
Abstract
Carboxylated poly-
Introduction
Water buffalo contributes >10% in global milk production and the increase in the buffalo population in the world has been reported as 88% between the years of 1970 and 2010. 1 Buffalo milk has a high fat to protein and casein to protein ratio, which makes it suitable for cheese making. 2 However, the production potential of buffalo is quite low compared to cattle, which can be improved through maximum utilization of assisted reproductive techniques, specifically through artificial insemination (AI). 3 AI allows rapid genetic improvement through exploiting the germplasm of superior bulls. Although the success of AI depends upon successful cryopreservation of semen, freeze-thaw cycles have deteriorating effects on sperm function. 4 Cold shock is one of the major stressors that disrupts the plasma membrane 5 and induces premature acrosomal reaction, oxidative stress, and loss of phospholipids, which ultimately affect motility, viability, and fertility of the sperm.3–6
The osmotic changes during the freeze-thaw process result in the formation of ice crystals at continuously decreasing temperatures. Ultimately, the cells become squashed between ice crystals, exposed to lethal concentrations of salts and dehydrated. 7 Polge and colleagues discovered the cryoprotectant properties of glycerol, a small molecule that penetrates the cell membrane, remains inside, and prevents excessive dehydration during freezing. 8 The glycerol is generally used at 7% (v/v) in buffalo semen extender.3,4 Even though it is an efficient cryoprotectant, the major problem in its use is its cytotoxicity, osmotoxicity, and contraceptive effect, which has been reported in human, pig, chicken, and turkey spermatozoa.9,10 In this regard, different studies have been conducted to replace or reduce the level of glycerol in semen extender with other permeating cryoprotectants such as dimethyl sulfoxide.9–11 Unfortunately, dimethyl sulfoxide has not gained popularity in semen cryopreservation industry due to contradictory results and its toxic and osmotic effects on buffalo spermatozoa.9–11 Therefore, exploration of alternate cryoprotectants with low cytotoxicity is needed.
The polyampholytes with an appropriate ratio of amino acid and carboxyl groups have antifreeze protein (AFP)-like properties and also have good cryoprotective efficiency. The successful cryopreservation of different cells can be done with these polyampholytes without the addition of any other cryoprotective agent in the media. One of these polyampholytes with AFP-like properties is carboxylated poly-
Materials and Methods
Ethical statement
The study was approved by Ethics Committee of Pir Mehr Ali Shah Arid Agriculture University Rawalpindi, Pakistan, for the use of animals.
Carboxylation of poly-l -lysine solution
CPLL solution (0.65) was prepared as described by Matsumura and Hyon.
12
For this purpose, poly-
Preparation of experimental extenders
Different experimental extenders were prepared as described in Table 1. After preparation, pH and osmolality were measured for all the extenders.
Composition of Experimental Extenders for Buffalo Semen Cryopreservation
Study animals, semen collection, and cryopreservation
Semen was collected from four regular donor Nili-Ravi buffalo bulls of similar age (5–6 years), which were housed under uniform standard conditions at the Semen Production Unit, Qadirabad, Sahiwal, Punjab, Pakistan. Two consecutive semen ejaculates were collected from each bull for 5 consecutive weeks (replicates) using an artificial vagina (42°C), and placed in water bath (37°C) until initial evaluation. Sperm mass activity was evaluated by placing 5 μL of a neat semen sample on a prewarmed glass slide (37°C) and observing under a phase contrast microscope (Olympus BH-2, 400 × magnification). Mass activity was graded on a scale ranging from 0 to 5 where 0 represents no motion and 5 represent up to 100% progressive motion of semen. Sperm motility was assessed by placing 5 μL of semen sample on a microscopic slide (37°C) and covering it with a prewarmed coverslip (37°C). Subjective assessment of sperm motility was carried out using a standard scale ranging from no motile sperm to >90% motile sperm. 18 Sperm concentration was measured by a Bovine Photometer (ACCUCELL®; IMV Technologies, France) at 540 nm wavelength. After initial evaluation, semen ejaculate from each bull presenting a volume up to or greater than 1 mL, motility greater than 70%, and sperm concentration greater than 0.5 billion sperm/mL was pooled to eliminate the bull effect and incubated at 37°C for 15 minutes before further processing. Pooled semen samples from four experimental bulls were aliquoted and diluted with different experimental extenders at 37°C to attain the final sperm concentration of 50 million sperm/mL. The extended semen samples were slowly cooled to 4°C within 2 hours, equilibrated for 4 hours (4°C), and filled into French straws (0.5 mL; IMV) at 4°C in a cold cabinet. The French Straws were frozen in a programmable automatic freezer (Digit Cool; IMV Technologies) with programming set for 2 minutes of initial holding of straws at +4°C, freezing at the rate of 10°C/min from +4°C to −10°C, 15°C/min from −10°C to −20°C, −60°C/min from −20°C to −120°C, and the final holding for 30 seconds at −120°C. 10 Finally, the straws were plunged into liquid nitrogen (−196°C) until evaluation and thawing of straws was done at 37°C in a water bath for 30 seconds.
Semen quality assessment assays
Sperm motility
Sperm motility was assessed by placing 5 μL of semen sample on a microscopic slide (37°C) and covering it with a prewarmed coverslip (37°C). Spermatozoa from five different microscopic fields were evaluated by a phase contrast microscope (Labomed LX400, USA) at 400 × . Subjective assessment of sperm motility was carried out using a standard scale ranging from no motile sperm to >90% motile sperm.2,3,18
Sperm plasma membrane integrity
Plasma membrane intactness of sperm was evaluated using the hypo-osmotic swelling test (HOST). 19 For this purpose, 0.735 g of sodium citrate (Merck, Germany) and 1.351 g fructose (Scharlau, Spain) were dissolved in 100 mL of distilled water to make the HOST solution. A semen sample (50 μL) and HOST solution (500 μL) were mixed and placed in water bath for 30 minutes at 37°C. An incubated sample of 10 μL was placed on a prewarmed glass slide (37°C) and covered with a cover slip (37°C). The slides were randomly selected and observed using a phase contrast microscope (Labomed LX400, USA) at 400 × magnification and a total of 100 sperm were assessed in different microscopic fields. Swollen and bent tailed spermatozoa signify intactness of plasma membrane, while the straight and unswollen tailed spermatozoa signify damaged plasma membrane. 18
Sperm viability and livability
Sperm acrosome integrity
The sperm normal apical ridge was evaluated by fixation of thawed semen samples in formal citrate solution (2.9 g trisodium citrate dihydrate [Merck] and 1 mL of formaldehyde solution [37%] [Merck] in 99 mL of distilled water) and observed under a phase contrast microscope (Labomed LX400, USA) at 1000 × magnification. A total of 100 sperm were selected randomly from different fields and evaluated. Sperm with clear normal apical ridge were considered normal, whereas those with a blurred acrosome region were considered abnormal. 21
Sperm chromatin integrity
Sperm chromatin integrity of buffalo spermatozoa was assessed by acridine orange test modified by Shah et al. 11 and formerly described by Martins et al. 22 A semen smear of each experimental extender was prepared on microscopic slides (37°C), allowed to air dry at room temperature, and fixed in Carnoy's solution (glacial acetic acid and methanol, 3:1 proportion; Merck) for 2 hours at room temperature. After fixation, air-dried microscopic glass slides were placed at 75°C for 5 minutes in Tampon solution (15 mmol/L Na2HPO4·7H20; 80 mmol/L citric acid; pH 2.5; Merck). Finally, the slides were dipped in freshly prepared acridine orange (0.02%; Sigma-Aldrich) for 2 to 3 minutes, washed with distilled water, and placed under a cover slip in wet condition. A total of 200 spermatozoa form different microscopic fields were evaluated under an epifluorescence microscope (Labomed LX400, USA) equipped with an excitation filter (480) and barrier filter (550 nm) at 400 × magnification. Spermatozoa having damaged DNA emitted yellow to red fluorescence, while spermatozoa having intact DNA emitted green fluorescence. 22
Lipid peroxidation
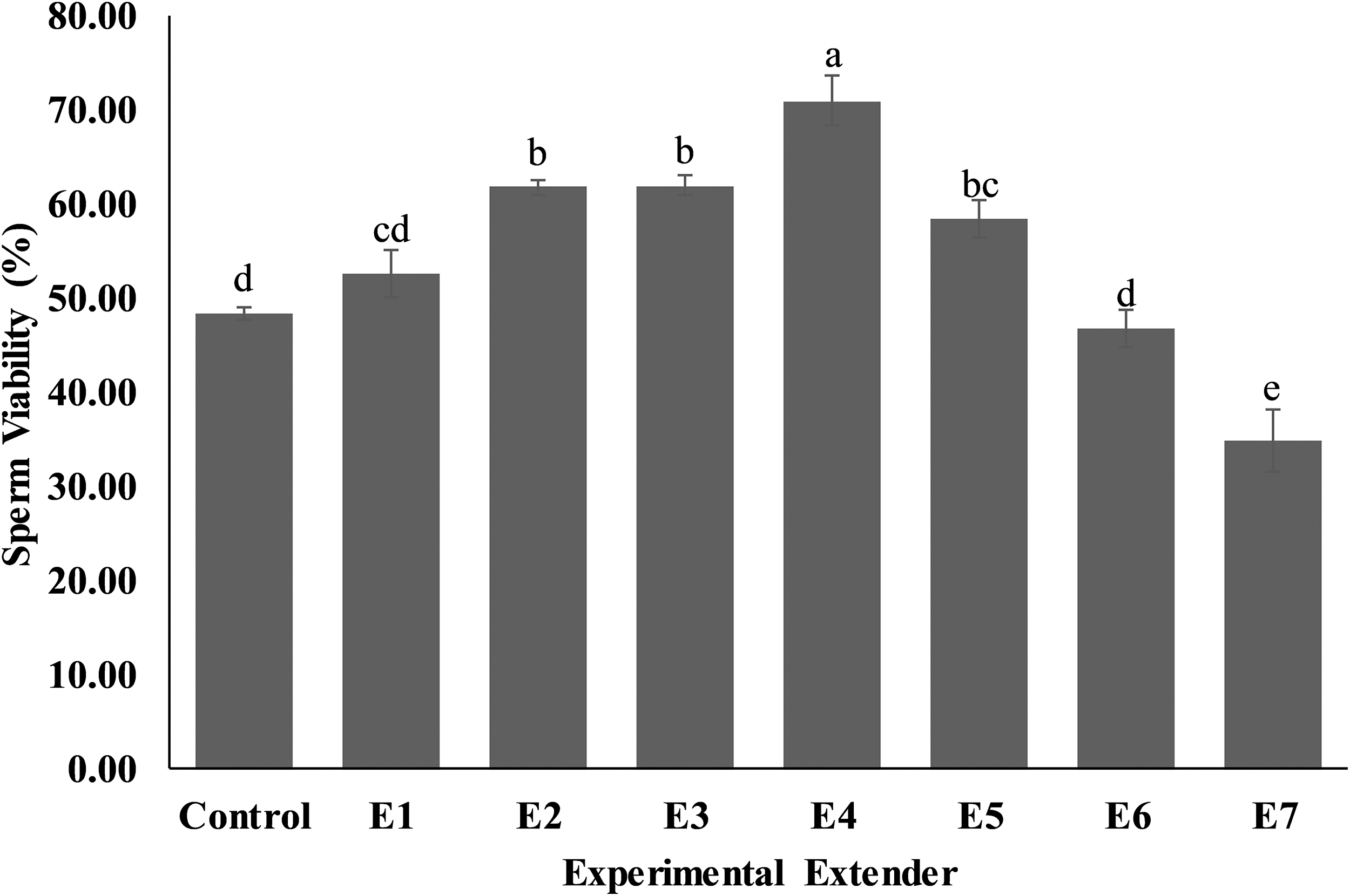
The thiobarbituric acid assay (TBA) was performed to assess malondialdehyde (MDA) concentration of sperm as indices of lipid peroxidation. 23 The thawed semen sample was centrifuged at 1500 × g for 10 minutes and the supernatant was discarded. The sperm pellet was washed with phosphate-buffered saline (PBS) (pH 7.4 at 25°C; Sigma-Aldrich), centrifuged, and the supernatant was discarded. Finally, the sperm pellet was diluted in 2 mL PBS. One part of the sperm suspension and two parts of TBA-trichloroacetic acid (TCA) reagent (0.375% w/v TBA, 15% w/v TCA and 0.25 N HCl) were mixed and placed in boiling water for 15 minutes. The sperm suspension was cooled and centrifuged, and the supernatant was subjected to assessment of absorbance at 535 nm. The MDA concentration was determined by the following formula:
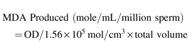
Total antioxidant capacity (ferric reducing ability of plasma assay)
After thawing, each semen sample was centrifuged (1500 × g, 10 minutes), and the supernatant was aspirated and used for the assessment of total antioxidant activity by the ferric reducing ability of plasma (FRAP) assay.
24
A working FRAP reagent solution was prepared by mixing acetate buffer: (3.1 g sodium acetate trihydrate [Merck] and 16 mL glacial acetic acid [Merck], and raised to 1 L with distilled water), 2,3,5-triphenyltetrazolium chloride (TPTZ): (0.031 g TPTZ [Sigma-Aldrich] per mL of 40 mM HCl [Merck]), and ferric chloride: ([Merck] [0.054g in 10 mL distilled water]) in ratios 10:1:1, respectively. A 3 mL sample of the working FRAP reagent was taken as the blank and 3 mL of distilled water was taken as the negative control. A standard was prepared by adding 100 μL of ascorbic acid (1 mM; Merch, Germany) to 3 mL FRAP reagent. Seminal plasma (100 μL) of a control and each experimental extender were mixed with 3000 μL FRAP reagent and the absorbance of the blank, standard, and sample was taken by a spectrophotometer at 593 nm. The FRAP value for semen sample was calculated by the following formula:
Statistical analysis
Results are described as mean ± standard error of mean. The effects of different experimental extenders on sperm progressive motility, plasma membrane integrity, viability, livability, DNA integrity, sperm malondialdehyde, and total antioxidant capacity were analyzed by analysis of variance by using IBM SPSS Statistics (Version 23) software at a 5% level of significance. As a post hoc test, Duncan's test was applied.
Results
Sperm motility (%)
According to the results, post-thaw sperm motility (Fig. 1) was higher (p < 0.05) in extenders E1 (80.00 ± 1.58), E2 (84.00 ± 1.87), E3 (86.00 ± 1.87), E4 (86.00 ± 1.87), and E5 (82.00 ± 2.55) compared to E6 (59.00 ± 2.92), E7 (33.00 ± 2.00), and control (65.00 ± 2.24) extenders.
Effect of different concentrations of CPLL solution and glycerol in extender on motility of Nili-Ravi buffalo bull spermatozoa. Different letters (a–c) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol. CPLL, carboxylated poly-
Sperm plasma membrane integrity (%)
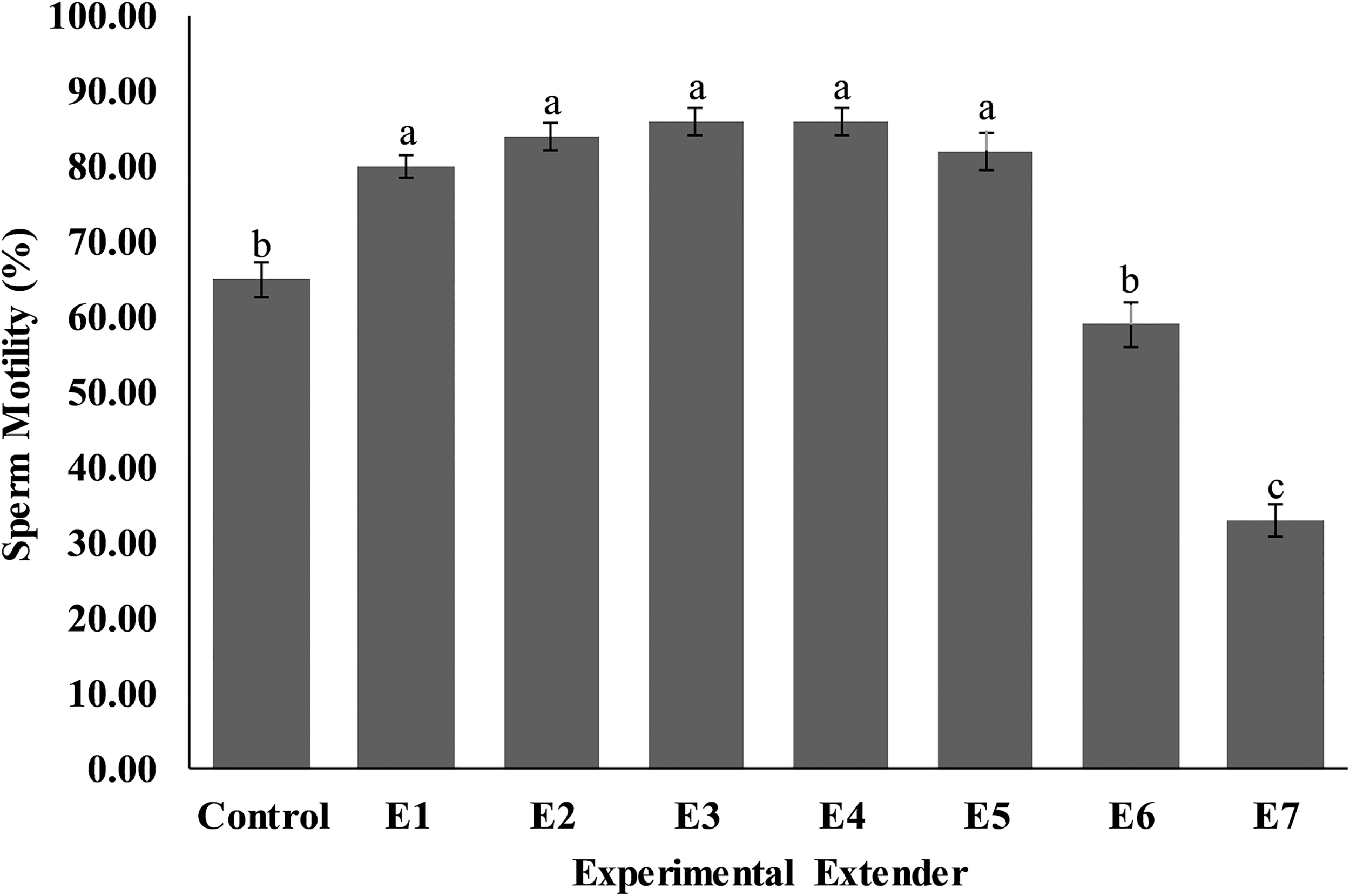
Post-thaw sperm plasma membrane integrity (Fig. 2) was significantly higher (p < 0.05) in extenders E1 (47.00 ± 2.21), E2 (47.20 ± 1.46), E3 (53.00 ± 1.58), E4 (47.00 ± 1.30), and E5 (46.40 ± 3.40) compared to E6 (40.60 ± 1.63), E7 (28.20 ± 2.20), and control (39.80 ± 0.86) extenders.
Effect of different concentrations of CPLL solution and glycerol in extender on plasma membrane integrity of Nili-Ravi buffalo spermatozoa. Different letters (a–d) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.
Sperm viability and livability (%)
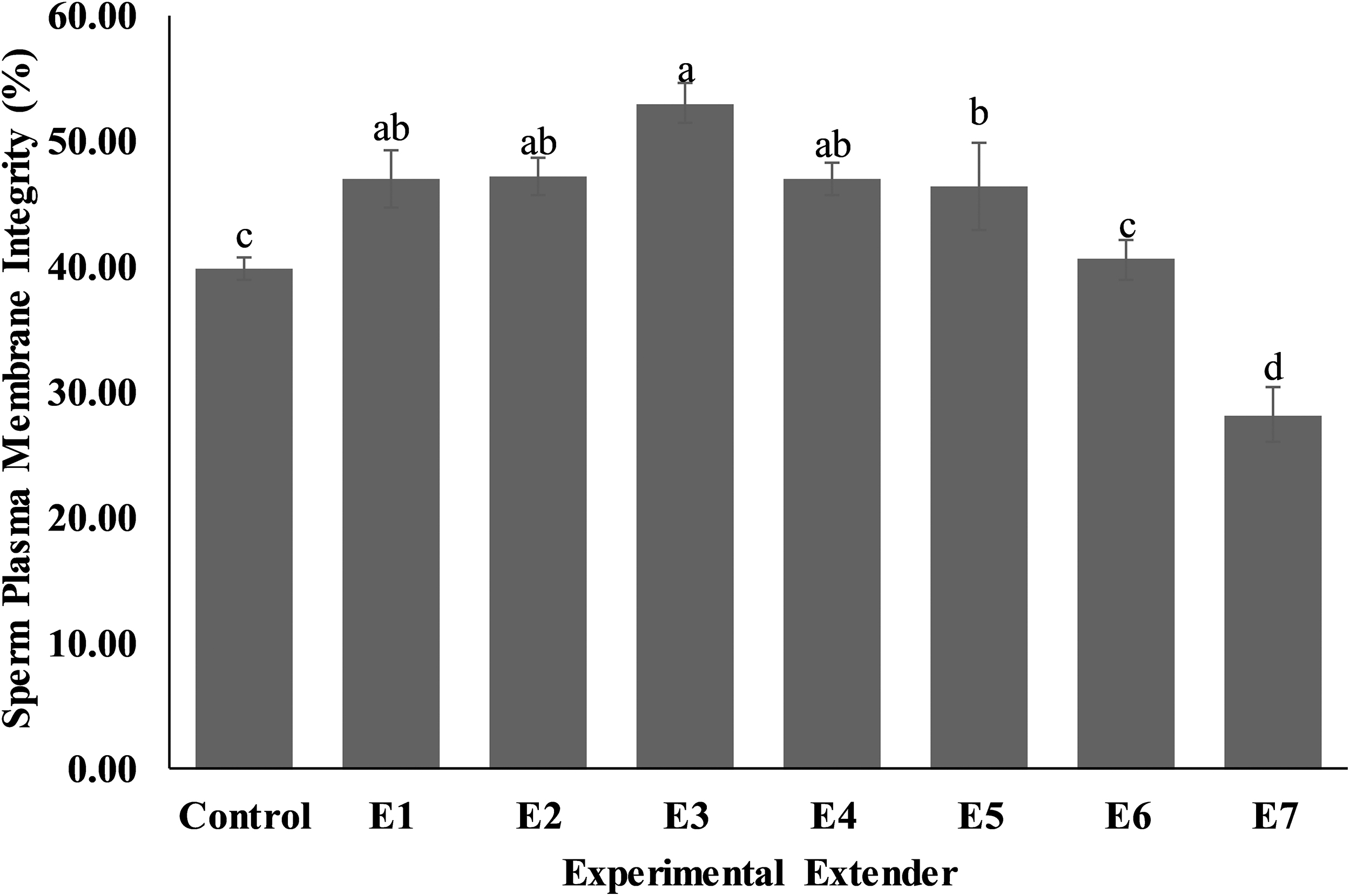
Sperm viability (Fig. 3) was recorded as higher (p < 0.05) in extender E4 (71.00 ± 2.53) compared to control (48.40 ± 0.68), E1 (52.60 ± 2.40), E2 (61.80 ± 0.73), E3 (62.00 ± 1.00), E5 (58.40 ± 2.01), E6 (46.80 ± 2.08), and E7 (34.80 ± 3.29) extenders. Similar results were observed for sperm livability (Fig. 4) in which extender E4 showed good results (p < 0.05) in comparison to other extenders viz., control (60.60 ± 1.89), E1 (58.40 ± 2.77), E2 (67.60 ± 1.29), E3 (68.20 ± 1.66), E5 (63.40 ± 1.94), E6 (58.00 ± 3.27), and E7 (49.60 ± 0.75).
Effect of different concentrations of CPLL solution and glycerol in extender on viability of Nili-Ravi buffalo spermatozoa. Different letters (a–e) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.

Effect of different concentrations of CPLL solution and glycerol in extender on livability of Nili-Ravi buffalo spermatozoa. Different letters (a–d) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.
Sperm acrosomal integrity (%)
The highest (p < 0.05) sperm acrosomal integrity (Fig. 5) was observed in extender E1 (88.40 ± 0.81), E2 (87.20 ± 1.66), E3 (88.20 ± 1.39), E4 (93.00 ± 0.89), and E5 (91.60 ± 1.57) compared to control (75.20 ± 5.08), E6 (76.00 ± 1.58), and E7 (62.00 ± 7.53) extenders.
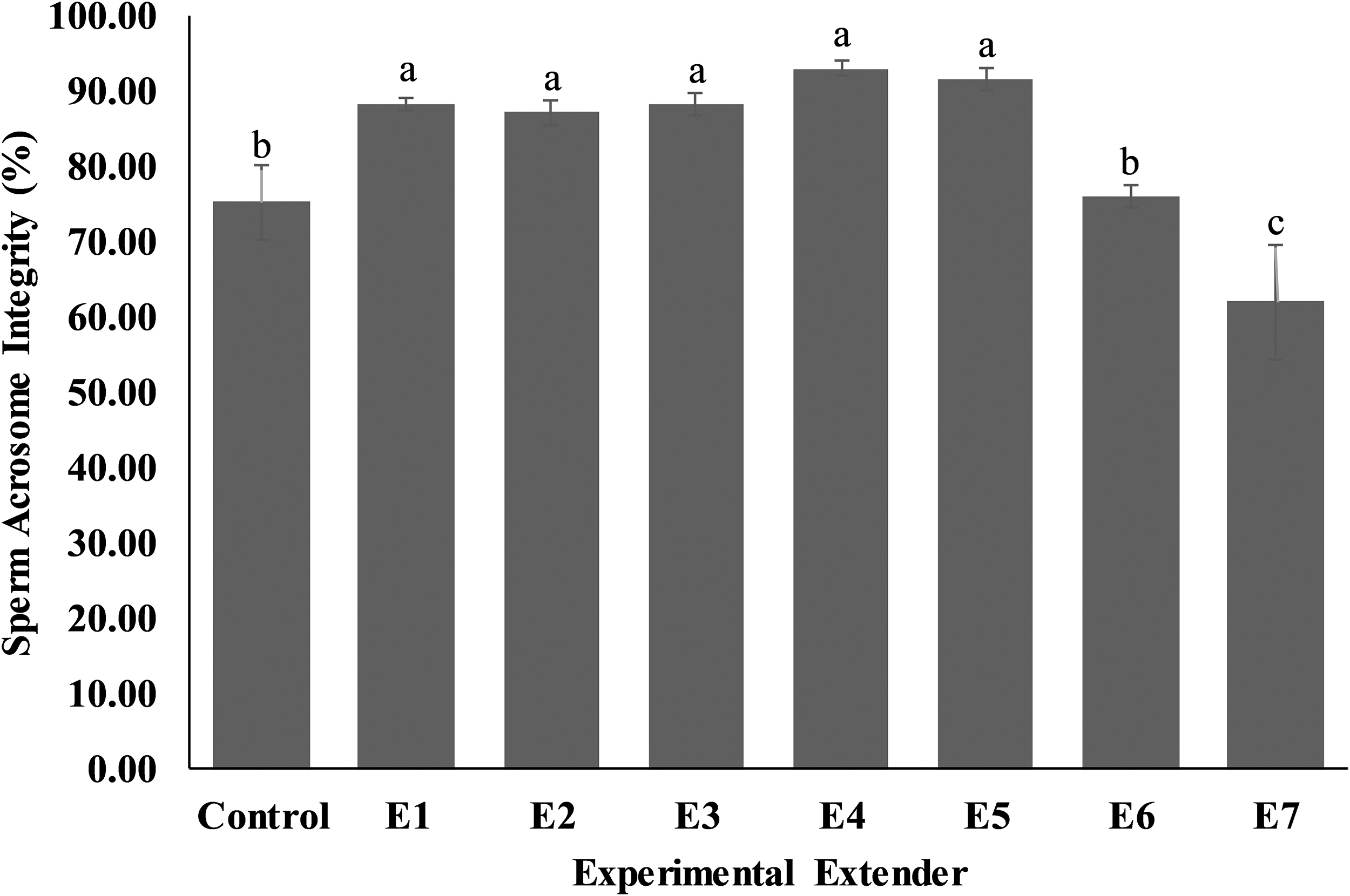
Effect of different concentrations of CPLL solution and glycerol in extender on acrosome integrity of Nili-Ravi buffalo bull spermatozoa. Different letters (a–c) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.
Sperm DNA integrity (%)
Sperm DNA integrity (Fig. 6) was higher (p < 0.05) in E2 (95.00 ± 1.26), E3 (95.00 ± 0.89), E4 (95.00 ± 0.55), and E5 (94.60 ± 0.60) compared to control (90.60 ± 0.81), E1 (90.80 ± 0.73), E6 (91.60 ± 0.81), and E7 (90.80 ± 0.80) extenders.

Effect of different concentrations of CPLL solution and glycerol in extender on sperm DNA integrity of Nili-Ravi buffalo bull. Different letters (a, b) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.
Lipid peroxidation (mol/mL/million sperm)
Sperm MDA levels (Fig. 7) were lower (p < 0.05) in E3 (423.07 ± 2.33), E4 (419.33 ± 2.33), E5 (410.59 ± 3.64), and E6 (401.85 ± 5.07) compared to control (766.27 ± 5.44), E1 (757.54 ± 4.67), E2 (767.52 ± 2.33), and E7 (778.75 ± 2.50) extenders.
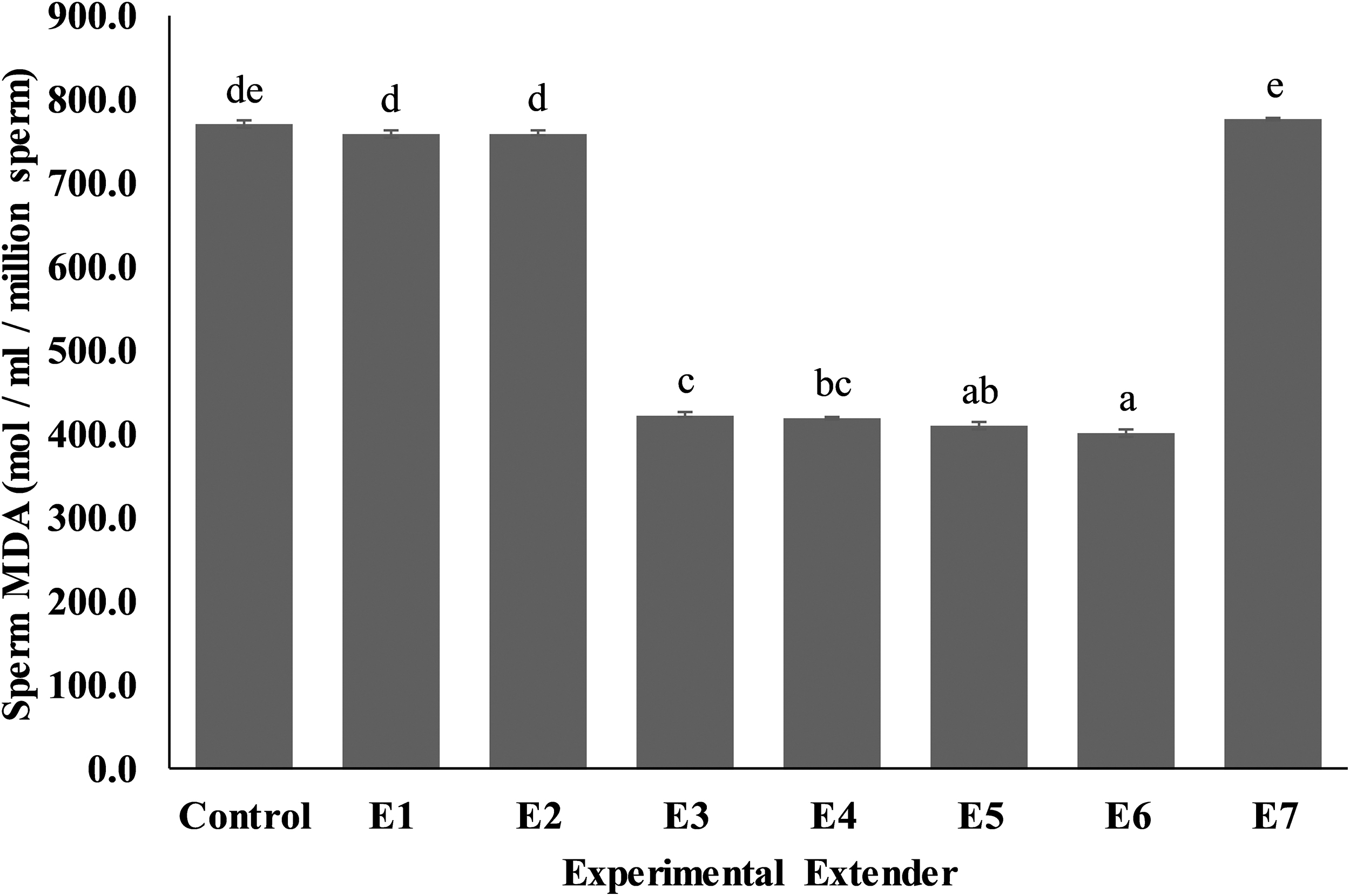
Effect of different concentrations of CPLL solution and glycerol in extender on MDA levels of Nili-Ravi buffalo bull spermatozoa. Different letters (a–e) on each bar show significant difference (p < 0.05) among treatments. Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol. MDA, malondialdehyde.
Total antioxidant capacity of seminal plasma (μM)
Total antioxidant capacity of seminal plasma (Fig. 8) was higher (p < 0.05) in extender E5 (29.28 ± 1.01), E6 (27.92 ± 1.96), and E7 (28.43 ± 1.61) compared to control (20.56 ± 2.09), E1 (16.19 ± 1.32) E2 (17.02 ± 1.25), E3 (14.55 ± 1.42), and E4 (19.16 ± 0.68) extenders.
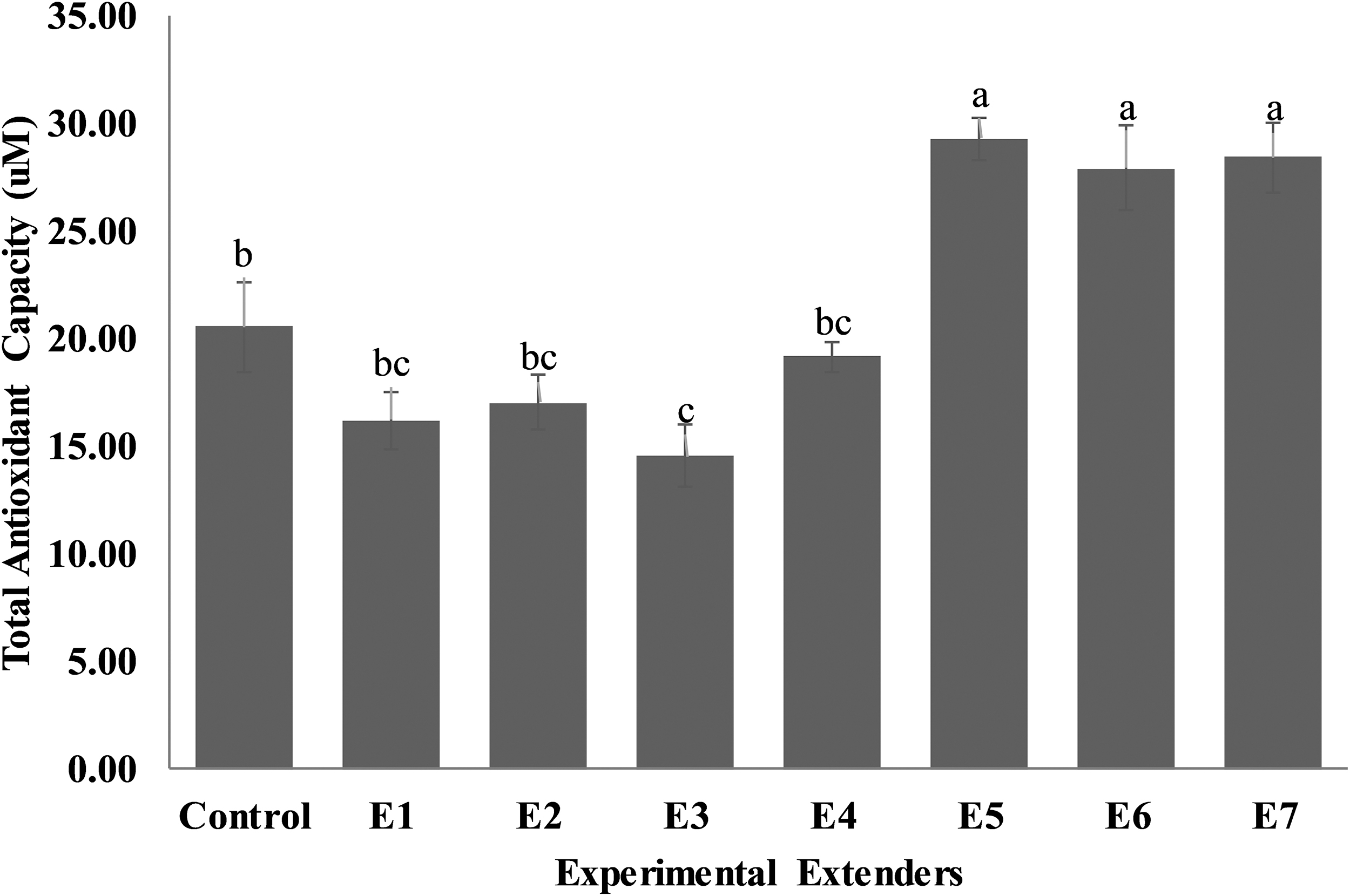
Effect of different concentrations of CPLL solution and glycerol in extender on total antioxidant capacity of Nili-Ravi buffalo bull of seminal plasma. Different letters (a–c) on each bar show significant difference (p < 0.05) among treatments. *Control: 0% CPLL +7% Glycerol E1: 1% CPLL +6% Glycerol. E2: 2% CPLL +5% Glycerol E3: 3% CPLL +4% Glycerol. E4: 4% CPLL +3% Glycerol E5: 5% CPLL +2% Glycerol. E6: 6% CPLL +1% Glycerol E7: 7% CPLL +0% Glycerol.
Discussion
Since the discovery of glycerol, sperm cryopreservation has become a reality with numerous advancements, achieving optimum conception rates (69.6%–78.78%) using AI in cattle. 25 It is a matter of concern that AI is not very successful in buffalo as it is in cattle. The factors contributing to this issue are silent heats on the female side and poor quality of cryopreserved sperm from male side. Therefore, cryobiologists are continuously investigating the buffers, chemically defined extenders,26,27 antibacterials,28–30 antioxidants, 18 and AFPs31–33 for buffalo sperm preservation. Studies are also being conducted to investigate nonpermeating and permeating cryoprotectants for their replacement with better agents in terms of improved freeze-thaw quality and reduced oxidative stress.9–11,34,35
Glycerol has been used traditionally as a permeating cryoprotectant for >70 years, inevitably associated with higher cytotoxicity and less cryoprotectant ability compared to modern day cryoprotectants like CPLL. 12 CPLL has low toxicity compared to other cryoprotectants. It is also known to cause noncolligative freezing point depression even at lower concentrations, induce freeze tolerance, and inhibit recrystallization by binding to ice during freeze-thaw process.36,37 Furthermore, CPLL can protect sperm cells from hypothermic and freezing damage by protecting the membrane, as it has the ability to bind cell membranes, 38 a characteristic that make it a suitable candidate for replacement of glycerol as a freezing injury inhibitor. 39 Previous studies have reported its use as a cryoprotectant for cell12–14 and oocyte 15 and sperm. 16 A recent study on Nili-Ravi buffalo sperm investigated two levels of glycerol (7% and 5%) with 0.25%–1% levels of CPLL for cryopreservation of Nili Ravi buffalo sperm. 17 This study is a step further to find some positive synergism between glycerol and CPLL at some lower level of glycerol, with a corresponding increased level of CPLL for buffalo bull sperm cryopreservation.
The sperm plasma membrane is a prime site of damage during cryopreservation, which results in reduced livability and viability and ultimately the fertilizing ability of sperm. Although supra vital stains were used in assessing the viability of buffalo sperm, it has been reported that viability (live sperm with intact acrosome) assessed through a dual staining procedure using Trypan blue and Giemsa stain is highly related with fertility.40–43 CPLL has (have) low permeability through the membrane and is (are) reported to interact and protect the cell membrane. In this study, when glycerol was gradually reduced with a concurrent increase in CPLL, sperm plasma membrane integrity, viability, and livability were improved up to the addition of 4% CPLL replacing 4% glycerol. Combinations of 0% to 1% glycerol with 7% and 6% CPLL did not show any beneficial effect on the aforementioned parameters. It is suggested that CPLL is a nonpermeating cryoprotectant and might not have been dehydrated adequately for cryopreservation, 16 while, glycerol, being a permeating cryoprotectant, might have induced rapid dehydration to protect cells from freezing. It has previously been confirmed that partial replacement of glycerol with CPLL protects the bovine sperm through reducing the membrane damage. 16 A recent study reported improvement in post-thaw quality and fertility of buffalo sperm with 0.75% CPLL and 5% glycerol. 17 It is relevant to mention that CPLL possesses AFP-like properties that might have added protection to the sperm cells as has been reported using AFPs for buffalo sperm.31–33
Sperm DNA integrity is crucial for successful fertilization and early embryonic development. Sperm with apparent normal post-thaw quality can fertilize oocytes; however, the subsequent embryos may show impaired development in the case of DNA damage initiation. It is also relevant to mention that reactive oxygen species resulting from dead and damaged cells during cryopreservation are the major stressors leading to DNA damage. Antioxidants are therefore added to the extender to reduce oxidative stress and improve cryosurvivability and DNA integrity of sperm.18,34 Whether CPLL possesses any antioxidant activity or not has not been reported yet. However, we investigated the lipid peroxidation level and antioxidant activity of buffalo sperm cryopreserved with different levels of CPLL replacing glycerol. Interestingly, the lipid peroxidation level was reduced, concomitant with increase in antioxidant activity and DNA integrity, as the CPLL was increasingly replaced with the glycerol. These findings are suggestive of some antioxidative effect of CPLL or reduction in oxidative stress or cytotoxicity associated with glycerol, which might have decreased the global oxidative stress.
Conclusion
It is concluded that synergism between CPLL and glycerol (4% CPPL and 3% Glycerol) seems to improve the freezability of Nili-Ravi buffalo semen by reducing oxidative stress.
Footnotes
Authors' Contributions
S.A. and M.A.A. planned/executed the study and drafted the article. M.A.A., J.A., and S.I. were involved in practical work. B.A.R. and M.S.A. critically proof read the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The study was funded by Higher Education Commission, Islamabad, Pakistan, under National Research Programme for Universities by project No. 20-4003/NRPU/R&D/HEC/14.