
Abstract
In the present study, four experimental groups were used: fresh embryos, cultured during in vitro maturation and in vitro culture in media supplemented with bovine serum albumin (BSA) (fresh BSA) or fetal bovine serum (FBS) (fresh FBS); and two groups of cryopreserved and thawed embryos, produced under the same conditions (frozen BSA and frozen FBS). Experiment 1 evaluated the protein source effect on embryo development and response to cryopreservation. At day 7, half of the expanded blastocysts (Bx) from each group were cryopreserved and warmed and the other half were used as controls. After warming, embryos were incubated under the same conditions for 48 hours, and the hatching rate was measured at 24 and 48 hours. The total and the apoptotic cell numbers were measured in a subset of Bx after 24 hours. Experiment 2 used the Bx of experiment 1 to compare the expression of KRT8, PLAC8, FOSL1, HSP1A1, and HSPA5 genes in hatched blastocysts at 24 and 48 hours for all groups. The FBS group showed a higher percentage (p < 0.05) of embryos (42.8% vs. 27.9%) and higher rates of Bx (75.0% vs. 63.8%) on day 7, compared with the BSA group. At 24 hours postwarming, the fresh FBS group showed the highest hatching rate (p < 0.05) in comparison with other treatments. However, at 48 hours, the hatching rate was similar (p > 0.05) among groups: fresh FBS (68.1% ± 23.3%), fresh BSA (70.0% ± 31.0%), frozen FBS (39.2 ± 27.1), and frozen BSA (38.2 ± 23.9). After 24 hours, frozen BSA showed a higher number of cells compared with frozen FBS (p < 0.05). The expression of the PLAC8 gene was higher (p < 0.05) in fresh BSA embryos compared with frozen FBS embryos at 24 hours. In the present study, BSA replacement reduced embryo development, but did not affect the response to cryopreservation. However, upregulation of the PLAC8 gene suggests that embryos cultured in BSA might have better quality to support further development.
Introduction
Several studies have shown that embryos produced in vitro (IVP) are less resistant to cryopreservation than those produced in vivo, which has been associated with different physical and morphological characteristics of IVP embryos.1,2 Among other features, IVP embryos tend to contain a higher number of vacuoles, reduced expression of intercellular communication, less pronounced compaction among cells in the embryonic disc, and a lower number of cells compared with in vivo embryos.3,4 Moreover, they have a large intracellular accumulation of lipids and a decreased density of mature mitochondria. 5
Changes in cryopreservation techniques in the past decades have provided an increase in survival rates after thawing. However, an improvement in embryo cryoresistance is necessary to obtain results compatible with the commercial use of this technique.
6
Many strategies have been used to increase cryoresistance, such as the use of forskolin,
7
phenazine ethosulfate,
8
antioxidants,9,10
According to the literature, the main feature that affects the cryotolerance of IVP bovine embryos, which has been the subject of research in this area, is the high lipid content in the cytoplasm.15,16 However, it is not clear yet how and why lipid accumulation occurs in the cytoplasm of those bovine embryos. Such accumulation has been attributed to the presence of serum. In fact, the serum concentration affects the number and shape of lipid droplets in the cytoplasm of the IVP embryo.17,18 On the other hand, embryos cultured in serum-free systems showed a reduction in accumulation of lipid droplets and high resistance to cryopreservation. 19
Bovine serum albumin (BSA) is a source of protein that can be used as an alternative to embryo culture, considering that free fetal bovine serum (FBS) systems increase embryo cryotolerance. 18 Although there have been several studies using BSA supplementation in embryo culture media,20,21 studies carried out with both steps, in vitro maturation (IVM) and in vitro culture (IVC), in the absence of serum are still scarce. 22 It is noteworthy that besides the lack of information about embryo cryoresistance, when maturation and embryo culture are performed free of FBS, there is also little information on the use of the slow freezing method.
Most authors have reported positive results with IVP embryo cryopreservation using the vitrification method, 23 and it has been the most widely used technique for these types of embryos24,25 and oocytes.26,27 Although better results are obtained with vitrification for in vitro embryos, it requires the presence of a specialized technician both at the time of vitrification and in warming, which often complicates the logistics of IVP embryo transfer in farms. Therefore, to optimize the use of cryopreserved in vitro embryos, we propose to improve the results of slow freezing by using BSA supplementation not only during embryo development but also during oocyte maturation. We expected that by removing serum from culture, we would increase cryoresistance of in vitro embryos, allowing their direct transfer. Thus, embryos developed in different systems (BSA vs. FBS) were evaluated (after thawing) for reexpansion and hatching, total number of cells and apoptotic cells, and expression of selected genes related to embryo quality.
Materials and Methods
Chemicals and supplies
Unless otherwise indicated, chemicals were purchased from Sigma (St. Louis, MO).
Oocyte recovery and IVM
Ovaries from crossbred cows (Bos indicus x Bos taurus) were collected immediately after slaughter and transported to the laboratory in saline solution (0.9% NaCl) supplemented with penicillin G (100 IU/mL) and streptomycin sulfate (100 g/mL) at 35°C. Cumulus–oocyte complexes (COCs) were aspirated from 3 to 8-mm diameter follicles with an 18-gauge needle. Only COCs with homogeneous cytoplasm and at least three layers of cumulus cells were used. Selected COCs were washed and transferred in batches of 25–30 to a 200-μL drop of maturation medium under silicone oil and incubated for 24 hours at 39°C with 5% CO2 in the air. The maturation medium consisted of TCM-199 (Invitrogen, Carlsbad, CA) supplemented with 10% of FBS or 0.4% of BSA fatty acid free (BSA-FAF), 0.01 IU/mL of follicle stimulating hormone, 0.1 mg/mL of
In vitro fertilization and embryo IVC
After maturation, COCs (groups of 25–30) were transferred to a 200-μL drop of fertilization medium. For fertilization, frozen semen from a Nellore bull previously tested in the laboratory for in vitro fertilization was used. Motile spermatozoa were obtained by the Percoll method 28 and they were added to droplets containing COCs at a final concentration of 1 × 106 spermatozoa mL−1. Tyrode's albumin lactate pyruvate (TALP) medium was used as the fertilization medium 29 supplemented with penicillamine (2 mM), hypotaurine (1 mM), epinephrine (250 mM), and heparin (10 μg/mL−1). Spermatozoa and oocytes were coincubated for 18 hours at 39°C with 5% CO2 in the air, and the day of in vitro insemination was considered as day 0. After coincubation, the presumptive zygotes (n = 25–30) were washed and transferred to 200-μL drops of SOFaaci medium, 30 supplemented with 2.77 mM of myo-inositol and 5% FBS or 0.4% BSA-FAF, and cultured at 39°C with 5% CO2 in the air for 8 days. Embryos were evaluated on day 2 postinsemination (pi) for cleavage and on days 6 and 7 pi for the blastocyst rate.
Embryo cryopreservation using slow freezing
Expanded blastocysts (Bx) with quality I and II 31 were cryopreserved by the slow freezing method according to Stinshoff et al., 2011. 32 Briefly, Bx were exposed to the freezing solution consisting of 1.5 M ethylene glycol (EG) (Vitaflex, Brazil) at 36°C for 10 minutes. Embryos were loaded into 0.25-mL straws, placed on a central column of a 1.5 M solution of EG, surrounded by two other columns of the EG, and separated by air columns from each other. After being loaded, the straws were placed in a freezing machine (FREEZE CONTROL®, Model CL-863 System; Cryologic, Australia) that had been previously stabilized at −6°C. Two minutes after being placed in the machine, crystallization (seeding) of the columns immediately above and below the embryo column was conducted. Embryos remained at −6°C for 10 minutes.
The freezing curve was initiated, lowering the temperature by 0.5°C per minute to −35°C. At the end of the freezing curve, the embryos were immersed directly in liquid nitrogen.
Thawing and culture postcryopreservation
After freezing, the straws were removed from the LN2, held for 5 seconds at room temperature, and then immersed in water at 30°C for 30 seconds. After thawing, the embryos were washed three times in DPBS (Nutricell, Campinas, São Paulo, Brazil) and transferred to synthetic oviductal fluid (SOF) medium under the same conditions they had been before cryopreservation. Embryos were evaluated for expansion and hatching rate at 24 and 48 hours post-thaw. The control groups of both treatments remained on the bench in holding medium during freezing and thawing. Afterward, they were washed and transferred to the same culture medium drop and placed back into the incubator (FBS or BSA).
Embryo collection for gene expression
Gene expression was performed at two different time points, first at 24 and then at 48 hours after thawing. At both time points, only the hatched blastocysts were used. Embryos from each group were washed four times in PBS that was calcium and magnesium free, then stored in microtubes with RNAlater at −20°C until RNA extraction.
Six embryos from each group were evaluated. For expression quantification, the genes selected were Keratin 8, Type II (KRT8) and Placenta-Specific 8 (PLAC8) related to embryo quality, FOS-like Antigen 1 (FOSL1) related to apoptosis, and heat shock protein A1 (HSP1A1) and heat shock protein A5 (HSPA5) related to heat stress. The peptidylprolyl isomerase A (PPIA) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) genes were used as constitutive genes.
mRNA extraction and cDNA synthesis
Total RNA was isolated using the RNeasy Plus Micro Kit (Qiagen®, Hilden, Germany) according to manufacturer's instructions (with minor modifications). The isolated total RNA was used for cDNA synthesis, which was performed using the First-Strand cDNA Synthesis Kit (Invitrogen)—SuperScript® III (200 U/μL) and oligo(dT) primer (0.5 μg/μL) in a final volume of 40 μL. Reactions were performed at 65°C for 5 minutes and 50°C for 50 minutes, followed by inactivation of the enzyme at 85°C for 5 minutes. The quantitative polymerase chain reaction (qPCR) analysis was performed using a Fast SYBR Green Master Mix (Applied Biosystems). The reactions were optimized to provide maximum amplification efficiency for each gene (76%–110%) by calculations using standard curves relating to the program 7500 2.0.3 (Applied Biosystems). Each sample was analyzed in triplicate, and the specificity of each PCR product was determined by analysis of the melting curve and amplicon size on agarose gel. The reactions were performed in a final volume of 25 μL, using cDNA corresponding to the 0.33 fraction of an embryo. The conditions of PCR cycles were 95°C for 5 minutes, followed by 50 cycles of denaturation at 95°C for 10 seconds, and annealing at 60°C for 30 seconds. The name, primer sequence and concentration, amplicon size, and annealing temperature of each gene are shown in Table 1.
Information of Specific Primers Used for Amplification of Gene Fragments for Real-Time Quantitative Polymerase Chain Reaction Analysis
F, primer forward; FOSL1, FOS-like Antigen 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HSP1A1, heat shock protein A1; HSPA5, heat shock protein A5; KRT8, Keratin 8, Type II; PLAC8, Placenta-Specific 8; PPIA, peptidylprolyl isomerase A; R, primer reverse.
The expression levels of the two constitutive genes, GAPDH and PPIA, were subjected to the geNorm analysis program. The GAPDH level was used as a reference for normalization of data. The relative expression of each gene was calculated using the ΔΔCt method with efficiency correction by the Pfaffl method. 33
Total cell number and apoptotic cell ratio (terminal deoxynucleotidyl transferase dUTP nick end labeling)
To determine the total cell number and apoptotic cell ratio, a subset of expanded blastocysts was removed on day 8 and stained with terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) and Hoechst 33342. Blastocysts were washed in warm PBS (Life Technologies) supplemented with polyvinylpyrrolidone (PVP) (1 mg/mL) before fixation in 4% paraformaldehyde for 1 hour. All incubation steps occurred at room temperature in the dark unless otherwise noted, and embryos were washed in 1 mg/mL of PVP between each incubation step. After washing with 1 mg/mL of PVP, the blastocysts were incubated in 0.5% Triton X-100 for 60 minutes. Subsequently, the positive (artificial DNA denaturation, TUNEL, and Hoechst 33342 staining) and negative (artificial DNA denaturation and Hoechst staining) controls were incubated with 50 U/mL DNase (Roche, Vilvoorde, Belgium) for 1 hour. Blastocysts (except negative controls) were then stained with a TUNEL enzyme-labeling mix (Roche) for 60 minutes at 37°C and Hoechst 33342 staining for 10 minutes. Finally, the blastocysts were washed in PVP, mounted on glass slides, and observed under a fluorescence microscope. For each blastocyst, the individual total cell number (blue nuclei, Hoechst 33342) and the total number of apoptotic cells (green nuclei, TUNEL) were determined. The apoptotic cell ratio was defined as the percentage of apoptotic cells of the total cell number.
Experimental design
Two experiments were performed to evaluate the effect of the protein source, during oocyte maturation and embryo culture, on resistance to cryopreservation and quality of in vitro produced bovine embryos.
Experiment 1—embryo development and resistance to cryopreservation
This experiment aimed to determine whether the protein source, used for supplementation in culture media, affects the quantity and quality of embryos and their response to cryopreservation. A total of 1302 grade I and II oocytes from ovaries obtained from a slaughterhouse were used in 11 replicates. After selection, the COCs were distributed into two treatments: fresh FBS (control): the media were supplemented with FBS during IVM (10%) and IVC (5%); and fresh BSA (BSA): the media were supplemented with BSA during IVM (0.4%) and IVC (0.4%). Cleavage rates and blastocyst development were evaluated on days 2 and 7, respectively. To evaluate the response to cryopreservation, expanded blastocysts classified as grade I (symmetrical embryo blastomeres of the same size, uniform color, and without the extrusion of cells) from each treatment were divided into fresh and frozen groups. Then, the embryos were evaluated at 24 and 48 hours for hatching and reexpansion in IVC media of the four experimental groups. Besides embryo kinetics, the total cell number and number of apoptotic cells were also assessed.
A total of 443 embryos in the expanded blastocyst stage were used for cryopreservation, on day 7, produced using BSA (n = 194) or FBS (n = 279). Half of the embryos from each group were submitted to slow freezing (frozen) and the remaining ones were used as controls (fresh). After freezing/thawing, embryos were cultured for 24 hours and then used for TUNEL evaluation at the moment that transfers would be made to determine the total number of cells and the total number of apoptotic cells.
Experiment 2—gene expression of fresh and frozen embryos produced with FBS or BSA supplementation during IVM and IVC
For the second experiment, the objective was to compare the gene expression of fresh or frozen/thawed embryos produced in a medium supplemented with different sources of protein (BSA or FBS). The embryos produced in experiment 1 were evaluated on day 7 of development and only expanded blastocyst (Bx) quality I, according to the classification of the International Embryo Technology Society, 31 were used. During cryopreservation, control embryos were maintained in holding media (TCM 199-Hank's) supplemented with the same source of protein used during all the steps of IVP over the heating plate at 36°C. After warming the frozen groups, the embryos of the four experimental groups were cultured in the SOF medium for additional 24 and 48 hours. Embryos that had hatched after 24 hours were removed from culture, while the remaining ones were kept in the incubator until completing 48 hours. At 24 and 48 hours, the hatched embryos were stored individually in RNAlater at −20°C for further molecular analysis. For the gene expression assay, six embryos were used for each of the four experimental groups at 24 and 48 hours. The evaluated genes were KRT8, PLAC8, FOSL1, HSP1A1, and HSPA5.
Statistical analyses
All data were analyzed using Statistical Analysis System software (SAS, 1999). Cleavage, fertilization, and blastocyst development were compared according to χ 2 (p < 0.05). ANOVA was used to determine the total cell and apoptotic cell numbers, and the means were compared by Tukey's test (p < 0.05). Comparison of gene expression among groups was performed using one-way ANOVA or the Kruskal–Wallis test, and means were compared by Tukey's test, depending on data distribution (p < 0.05). Data are expressed as mean ± standard error of the mean.
Results
Experiment 1—embryo development and resistance to cryopreservation
The cleavage rate was similar (p > 0.05) between the two systems (Table 2). However, the group in which the oocytes and embryos were cultured with BSA showed the lowest (p < 0.05) blastocyst production on day 7. The percentage of expanded blastocysts was also lower (p < 0.05) than those produced with FBS.
Number (n) and Percentages (%) of Cleaved at D2, Blastocyst at D7, and Hatched Blastocyst at D7 of In Vitro Matured Oocytes and In Vitro Cultured Embryos in the Presence of Fetal Bovine Serum or Bovine Serum Albumin
Values with different superscripts in the same column are significantly different by χ 2 test (p < 0.05).
Expanded blastocyst rates are calculated according to the total of blastocysts at D7.
BSA, bovine serum albumin; FBS, fetal bovine serum.
At 24 hours after thawing, the frozen BSA group presented similar reexpansion and hatching rates and did not differ from fresh BSA embryos. At 48 hours, the hatching rate was similar (p > 0.05) for BSA and FBS in the fresh groups (Table 3). Similar results were observed when comparing BSA and FBS frozen groups (Table 3). However, frozen groups had lower hatching rates (p < 0.05) than the fresh groups, regardless of the protein source. At both times points (24 and 48 hours after thawing), the FBS and BSA groups were similar regarding degenerated and viable rates. The only difference observed for the degenerated and viable embryo rates was between BSA frozen and FBS fresh groups.
Evaluation at 24 and 48 Hours of Reexpansion—Hatching and Degeneration Rates of Fresh or Frozen Thawed Bovine Embryos Produced In Vitro in the Presence of Fetal Calf Serum or Bovine Serum Albumin
The data are presented as numbers (n) and percentages (%).
Values with different superscripts in the same column are significantly different by χ 2 test (p < 0.05).
After 24 hours, embryos from the frozen BSA group showed a higher total cell number compared with the frozen FBS group (p < 0.05). However, the number of apoptotic cells was also higher (p < 0.05) for the BSA group (Table 4). Fresh control groups, both BSA and FBS, were similar (p > 0.05) regarding the total cell number, and the frozen BSA group had a higher (p < 0.05) number of apoptotic cells (Table 4).
Evaluation of Total and Apoptotic Cell Numbers (n) and Standard Deviations, as the Ratio of Both (%), for Fresh and Frozen Thawed Blastocysts 24 Hours After Warming
The blastocysts were in vitro produced in the presence of FBS or with BSA.
Values with different superscripts in the same column are significantly different by Tukey's test (p < 0.05).
SD, standard deviation.
Experiment 2—gene expression of fresh or frozen embryos produced with FBS or BSA supplementation in IVM and IVC
There was a higher (p < 0.05) expression of the PLAC8 gene (Fig. 1) in the fresh group on day 8 for hatched embryos (24 hours) produced with BSA, compared with the frozen FBS embryos (p < 0.05). For the other evaluated genes (FOSL1, KRT8, HSP1A1, and HSPA5), no differences (p > 0.05) regarding the expression pattern were detected between groups, regardless of the time, at 24 (Fig. 1) or 48 hours (Fig. 2).
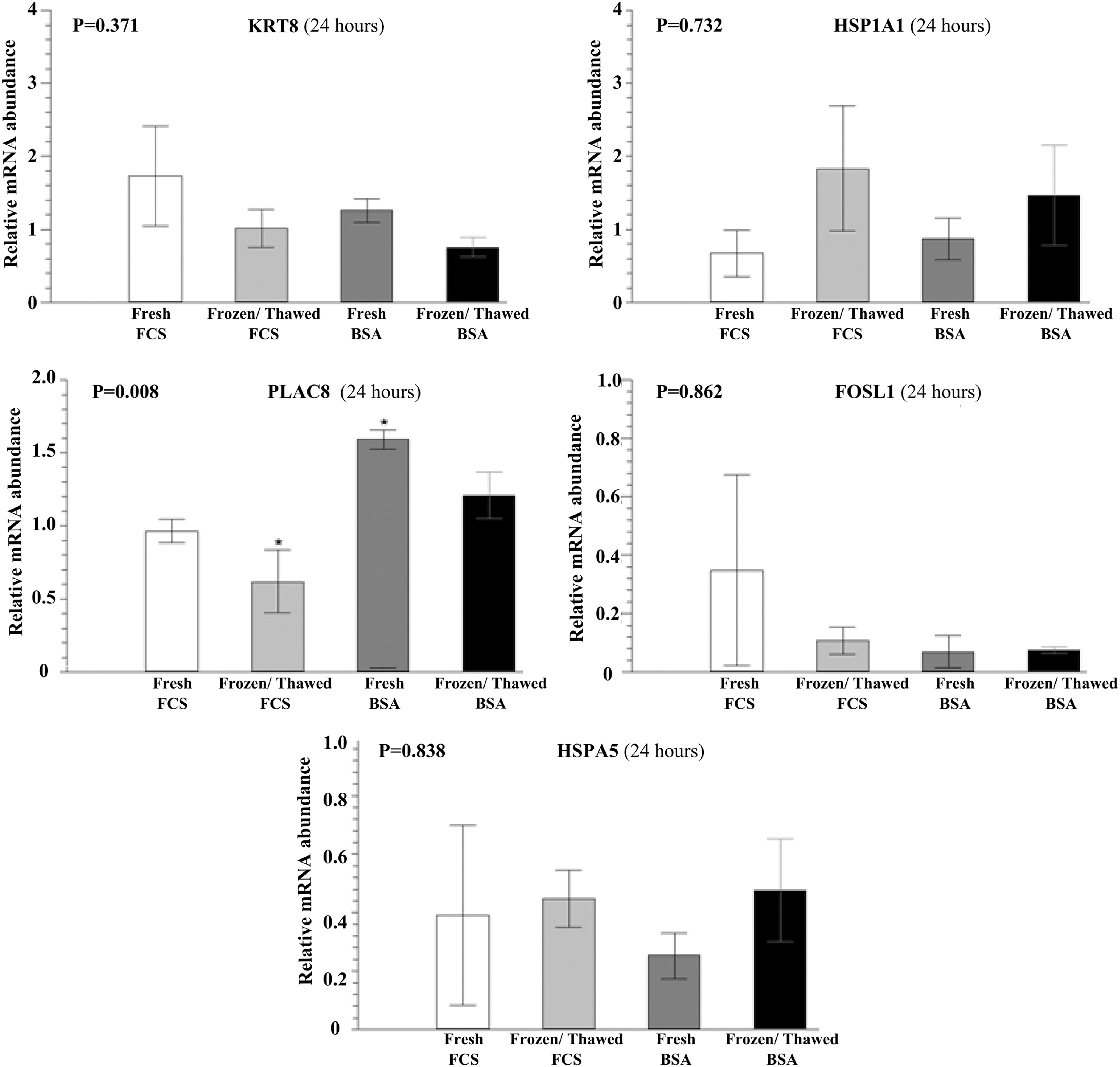
Relative abundance of mRNA encoding KRT8, HSP1A1, PLAC8, FOSL1, and HSPA5 determined by qPCR at 24 hours in post-thawed hatched bovine blastocysts. Mean ± SEM of three biological replicates. The data (mean ± SEM) were normalized using the ΔΔCt method, with GAPDH as an endogenous control. *Indicates significant differences between treatments obtained by ANOVA or the Kruskal–Wallis test, and means were compared by Tukey's test, depending on data distribution. BSA, bovine serum albumin; FBS, fetal bovine serum; FOSL1, FOS-like Antigen 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HSP1A1, heat shock protein A1; HSPA5, heat shock protein A5; KRT8, Keratin 8, Type II; PLAC8, Placenta-Specific 8; qPCR, quantitative polymerase chain reaction; SEM, standard error of the mean.
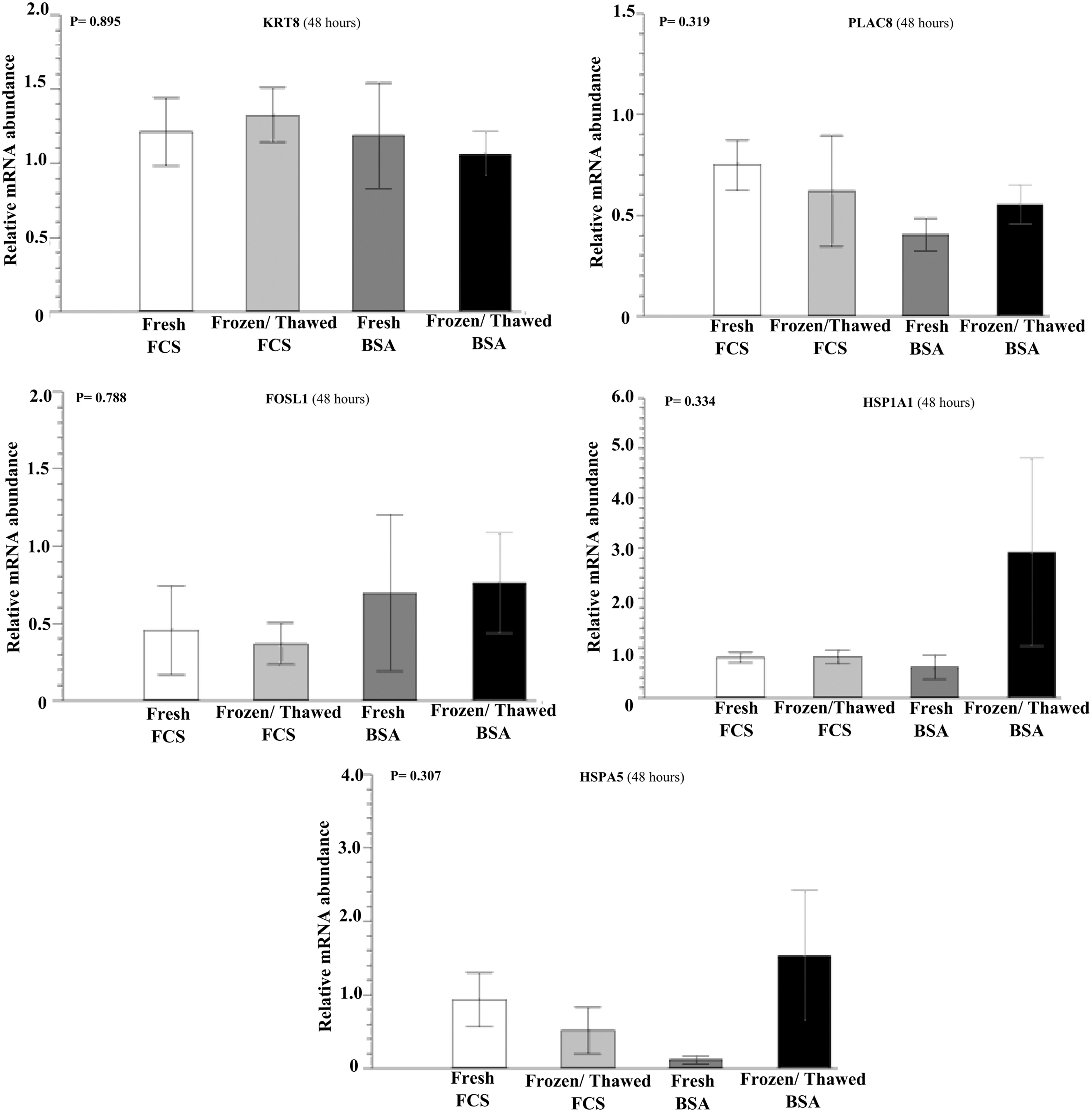
Relative abundance of mRNA encoding KRT8, HSP1A1, PLAC8, FOSL1, and HSPA5 determined by qPCR at 48 hours in post-thawed hatched bovine blastocysts. Mean ± SEM of three biological replicates. The data (mean ± SEM) were normalized using the ΔΔCt method, with GAPDH as an endogenous control. Data were analyzed by ANOVA or the Kruskal–Wallis test, and means were compared by Tukey's test, depending on data distribution.
Discussion
One of the disadvantages of using IVP is the difficulty to cryopreserve the embryos. Several studies have suggested that the main factor responsible for the lower cryoresistance of IVP embryos is intracellular accumulation of lipids. 34 Although it is not very clear what causes accumulation of lipids, there are many pieces of evidence indicating that this fact is directly related to medium supplementation with FBS. 18
During oocyte maturation and embryo culture, BSA can be used as an alternative when it is intended for use in cryopreservation. Associated with the use of BSA, the slow freezing method was chosen to cryopreserve the produced embryos. This method has the advantage of making possible the transference of embryos after they have been thawed in the straw, facilitating the dissemination and logistics of embryo transfer. Therefore, this study was designed to focus on those two aspects: (1) the protein source replacing FBS by BSA during oocyte maturation and embryo culture and (2) the slow freezing method for cryopreservation of IVP embryos.
Several studies have shown that the presence of serum during embryo culture produces more embryos than when it is replaced by BSA or other macromolecules.35,36 However, in many studies, the protein source is only changed during embryo development, and studies on the use of both steps, IVM and IVC, in the absence of serum are still scarce.20,22 Considering this reduction in blastocyst development, due to serum removal from the system, the results of the current study (almost 30% of blastocysts on day 7) demonstrate that the system efficiency is higher than that observed in other studies cited above.
When the speed of embryo development was evaluated, it was observed that FBS embryos had higher expansion rates on day 7, compared with the BSA ones. These results confirm the existence of the biphasic effect of FBS, which inhibits early cleavage divisions and accelerates development of morula to the blastocyst stage. 4 The serum stimulation mechanism at this stage of embryonic development is unknown, but this stimulation is related to growth factors and/or amino acids, or also to its antioxidant properties, which reduce the formation of superoxides. 37 These features induce a higher production of embryos reaching the blastocyst stage earlier. The literature assumes that the speed of development of IVP embryos is related to its quality and therefore embryos that develop earlier are more likely to generate pregnancy. 38 It could be assumed that embryos from the FBS group have better quality than those from the BSA group. However, it should be taken into account that the BSA embryos may have a deficiency in stimulating elements for blastocyst development; these embryos have slower cell cycles and reach the blastocyst stage later. 39 Therefore, in this study, the development speed may not be a good parameter of embryo quality.
In experiment 1, the behavior of embryos cultured in media supplemented with FBS or BSA was also evaluated after freezing. It was observed that at 24 hours of culture post-thawing, the frozen FBS group had a higher rate of reexpansion and hatching than the other groups. However, at 48 hours after thawing, the hatching rate was similar (p > 0.05) for BSA and FBS in fresh (control) groups. A similar pattern was observed in the frozen group. This finding differs from that by Gómez et al., 34 who found that the hatching rate at 24 and 48 hours after vitrification was higher in embryos cultured in BSA. Since we used only quality I embryos, according to the classification of the International Embryo Technology Society (IETS), for both groups, the embryos showed similar ability to withstand cryopreservation.
In the evaluation of the total cell number of produced embryos in the two systems, it was observed that at 24 hours of culture, fresh FBS embryos showed many cells similar to fresh BSA embryos and a larger number of cells in comparison with frozen groups. These data reinforce the findings by Nedambale et al., 40 who observed that the addition of FBS causes an increase in the total cell number compared with nonsupplemented groups. The fresh FBS group had a lower number of apoptotic cells than the other groups, whereas the frozen BSA group had a higher number of apoptotic cells. These observations are different from those described by Sudano et al., 8 who found that the number of apoptotic cells steadily increased with FBS medium supplementation. The variation between these results may have occurred mainly due to differences in embryo culture systems and cryopreservation methods used in each study. Apoptosis plays an important role in embryonic development and cellular homeostasis, acting as a quality control mechanism for removal of cells that are damaged, nonfunctional, abnormal, and excessive in number. 41 Thus, the number of apoptotic cells might be used to evaluate the embryo quality and based on that we conclude that FBS have beneficial effects during IVM and IVC.
Among all evaluated genes, only PLAC8 expression was affected by the treatments. The hatched embryos from the fresh BSA group showed a higher gene expression for PLAC8 on day 8. The PLAC8 gene is an important marker of embryo viability and it is usually used to predict quality in IVP programs. 42 In domestic animals, the PLAC8 gene does not have a well-established pathway, but it is known that its expression is related to placental development, 43 playing a role in maternal–fetal communication. 44 A high gene expression was reported in bovine blastocysts that succeeded in pregnancy, compared with those that had been reabsorbed, 45 and in the endometrium of pregnant cows compared with nonpregnant ones. 46 The results suggest that despite a higher number of apoptotic cells, supplementation of IVM and IVC with BSA generates embryos that after cryopreservation might be more likely to establish pregnancy after transfer.
In conclusion, we found that supplementation with BSA during IVM and IVC negatively affects the developmental competence of embryos. Therefore, the use of BSA as a replacement of FBS did not improve resistance to cryopreservation. However, we found that BSA supplementation preserved the PLAC8 expression even after cryopreservation, an indication that these embryos might have better chances to establish pregnancy after transfer. All observations together suggest that the benefits of the use of BSA during IVM and IVC are still not clear. Finally, we conclude that when good IVP embryos are used, slow freezing is efficient regardless of the protein source.
Footnotes
Acknowledgment
The authors thank the Qualimaxima slaughterhouse (Brasilia-DF, Brazil) for providing the ovaries used in the experiment.
Author Disclosure Statement
The authors declare no conflict of interests.
Funding Information
The first author received a scholarship that was supported by CAPES, Brazil.