
Abstract
The aim of the article is to report a review on different sperm cryopreservation techniques, various stress-related freeze–thaw damages altering sperm structure and function during conventional cryopreservation, and strategies to minimize these stresses. Sperm cryopreservation has allowed indefinite storage and successful transportation of valuable germplasm from proven sites at distant locations, for genetic upgradation through implementation of reproductive techniques, such as artificial insemination. Different techniques for sperm cryopreservation have been proposed such as conventional freezing techniques, directional freezing, and sperm vitrification. Drawbacks related to conventional freezing methods, such as heterogeneous ice nucleation and repeated freeze–thaw cycles at the ice front that disrupts and kill sperm cells, led to the emergence of the directional freezing technique. Sperm vitrification is advantageous as there is no ice crystal-induced physical damages to sperm. However, sperm vitrification has less applicability as encouraging results are only reported in human, dog, and cat. In spite of several drawbacks, conventional freezing techniques are still most widely used for sperm cryopreservation. Spermatozoa experience stresses in the form of cold shock, osmotic stress, and mainly oxidative stress during conventional cryopreservation ultimately reduces the sperm viability and fertility. Several attempts have been made in the past to minimize all these stresses individually or in combination. Membrane fluidity was increased to prevent the cold shock and cryocapacitation-like changes by the addition of cholesterol to the membrane. Antifreeze proteins were added in semen extender to minimize freeze–thaw damages due to heterogeneous ice nucleation and ice recrystallization. Oxidative stress was reduced either by neutralizing reactive oxygen species (ROS) through enzymatic, nonenzymatic, plant-based antioxidants or reductants; or by minimizing the level of sources like the semen radiation exposure, leucocytes, and dead and defective spermatozoa, which lead to ROS production during the semen cryopreservation process. A novel approach of minimizing oxidative stress was to reduce the oxygen tension in sperm microenvironment that is, extender by partial deoxygenation process, as a number of literatures pointed out direct link of O2 with ROS production. When compared with other strategies, partial deoxygenation of semen extender with N2 gassing is found as a cost-effective, comparatively easy and a potential approach to large-scale frozen semen production.
Introduction
Sperm cryopreservation has made possible the successful implementation of reproductive techniques such as artificial insemination (AI) and in vitro fertilization (IVF) worldwide. 1 AI with frozen semen became essential in breeding and selection criteria for enhancing the production potential of domestic animal species. Conventional freezing techniques are most widely used for sperm cryopreservation in spite of advances in the form of directional freezing technique and sperm vitrification. Freezing during cryopreservation is mainly done to halt the metabolic processes of the spermatozoa, allowing indefinite storage for future use or for transportation to distant locations. 2 On the other hand, the cryopreservation process is detrimental to sperm viability and fertility. 3 Dramatic changes in temperature, osmotic or toxic stresses due to cryoprotectants and mainly the effects of reactive oxygen species (ROS) due to oxidative stress, result in semen cryopreservation leading to structural and functional damages to sperm. The present review deals with different techniques of sperm cryopreservation and describes in detail various stress-related freeze–thaw damages during conventional cryopreservation, with possible strategies to minimize these stresses. It mainly covers domestic species while other species like human and wildlife species are also mentioned wherever required, considering the diversity of the topic.
Sperm Cryopreservation Techniques
Conventional freezing technique
Freezing methods for sperm cryopreservation can be broadly characterized into three basic modes of freezing that is, Equiaxed freezing, directional freezing, and vitrification. Equiaxed (star-shaped ice crystals) freezing, is also known as programmable or nonprogrammable conventional freezing, of which most widely used is the manual freezing technique. 4 In this technique, 0.25- or 0.5-mL plastic straws filled with extended semen are placed in contact with liquid nitrogen (LN2) vapor at 4–5 cm height above LN2 for a time not greater than 15 minutes (where 10 minutes is recommended), before being plunged into the LN2 for storage. The freezing rate is ∼60°C/min. 5 In programmable freezing, different freezing/cooling rates for varied temperature ranges are programmed in a biofreezer for freezing the straws before plunging in LN2. It is species specific and varies from species to species depending on sperm quality or cryosurvivability after the freeze–thaw process. 5
To minimize cryoinjury, the cooling rate should be optimum and it should neither be high (physical damage due to intracellular ice crystallizations), nor be low (osmotic damage/architectural loss due to excessive cellular dehydration in response to extracellular ice crystallizations, “Solution effect”). The same principle is applicable for the thawing rate as low thawing rates over the critical temperature range (−60°C to −15°C) cause physical damage to sperm 6 due to ice recrystallization (smaller ice crystals agglomerate to form deformed or bigger ice crystals), and the high thawing rate causes osmotic damage to sperm as cryoprotectants are unable to leave the cell fast enough, resulting in osmotic entry of extracellular water inside the cell, cell swelling, and finally cell death. 7 Generally, thawing is done at 37°C for 30 seconds (ranging 45–60 seconds). Improper thawing procedures such as rolling of straws between hands and placing under the armpit or in pockets are often seen at the field level, which leads to sperm damage and deterioration of post-thaw semen quality. Therefore, frozen semen needs to be thawed well before insemination and needs to be inseminated as soon as it is thawed. Once straws are taken out of the container, they should not be put back into a LN2 container again. However, the overall success of sperm cryopreservation also depends on factors such as the type of freezing media and cryoprotectants, lecithin or protein source (egg yolk, soya milk, or low-density lipoproteins), and mechanical stresses involved in the processing of semen, like pipetting, dilution, mixing, packaging, storage, and transfer.
Drawbacks related to conventional freezing methods are (1) Heterogeneous ice nucleation or uncontrolled ice crystal growth in terms of both velocity and morphology that disrupts and kills sperm cells, (2) Inefficient latent heat dissipation that is, repeated freeze–thaw cycles at the ice front causing mechanical damage to cells. These drawbacks led to the emergence of a new cryopreservation technique that is, directional freezing technique. 8
Directional freezing technique
The alternative technique for sperm cryopreservation is the directional freezing technique. In this technique, a multi-thermal gradient device is used. 8 After dilution with freezing extender, semen is cooled to 4–5°C at ≈0.3°C/min and packaged in prechilled hollow tubes (5, 8, or 12 mL). Tubes are advanced through a linear temperature gradient, from 5°C (warm block) to −50°C (cold block) at a constant velocity that is, 1 mm/sec (1.0–1.5 mm/s). Thereafter, seeding is done for ∼60 seconds and through a cold block at its other end, tubes are moved to a collection chamber (−100°C). Then, these tubes are collected and transferred into LN2 for storage. For evaluation, tubes are first air thawed at ambient temperature for 90 seconds and then placed into a specialized hollow tube thawing unit (Interface Multigrad Technology, Israel) inside a water bath at 37°C for 60 seconds. Saragusty et al. 9 proposed that when freezing a large volume of semen by the directional freezing technique, a controlled ice nucleation (CIN) or seeding stage is not required. Later, double freezing technique was introduced to make use of semen of valuable sire, frozen, and stored by directional freezing technique for AI at distant locations. 10
Continuous seeding, CIN, homogeneous cooling, efficient latent heat dissipation, and higher storage economy make this technique advantageous and superior than the conventional freezing methods. 11 It allows freezing of large volumes of semen in reusable tubes, reduces mechanical damage to sperm, resulting in a high survival rate after the freeze–thaw process.11,12 Directional freezing for sperm cryopreservation has been reported in a wide range of domestic and wildlife species13–17 and also found to be superior to conventional freezing in straws.
Sperm vitrification
Vitrification is a process of transforming a solution containing high concentration of cryoprotectant in a glass-like (Latin “Vitreum”) state without ice crystal formation through an ultrarapid cooling process. 18 Three important factors, which affect the probability of success of vitrification, are cooling rate, viscosity, and volume. High cooling rate, high viscosity, and low volume that is, in cryo-loops (1 μL), droplets (30 μL), and open pulled straw (5 μL) increases the probability of a successful outcome. 12
It is a rapidly emerging technique for cryopreservation and advantageous compared with conventional cryopreservation in many aspects. It involves short equilibration times, fast cooling rates, no expensive equipment such as a programmed biofreezer, and there is no physical damage to the cells due to ice crystals. It is relatively cheaper, much faster, and simpler in application than conventional freezing methods.
Vitrification of frog spermatozoa was first reported early in 1938 by Basile J. Luyet and Eugene L. Hodapp. In spite of this, this technique had only been investigated and successfully applied to oocytes and embryos of different mammalian species.19–22 It was due to lethal osmotic effects of a high percentage of permeable cryoprotectants (30–50%) used in vitrification media on spermatozoa, which had limited its successful application. However, Isachenko et al. 23 demonstrated that vitrification of spermatozoa without permeable cryoprotectants was possible and it resulted in a higher motility after thawing as compared with conventional freezing with cryoprotectants. Conventional freezing of human spermatozoa in 0.25 mL insemination straws with permeable cryoprotectant was compared with aseptic cryoprotectant-free vitrification method (vitrification media: human tubal fluid supplemented with 1% human serum albumin and 0.25 M sucrose). For vitrification, a 0.25 mL insemination straw was filled with 100 μL of the sperm suspension, placed in a 0.5 mL insemination straw, which was hermetically closed at both ends and directly plunged into LN2. For semen evaluation, thawing was done by immersing the straw into a water bath at 42°C for 20 seconds. Both cryopreservation techniques demonstrated similar results. 24
Vitrification of human spermatozoa has been extensively studied and modified for over a decade25–27 because human spermatozoa are the smallest and shows maximum cryostability as compared with spermatozoa of animals. Spermatozoa of human > stallion > dog > cat > rabbit > ram > bull > boar have cryostability in decreasing order, and sperm of bull and boar have the lowest stability to cold shock due to their big blade-shaped flat head, therefore, less likely to sustain a vitrification process. In general, there is negative correlation between the sperm head size and vitrification success. 28 Sperm vitrification has also been reported in dog, 29 cat, 30 and ram31,32 with variable results.
In spite of several advantages of the directional freezing technique and sperm vitrification, conventional cryopreservation is the most commonly used worldwide technique for freezing of domestic animal sperm cells. Therefore, this review will now focus on various stress-related freeze–thaw damage that result in altering sperm structure and function during conventional cryopreservation, and strategies to minimize these stresses are discussed in the following section.
Temperature stress (cold shock)
Temperature stress or cold shock is related to phase change in lipids and the altered functional state of membranes33,34 as freeze fracture studies of membranes before, during, and after cooling clearly showed the phase separation events.35,36 One way to prevent membrane damage due to phase change from fluid to the gel state was to increase the membrane fluidity at low temperatures by adding cholesterol to the membrane. Different concentrations of cholesterol-loaded-cyclodextrins (CLC) have been used in cattle, 37 buffalo bulls, 38 rams, 39 stallions, 40 and boars 41 to minimize cryocapacitation-like changes and to improve cryosurvivability of spermatozoa in this regard.
Osmotic stress
Addition of cryoprotectants leads to osmotic or toxic stress, which depends upon the relative permeability of that particular cryoprotectant. 42 When a solution is cooled below the freezing point, ice crystals are nucleated and pure water crystallizes out as ice. The solutes are dissolved in the remaining liquid water fraction and the osmotic strength of the solution rises, causing a solution effect. These stresses are due to osmotic pressure change in the unfrozen fractions as a result of ice crystal formation. 43 One way to reduce freeze–thaw damages or stresses, due to heterogeneous ice nucleation and ice recrystallization, was addition of antifreeze proteins (AFPs) in semen extender to modify or inhibit ice crystal formation. These AFPs are produced by polar fish, some species of insects like beetle (Dendroides canadensis) and winter/cold-acclimated plants like winter rye (Secale cereale), carrot (Daucus carota), peach (Prunus persica), perennial ryegrass (Lolium perenne), and bittersweet nightshade (Solanum dulcamara). Beneficial effects of AFPs have been reported for sheep, cattle, and buffalo bull sperm.44–48
Oxidative stress
Oxidative stress is one of the most important factors contributing to poor post-thaw semen quality during sperm cryopreservation. 49 Oxidative stress arises due to an imbalance between the production of ROS and a biological system's ability to detoxify it that is, when oxidants exceed antioxidants. 50 It is often associated with an increased rate of cellular damage induced by oxygen (O2) and oxygen-derived oxidants. 51
ROS broadly is represented by radicals such as the hydroxyl ion, superoxide, nitric oxide, peroxyl ion, and nonradicals like peroxynitrate, ozone, nascent oxygen, and hydrogen peroxide. 52 Two ROS-generating systems are possibly involved, that is, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase at the level of sperm membrane 53 and sperm diaphorase (mitochondrial nicotinamide adenine dinucleotide–hydrogen, NADH-dependent oxidoreductase) at the level of mitochondria. 54 In bovine semen, ROS are also generated by immature, dead, and defective spermatozoa through an aromatic amino acid oxidase pathway. 55 Leucocytes and radiation exposure contribute in a minor way to the formation of ROS. 56
Inevitable fate of oxidative stress
Freeze–thaw stress enhances the sperm respiration or oxidative metabolism and excess formed electrons instead of completing the whole series of the electron transport chain in mitochondria, directly leak through mitochondrial inner membrane, and bind with oxygen resulting in the formation of free-radical superoxide (highly reactive molecule). Although ROS formation takes place during the whole cryopreservation procedure, excessive production of ROS during the freeze–thaw process mainly inflicts oxidative damage. Due to their highly reactive nature, they can combine readily with other molecules, directly causing oxidation that can lead to structural and functional changes and finally cellular damage. 57 All cellular components, including lipids, proteins, nucleic acids, and sugars are potential targets of oxidative stress. 56
ROS produced during the freeze–thaw process cause lipid peroxidation of sperm plasma membrane, which leads to the formation of a number of toxic by-products such as malondialdehyde (MDA), conjugated dienes, and lipid hydroperoxides (4-hydroxynonenal, 4- HNE and various 2-alkenals), which have damaging effects to the membrane system. 52 They decrease membrane fluidity, electrical resistance, and change the phase properties of membranes. Peroxidative attack on polyunsaturated fatty acids causes both structural and functional damage that is, the membrane becomes fragile and its semipermeable property is lost leading to membrane leaching. Cholesterol efflux, calcium influx, capacitation, and acrosomal reaction-like changes cumulatively depict cryocapacitation, which reduces viability and sperm fertility. 1 Membrane damage causes loss of adenosine triphosphate (ATP), proteins, and essential enzymes, which lead to reduction in motility of frozen–thawed spermatozoa. Toxic by-products also inhibit a large number of cellular enzymes such as glyceraldehyde 6-phosphate dehydrogenase (G-6-PDH) and adenosine triphosphatase (ATPase) that may also affect sperm movement. 58
Addition of anti-PDC-109 protein in semen ejaculates 59 and Met-anandamide (AEA) in freezing medium 60 successfully reduced cryocapacitation-like changes in frozen–thawed crossbred bull and ram spermatozoa, respectively, and thereby improved the post-thaw semen quality. Regucalcin (Senescence marker protein-30), a calcium-binding protein, also prevented cryocapacitation and played a cryoprotective role for buffalo bull spermatozoa. 61 The characteristics of membranes that affect their sensitivity include the cholesterol/phospholipid ratio, degree of hydrocarbon chain saturation, and protein/phospholipid ratio. 1
Along with plasma membrane damage, mitochondrial inner membrane and nuclear deoxyribonucleic acid (DNA) damage also occur due to ROS as these are more prone to oxidative damage. Sperm mitochondria are both the main source and target of ROS54,62 and have a role in many pathological implications of sperm cells. 63 Mitochondria are more likely to sustain damage from oxidative attack because mitochondrial DNA (mt DNA) is a circular double-stranded DNA organized in nucleoids near the electron transport chain (ROS source) and lacks introns. 64 DNA damage or mutation leads to defective protein formation during the process of transcription and translation, which affect spermatozoal metabolism. In humans, a level of 15%–25% spermatozoa with damaged DNA is considered to have reduced fertility,65,66 whereas bulls have been observed to have reduced fertility with over 10% damaged DNA spermatozoa. 67
Strategies to combat oxidative stress
Oxidative stress can be reduced either by neutralizing ROS, which are generated during semen cryopreservation process, or by minimizing the level of sources, which lead to ROS production (Fig. 1). Different strategies used to neutralize ROS, which are generated, are either addition of antioxidants or reductants (Fig. 1).
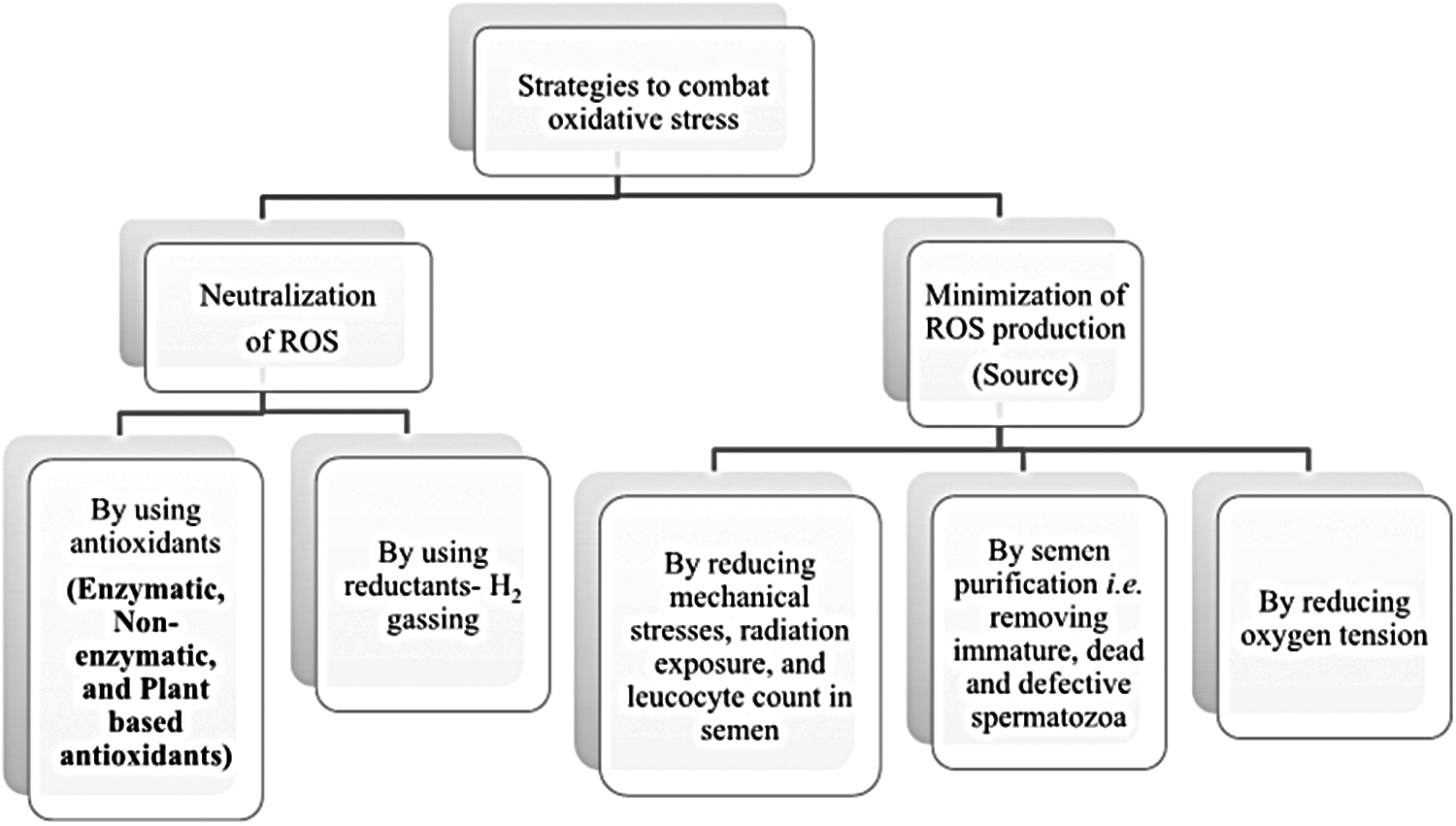
Schematic diagram showing different strategies to combat oxidative stress.
Antioxidants, in general, are the compounds and reactions which dispose, scavenge, suppress, or hinder the oxidative chain reaction in the formation of ROS, or oppose their actions. 68 Enzymatic antioxidants (natural antioxidants) include superoxide dismutase (SOD), catalase, glutathione peroxidase (GPx), and glutathione reductase (GR). 52 SOD dismutates superoxide anion (O2−) and forms O2 and hydrogen peroxide (H2O2); and catalase converts H2O2 to O2 and H2O. 51 The catalase enzyme has been reported to be present in cattle and ram spermatozoa, which control the aging process in sperm cells due to oxidative stress. 69 Addition of SOD and butylated hydroxytoluene to citrate–egg yolk extender caused the lowest production of MDA in frozen–thawed bovine semen. 70 Sperm viability was comparatively higher in samples extended with 100 U GPx/mL. 70
Nonenzymatic antioxidants, such as vitamin C and vitamin E, 71 ferrous sulfate and ascorbic acid, 72 vitamin B12, 73 astaxanthin, 74 canthaxanthin, 75 L-glutamine and L- proline, 76 inositol, 49 glutathione and cysteine, 77 cysteine and taurine, 55 methionine and raffinose, 78 raffinose and hypotaurine, 79 taurine and trehalose,69,80 trehalose and zinc sulfate, 81 cysteamine and hyaluronan, 69 manganese ion (Mn2+), 82 coenzyme Q10 (CoQ10) and ellagic acid, 83 melatonin, 84 and IGF-I, 85 have been successfully incorporated in semen extender to enhance the sperm cryosurvivability and post-thaw semen quality.
Some plant extracts having an antioxidant property such as pomegranate juice, 86 Rhodiola sacra aqueous extract, 87 Resveratrol, 88 Persimmon (Diospyros kaki) fruit extract, 89 Quercetin, 90 Lycopene, 77 Spirulina maxima extract, 91 Rosemary (Rosmarinus officinalis) extract, 92 Lycium barbarum, and Laminaria japonica plant extract 93 have been used in semen extenders to enhance post-thaw semen quality of mammalian species. El-Sheshtawy et al.94,95 concluded that pollen grains and honey improved preservability of bull semen due to its high content of antioxidants, flavonoids, vitamins, and organic acids. Supplementation of fresh bull semen with curcumin significantly increased the sperm content of GSH after thawing. 96 Maca (Lepidium meyenii) root powders or alcoholic extracts appeared to improve semen characteristics in men. 97 Plant extracts from African Guinea pepper (Xylopia aethiopica), Grape (Vitis vinifera) seed, and berry juices, which have a preservative effect, can be used as semen additives.98,99 Oleuropein, phenolic compound found in olives and olive oil, hydrolyzes to form hydroxytyrosol, which has potential antioxidant activity and may be used to improve semen quality. 100 Propolis, a resinous substance from plants decreased the percentages of dead and abnormal spermatozoa and increased testosterone level in rats. 101 Silymarin (milk thistle–Silybum marianum extract) as a natural additive to semen extenders improved preservability of frozen bull semen. 102
These antioxidants are efficient and reported to improve the post-thaw semen quality upon supplementation. But, many of the antioxidants function or their mechanism of action at the cellular level are not clearly known that is, how they act and at which steps of ROS production or oxidative damage in sperm cells? Also, are pro-oxidant activities of these antioxidant radicals formed after scavenging ROS? This question requires a better understanding of the dose–response relationship with respect to the efficacy and toxicity of antioxidants used for minimizing oxidative stress.
Reductants, such as the hydrogen molecule (H2), can easily diffuse across membranes and can react with cytotoxic •OH in all the organelles, including mitochondria and the nucleus, thus effectively protecting cells against oxidative damage. H2 also prevents a decrease in the cellular levels of ATP synthesized in mitochondria. 103 The rate of forward motility was increased by H2 treatment in experimentally damaged sperm suspensions from human patients. 104
The sources which lead to ROS production are radiation exposure, leucocytes, immature, defective, and dead spermatozoa; and oxygen tension present in microenvironment of spermatozoa (Fig. 1). Reducing the oxygen tension and minimizing the radiation exposure, leucocytes, dead and defective spermatozoa in the semen, and ROS production can be inhibited to a sufficiently low level (Fig. 1).
Radiation exposure should be minimized during transfer of semen from collection site to the laboratory and it should be built in a way to obviate direct exposure of sunlight during semen processing. Semen collection should be carried out from healthy, breedable bulls and not from diseased animals to minimize leucocyte count in semen and preputial washing should be done at regular intervals that is, 10 days or according to protocol followed at different semen stations. The bulls managed at a semen station should go through periodical testing and vaccinations as per the minimum standard protocol. Bulls should be tested against tuberculosis, Johne's disease, brucellosis, campylobacteriosis, trichomoniasis, and vaccinated against foot and mouth disease, hemorrhagic septicemia, black quarter, theileriosis and anthrax. However, vaccinations against bacterial diseases should be done only if there is an outbreak or prevalence of a particular disease in that particular area.
Methods of semen purification that is, removal of immature, dead, and defective spermatozoa are based on swim-up technique, Percoll density gradient centrifugation, Ficoll wash, glass wool-Sephadex (GWS) filtration, magnetic bead separation [magnetic-activated cell sorting (MACS) technique], and nanoparticle separation (semen nanopurification). Recovery of spermatozoa is based on sperm motility in overlaid media against the gravitational force (sperm swim-up), while recovery of spermatozoa by gradient centrifugation (Percoll) is based on separation of defective sperm as they were not able to pass through a layer of colloid during centrifugation. 105 Nowadays, PureSperm and OptiPrep have replaced Percoll in human IVF as these are optimized sperm gradient media and more effective in separation of damaged sperm. 106 Ficoll wash is an alternative to Percoll gradient centrifugation in livestock species. 107
Clusterin (CLU) from seminal plasma has affinity toward defective spermatozoa, therefore, CLU-positive frozen–thawed bull spermatozoa were retained in glass wool during GWS filtration, which improved the overall semen quality. 108 Leukosorb filters also served the same purpose as GWS for the removal of defective spermatozoa from stallion semen. 109 In MACS, magnetic beads conjugated with Annexin V bind to sperm with damaged plasma membranes, allowing for their depletion by a magnet. 110 Within livestock species, MACS was successfully applied to improve post-thaw motility of bull spermatozoa. 111
Semen nanopurification, a nanoparticle-based magnetic purification method, was developed to remove damaged or defective spermatozoa. Two types of iron oxide nanoparticles that is, particles coated with antibody against ubiquitin (present on the surface of defective spermatozoa), and particles coated with the lectin (peanut agglutinin) PNA (bind to glycans exposed by acrosomal damage in defective spermatozoa) were used for nanopurification of bull semen. 112 This approach enhanced bull semen quality before semen cryopreservation. A comparative efficacy of the nanotechnique with respect to the swim up technique for the purification of buffalo bull spermatozoa has recently been reported from our laboratory. 113 Nowadays, nanoparticles coated with antibody against damaged DNA and mitochondria are also being used in nanopurification of semen.
In these methods described for semen purification, the recovery rate of sperm is low in most of them as live or normal sperm also gets trapped along with dead and defective sperm during execution of these methods. Dead and defective sperm are low in good-quality semen; therefore, these techniques cannot be solely implied to improve the post-thaw semen quality as ROS produced due to cryopreservation stress cannot be minimized effectively. Some are not suitable for routine use in semen collection and cryopreservation due to cost, labor intensity, procedural time lags, and expensive equipment.
A novel approach for minimizing oxidative stress is to reduce the oxygen tension in sperm microenvironment as a number of literature pointed out a direct link of O2 with ROS production. In 1943, MacLeod 114 documented the toxicity of oxygen on human spermatozoa. The rate of lipid peroxidation and subsequent motility loss in sperm are linear functions of the partial pressure of O2 in the medium. 115 During the freezing–thawing process, semen is exposed to atmospheric oxygen, which in turn increases the ROS production and lipid peroxidation. 116 Because of the principal role of O2 in the formation of ROS, post-thaw sperm quality can be improved if the cryopreservation procedure is carried out under anaerobic conditions. 117
To achieve low O2 tension, Darr et al. and London et al.64,118 incorporated a commercially available Escherichia coli membrane preparation that is, Oxyrase, in the cryomedium, which improved post-thaw semen qualities of the stallion. It is a bacterial electron transport system that in the presence of a suitable hydrogen donor (i.e., lactate), decreases O2 level in cryomedia. 119 These bacterial electron transport chains (Oxyrase) act as acceptors for the excess electrons formed due to increased sperm metabolism as a result of freeze–thaw stress, to finally form harmless H2O instead of harmful O2−. The use of Oxyrase, and a mitochondrial uncoupler (2, 4-dinitrophenol) has been shown to reduce oxidative damage and improved the post-thaw semen quality of rhesus monkey. 120
Oxygen tension in the sperm microenvironment that is, extender, can also be reduced by partial deoxygenation process. In successive years, partial removal of oxygen from semen extender has been carried out using different approaches as reported from our laboratory that is, through flash-frozen/freeze-pump thaw cycle process using LN2, through a N2 gassing process/2–3 bubbles per second using gaseous N2 and through applying negative pressure to the diluent using modified vacuum pump. A flash-freezing process was used to degasify semen extender, which gave better results in semen quality parameters at post-thaw stage. 121 Balamurugan et al. 122 compared both the LN2 freezing method (by flash-freezing process) and mechanical method (by applying negative pressure) with that of a control to observe their effects on post-thaw semen quality of buffalo bull. But in both these methods, the dissolved oxygen (D.O.) in extender was not maintained at a desired level to observe its effect and to determine optimum oxygen level indicating better semen freezability. Therefore, we used the N2 gassing method and D.O. concentration was recorded with the help of a D.O. meter, before and after N2 gassing for varied time intervals. 123
Nitrogen gassing has been reported to improve the storage efficiency of Caprogen diluent at room temperature or at 5°C.124,125 Storage of diluted semen under N2 gas reduces the oxygen tension as well as metabolic activity of spermatozoa without affecting the pH of the medium. 125 Verberckmoes et al. 126 documented that storage of bovine liquid semen for up to 3 days in Caprogen diluent enables a 10-fold reduction in insemination dose without affecting sperm fertility. However, what is the D.O. concentration in normally used extender and how much N2 gassing should be done to displace oxygen to optimize the post-thaw semen quality? What should be the minimum D.O. concentration in semen extender essential for spermatozoa survival and metabolism? These were research gaps. The Egg Yolk-Tris-Glycerol (EYTG) extender kept in a water bath (35°C) had a D.O. concentration of 11.7 ppm (11–12 ppm), and it required N2 gassing for 16 and 9 minutes at 2–3 bubbles/sec to achieve D.O. concentration of ∼4 and 8 ppm, respectively. 123 The semen extender having 4 ppm of D.O. showed better post-thaw semen quality followed by 8 ppm.127,128 Adequate D.O. concentration in semen extender for spermatozoan survival and metabolism is 4–8 ppm, or more precisely 4 ppm as post-thaw semen quality was very poor in 2 ppm group. 128
In these methods of partial deoxygenation, approaches were used to minimize ROS production, but free radicals already present in the semen could not be effectively removed, which might affect the post-thaw semen quality. However, a procedural time lag of deoxygenation was also a limiting factor in the early processing of semen for cryopreservation. The techniques of quicker deoxygenation of the extender up to a desired dissolved O2 concentration would have shortened the processing time. When compared with other strategies, partial deoxygenation of semen extender with N2 gassing is a cost-effective, comparatively easy, and a potential approach to large-scale frozen semen production.
Conclusion
Advances in sperm cryopreservation techniques have taken place over time, and the directional freezing technique and sperm vitrification were proposed, as these techniques were found to be superior to conventional freezing in straws. However, conventional freezing techniques are still the most commonly used worldwide for semen cryopreservation of domestic species. During conventional cryopreservation, freeze–thaw damage occurs due to heterogeneous ice nucleation and ice recrystallization. To minimize these damages, AFPs were added in semen extender and beneficial effects were observed in terms of semen quality parameters. Apart from this, spermatozoa experience temperature stress (cold shock) and mainly oxidative stress during the whole cryopreservation process, which leads to structural and functional damages to sperm. One way to increase membrane fluidity and to prevent cold shock and cryocapacitation-like changes is the addition of cholesterol to the membrane in the form of CLC. Oxidative stress can be minimized either by neutralizing ROS through enzymatic, nonenzymatic, plant-based antioxidants or reductants; or by minimizing the level of sources like radiation exposure, leucocytes, and dead and defective spermatozoa, which lead to ROS production during the semen cryopreservation process. A novel approach of minimizing oxidative stress is the partial deoxygenation of semen extender to reduce the oxygen tension in sperm microenvironment. It is the process in which partial removal of oxygen from the extender is carried out through a flash-freezing process using LN2, N2 bubbling/gassing process, or applying negative pressure to the diluent using a modified vacuum pump. When compared with other strategies, partial deoxygenation of semen extender with N2 gassing has been found to be cost effective, comparatively easy, and a potential approach to large-scale frozen semen production.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.