG, the combination of DMSO, EG, and proline was used for the cryopreservation of Nannochloropsis oculata and Tetraselmis tetrathele, 27 and EG was combined with trehalose for the cryopreservation of Isochrysis galbana and three strains of Attheya sp. 28 However, this does not explain our result because EG used alone also maintained high Symbiodinium viability. Similarly, PG has previously shown incongruity in its cryoprotection ability for various microorganisms. However, regarding the cryopreservation of algae, PG was effective for the cryopreservation of Porphyra yezoensis when combined with DMSO. 29 Therefore, we deduced that the cryoprotection efficiencies of EG and PG are highly species dependent, 25 and may be affected by the cryopreserved sample; however, the exact mechanism remains unidentified.
Gly, MeOH, and DMSO are three of the most commonly used CPAs for marine algae. 30 However, in this experiment, only Gly maintained adequate cryoprotection for clade D Symbiodinium Previous studies have used Gly as the CPA for N. oculata, 31 Tetraselmis suecica, 32 Tetraselmis striata, 33 and many more. 30 Although the findings of these studies and our present experiments have demonstrated that Gly is effective, it lacks penetrability compared with MeOH and DMSO. 34 Therefore, a longer equilibration was needed for effective freezing with Gly. Our 30-min equilibration period was similar to that described by Chong et al., 13 but the clade G Symbiodinium recorded 30% of ATP viability with Gly, whereas clade D Symbiodinium exhibited a >50% post-thaw viability. Therefore, in addition to the permeation properties of Gly, the different permeabilities of the algae membrane structure may be a deciding factor in CPA cryoprotection. Notably, both MeOH and DMSO exhibited very little cryoprotection for the Symbiodinium sp. in the present experiment. The viability of DMSO and MeOH was not effective for clade D. This was contrary to previous studies on Symbiodinium sp. cryopreservation, because Chong et al. 13 reported high viability when clade G was cryopreserved with MeOH or DMSO. Moreover, Symbiodinium clade B isolated from P. elisabethae was successfully cryopreserved with MeOH or with ethanol, 12 and Symbiodinium sp. isolated from F. scutaria, P. damicornis, and P. compressa were successfully cryopreserved with a vitrification solution comprising MeOH and DMSO. 14 We inferred that although the longer equilibration period of 30 minutes was sufficient for Gly to permeate into the Symbiodinium for the freezing, it was unsuitable for MeOH and DMSO. Owing to their high permeability, MeOH and DMSO were efficient in cell dehydration but may have had a cytotoxic effect on different species of algae when used at high concentrations. 35
Although the cryoprotection ability of each CPA evidently varies according to the species and cell, the CPA concentration is equally important in deciding its effectiveness. This is because when used in unsuitably high concentrations, the chemicals may present a cytotoxic effect on the cell.36–38 The tolerance to different CPA concentrations varies among cells, but a greater tolerance to higher concentrations of CPA is generally preferable for faster permeability. Our present study of clade D Symbiodinium indicated that for all the CPAs tested (MeOH, EG, PG, Gly, and DMSO), 2 M was the upper limit, and 3 M resulted in the negative outcome. A concentration of 1 M of the CPAs was generally preferred for clade D; however, no significant difference was noted when 2 M CPA concentrations were used. A low CPA concentration was more effective and less harmful to Symbiodinium clade G; incubation with 1-M MeOH and 2-M DMSO resulted in higher viability than did incubation with 3-M MeOH and 3-M DMSO. 13 Nonetheless, CPA toxicity tolerance varied from cell to cell. Although Symbiodinium clades D and G preferred lower concentration of CPAs, clade B showed high viability even when cryopreservation was performed with a CPA mixture of 20% MeOH (6 M) or 20% ethanol (4 M). 12 Some algae may necessitate higher concentrations, which could be harmful. The manipulation of other conditions, such as decreasing the equilibration period and temperature, may reduce the cytotoxicity and osmotic stress of the CPA. This manipulation of time and temperature was utilized for clade B Symbiodinium cryopreservation, in which the incubation period was only 10 minutes and the temperature was lowered to 4°C. 12
Two-step freezing is the most effective freezing procedure for the cryopreservation of microalgae.12,30 Although the general procedure involves cooling a sample to a certain temperature and then immersing it in LN2 for storage, each step of the freezing method varies for different species of algae. This is because each species' tolerance to extreme low temperatures innately differs; therefore, the holding time, cooling rate, and cooling temperature should be optimized to provide the optimal post-thaw viability. 39 In the present experiment, we utilized a simple variation of the two-step freezing method by suspending the samples atop a freezing device similar to that described by Chong et al. 13 This variation of the freezing method is popular among cryopreservation studies of microorganisms because of its ease of setup and operation, mobility, and low cost. The distance of the sample from the LN2 was manipulated to achieve different cooling rates. Our sample was placed at a distance of 5 cm above the LN2 for 20 minutes. The cooling rate utilized was considered as uncontrolled cooling, and it exhibited a nonlinear trend. The initial cooling rate was rapid, and it then gradually decreased to the final temperature of −123.9°C. Although previous microalgae studies have reported high viability when samples are cooled to higher temperatures between −20°C and −70°C, 37 the present final temperature of −123.9°C appeared effective when used with EG and PG. Moreover, faster and lower cooling temperatures are more effective in preventing internal freezing due to slower chemical reaction rates. 40 Lower temperature is also optimal for long-term storage.41,42
The main aim of cryopreservation is to preserve samples for a prolonged period. Theoretically, samples that are stored in LN2 have a shelf life of 1000 years. 43 However, some microalgae experience degradation and decreased post-thaw viability after being cryopreserved for 1 year. Nevertheless, the viability was maintained up to two decades for some algae species through an optimum cryopreservation protocol. 44 Lower storage temperatures correlate with high viability during preservation. Storage in LN2 or at a temperature <−135°C is suggested for long-term preservation without deterioration. 41 Hence, to ensure that our present cryopreservation protocol was optimal, our samples were maintained in LN2 for a short-term storage of 10 days. Although cells often exhibit lowered viability after a long storage period, two of the CPA treatments revealed notable results. Clade D samples cryopreserved with 2 M concentrations of both EG and PG demonstrated an increase in ATP concentration even after 10 days of storage in LN2. Although the actual mechanisms for this improved viability are unknown, it is possible that an insult from cryopreservation might be compensated by cells' biochemical defense mechanisms.
Although viability assays were not directly involved in the cryopreservation protocol, the purpose was to determine its efficiency. Thus, it was imperative that the viability criteria emulate the actual physiology of the cryopreserved samples. Most cryopreservation studies consider reproductive ability as an indication of robustness or high viability, which is why most algae cryopreservation studies assessed algae viability through culturing the post-thaw sample.12,44,45 Indeed, the ability to multiply during culturing may indicate that the metabolic activity of the cells was optimum for mitosis or reproduction. However, a long culture period is required to ascertain the viability of the algae culture, and culture conditions such as light intensity and light regime would also require optimization to reduce the detrimental effects in the weakened post-thaw algae cells. 45 This would necessitate trials in various conditions to determine the optimal culture conditions for the post-thaw algae. Finally, the growth of the culture would also be determined through assessment methods such as fluorescent staining for fluorescent cytometry detection of the colony size12,46 or cellular metabolic activity, for example, photosynthetic efficiency14,45 and ATP levels. 13 In the present experiment, viability was also determined through cellular ATP concentration because this was regarded as the most vital molecule for normal cellular metabolic activity. Cellular ATP rapidly reacts with the luciferase enzyme to emit a fluorescent signal, which is then quantified with a luminometer.47,48 A major difference between the ATP bioassay and other cellular staining methods was that the ATP bioassay did not provide the actual number of dead algae but instead quantified the ATP in the sample. This flexibility allowed the user to determine the threshold value of a viable cell, wherein the higher ATP concentration of a treatment would indicate more viable algae and vice versa. The small sample volume needed for the assessment also guaranteed that the remaining cryopreserved sample could be used for algae culture.
Thawed Symbiodinium cryopreserved in EG and PG were selected for culture because their post-thaw results were promising. Indeed, Symbiodinium cryopreserved in both the CPAs showed a significant increase in ATP viability and cell density during the 2-month culture period. Notably, although ATP concentration during the culture period was significantly lower than that of the control in the initial culture period, the cell density exhibited the opposite relationship. We deduced that the difference in the viability assessment was caused by the overestimation of cell density. Dead and dying cells do not contain or produce ATP, and low ATP concentration correlates with cell injury and cell death. 49 Low viability, as indicated by the low ATP concentration, was highly possible because the Symbiodinium cells may have sustained various stresses and freezing injuries from the cryopreservation, thawing, and effect of CPA toxicity. By contrast, assessment disparity was common in cell counting because live and dead cells could not be distinguished without viability. Regardless of the conditions, ATP concentration may be a better assessment measure of cell survivability along with cell density assessment because ATP is essential for all cellular activities and growth. In the subsequent sessions of viability assessment, both ATP and cell density were observed to increase concurrently, indicating that Symbiodinium was indeed proliferating.
The present post-thaw culture results differed significantly from the culture results of Chong et al. 13 and Santiago-Vázquez et al. 12 The lipid levels, nitrogenous compounds, and cell wall components of cultured and endosymbiotic Symbiodinium sp. were significantly different.48,50 Lipid plays important role in cell cryotolerance, and endosymbiotic algae may have conferred tolerance to the cryopreservation because oocytes with a higher polyunsaturated lipid content are more tolerant to cooling temperatures.51,52 Moreover, the contrasting surroundings greatly affect the morphology. Endosymbiotic algae remained in a coccoid stage and may have been enveloped by a host tissue. 53 The presence of the tissue layer may have decreased the efficiency of some CPAs, whereas those with high permeability, such as MeOH or DMSO, may be a more suitable choice. The robustness of the cryopreserved Symbiodinium culture in this study could be attributed to the cultured origin of the Symbiodinium samples. Our experiment was conducted on cultured Symbiodinium sp., whereas Chong et al. 13 and Santiago-Vázquez et al. 12 worked on coral-isolated endosymbiodinium. Cultured Symbiodinium easily adapt to artificial environments and hence recover more rapidly after cryopreservation. Endosymbiodinium cells would require a longer period to acclimatize to a free-living state, and culture conditions for endosymbiodinium cells have not matured. Thus, in addition to the species differences between our and previous studies, the divergence in cellular physiology from vastly different habitats could affect cryopreservation.
In conclusion, Symbiodinium sp. (C, D, and G) exhibited clade specificity in their cryopreservation protocols. In addition, the different cryopreservation requirements of clade D Symbiodinium compared with those in previous Symbiodinium sp. studies may be attributed to the different origins of the endosymbiotic counterparts, wherein the host–symbiont relationship may have changed the morphology and physiology. Nonetheless, in this study, we identified the two-step freezing with either EG or PG at 1 or 2 M as the optimum cryopreservation protocol for clade D Symbiodinium.
Authors' Contributions
C.L. and S.T. conceived the experiment; P.T., C.J., F.-W.K, and L.-H.W. conducted the experiment; P.-J.M. and S.T. analyzed the results; and C.J. and C.L. wrote the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.