
Abstract
Modern taxonomy requires the preservation of biospecimens for both morphological and molecular applications. The utility of a previously identified preservative, dimethyldimethylhydantoin hydantoin (Dekafald®), to retain both physical diagnostic traits and the DNA integrity of biological specimens remains unknown. Using 439 eggs and 414 larvae from two North American fish species, we compared three hydantoin solutions at different concentrations (5%, 10%, and 20%) with gold standard preservatives (10% buffered formalin, 95% ethanol) to evaluate morphological trait retention up to 90 days, and DNA barcoding success up to 56 days. While the 5% hydantoin solution had the most sequencing success by 56 days, the 10% hydantoin solution was the best multipurpose preservative. Future work should assess the performance of ∼10% hydantoin solution over longer time periods, and its applicability to other taxa such as Arthropoda.
Introduction
Fluid preservation has been utilized since the 17th century to prevent the deterioration of relevant biological specimens. 1 Specimen preservation is critical for maintaining the integrity of taxonomically relevant morphological traits over long periods of time. With continuous advancements in molecular technologies, there is a modern need to retain specimens beyond their use for standard morphology. Increasingly, both physical traits and genetic material are required from preserved specimens for an integrative approach to taxonomy. 2 For example, fishery managers and regulatory agencies seek to characterize early life stage fishes for biomonitoring. 3 However, the small size and few distinct features of fish eggs and larvae present a challenge to both traditional and genetic methods of species identification, such as DNA barcoding. 4
Retaining the biological characteristics and contents of an aquatic specimen for both morphology and DNA analyses requires careful consideration of the selected preservative. Traditionally, morphological methods utilize either formalin (a solution of formaldehyde and water) or diluted ethanol (EtOH). Formalin rapidly and cost-effectively fixes specimens, which stops cellular activity while maintaining important morphological traits such as color, pigmentation, and pliability. 5 Formalin-preserved specimens can last hundreds of years; however, the solution quickly degrades DNA within days to months, 6 and formaldehyde is a known carcinogen and combustible liquid.
Conversely, EtOH is less effective for morphology. It has poorer trait retention (such as tissue shrinkage and color loss), is slower to arrest cellular activity, is expensive at the higher purities required for preservation, has a high evaporation rate, and is highly flammable. 7 Nonetheless, EtOH solutions can maintain high-quality DNA and osteological traits for several years, 7 and has a much lower associated toxicity than formalin. The apparent trade-offs between these solutions present a challenge to fishery researchers seeking a single, versatile preservative.
Dimethyldimethylhydantoin (DMDM hydantoin) is a formaldehyde-releasing organic compound found in personal care products such as soaps and shampoos under the trade names Glydant® and Dekafald®. Dam 8 initially identified DMDM hydantoin as a potential alternative for biological specimen preservation. Molin and Dam 9 further proposed that 5% DMDM hydantoin solution can preserve embryonic mouse tissue-derived proteins, mRNA, and enzyme activity comparable with 70% ETOH for up to 3 weeks.
Carter 10 suggested that both the lower toxicity of DMDM hydantoin compared with formalin and their similar chemical degradation signatures of pork tissue-based proteins after 2 months present the compound as a suitable alternative to formalin. Nevertheless, the effectiveness of DMDM hydantoin as a preservative of both organism morphology and DNA integrity for molecular identification remains unexamined.
The objective of this study was to assess the relative effectiveness of DMDM hydantoin (hereafter referred to as “hydantoin”) solutions for maintaining the morphological and genetic integrity of economically important fish eggs and larvae, compared with traditional formalin and EtOH treatments.
Materials and Methods
Specimen collection and fluid preservation
All eggs (E; contained eyed embryos) and larvae (L) from both fathead minnow (Pimephales promelas, PP) and Atlantic silverside (Menidia menidia, MM) were reared for research purposes and obtained from Aquatic Research Organisms (Hampton, NH). Animal experimentation complied with the American Fisheries Society Guidelines for the use of fishes in research. 11 Individual specimens were allocated to either morphological analysis (PP: n = 247, E = 126, L = 121; MM: n = 216, E = 108, L = 108) or DNA barcoding (PP: n = 218, E = 108, L = 110; MM: n = 172, E = 97, L = 75).
Sample sizes for both analyses were based on feasibility within the time frame of the study. Three preservative solutions were tested: 10% buffered formalin; three concentrates of hydantoin (5%, 10%, and 20% [from Dekafald solution, 55% DMDM hydantoin]); and 95% EtOH. All dilutions were made with distilled water.
Morphology
Specimens were morphologically assessed by three assessors on day(s) 1, 2, 7, 14, 28, 56, and 90 postpreservation. Day 0 specimens served as controls. A matrix of the experimental design, including sample sizes per day and species, is presented in Supplementary Table S1. An AmScope 3.5X-90X Trinocular Inspection Zoom Stereomicroscope and AmScope MU800 camera were used to capture specimen images at a known scale, which were imported into ToupView v3.7.6701. Specimen clarity and structural retention were visually assessed in real time on a qualitative basis relative to day 0 controls. The diameter of each egg (in mm) was measured at two perpendicular points and averaged. For larval specimens, head length, head depth, and total length were measured in mm (as defined by Refs.12,13).
DNA barcoding
Five specimens of each species and life stage were extracted from each preservative on days 7, 14, 28, and 56 postpreservation. Supplementary Table S1 outlines the experimental design, and any deviations from this general format. DNA extractions were conducted using the Qiagen (Hilden, Germany) DNeasy Blood and Tissue Kit, according to the manufacturer's protocol for animal tissues. Average total DNA concentration and 260/280 ratios were estimated for each extract with a NanoDrop™ ND8000 spectrophotometer (Supplementary Table S2).
For each genomic DNA sample, the mitochondrial cytochrome c oxidase I (COI-5P) DNA barcode region for fishes was amplified using previously published protocols. 1 Briefly, all polymerase chain reactions (PCRs) had a total volume of 12.5 μL and included the following: 6.25 μL of 10% trehalose, 2.00 μL of ultrapure water, 1.25 μL 10 × PCR buffer [10 mm KCl, 10 mm (NH4)2SO4, 20 mm Tris-HCl (pH 8.8), 2 mm MgSO4, and 0.1% Triton X-100], 0.625 μL MgCl2 (50 mm), 0.125 mL of each primer cocktail (0.01 mm), 0.0625 μL of each dNTP (10 mm), 0.0625 μL of Taq DNA polymerase (New England Biolabs), and 2.0 μL of DNA template. The thermocycler reaction profile was 94°C for 2 minutes, followed by 40 cycles of 94°C for 30 seconds, 52°C for 40 seconds, and 72°C for 1 minute, with a final extension at 72°C for 10 minutes.
PCR products were bidirectionally sequenced following the methods given in Ref. 15 Sequences and trace files were deposited in the Barcode of Life Database (BOLD) in the container project DMDMH.
Statistical analyses
Two data sets were created for morphology and genetic data, respectively (Supplementary Table S2). The morphological data set recorded (in mm) total body length, head length, and head depth for larvae, as well as mean egg diameter for eggs. The value “NA” was assigned to trait(s) that did not apply to a particular life stage. For DNA barcoding data, sequencing success was assigned a binary classification, with “Yes” indicating either a uni- or bidirectional sequence obtained with ≥98% species match to the target species using BOLD 16 and “No” was assigned to specimens that failed to sequence in any direction, or failed to obtain a BOLD ID match of ≥98% to the target species.
All statistical analyses were performed in R version 3.5.3. 17 Using two-way type III analysis of variances (ANOVAs) for unbalanced designs, 18 we evaluated the effects of both preservation and exposure time on each egg diameter, total body length, head length, and head diameter. Post hoc testing was performed using Dunnett's test, which compared each preservative to controls. 19 Standardized mean differences in trait measurements (effect size) were estimated relative to controls using Hedge's g 20 and its corresponding positive bias correction factor. 21
Results
Visual assessment
Specimens were qualitatively assessed for structural retention and visual clarity, relative to day 0 controls. Eggs preserved in 95% EtOH reacted poorly to treatment, turning opaque and yellow; however, larval specimens were preserved with the highest degree of clarity and structural integrity relative to the other preservatives. Formalin-treated eggs and larvae remained well preserved, although eggs exhibited a reduction in shape consistency, and embryo yolk sacs became opaque over time.
Eggs preserved in 5% hydantoin appeared to lose their spherical shape and clarity over time, and larval specimens exhibit the greatest degree of structural degradation and morphological ambiguity relative to all other treatments. However, eggs and larvae preserved in 10% and 20% hydantoin were found to be well preserved morphologically, and did not change meaningfully after 90 days of exposure. Figures 1 and 2 illustrate physical changes in fathead minnow egg and larvae specimens, respectively, from each treatment over time. Of the original 247 fathead minnow specimens photographed, 8 eggs and 2 larvae were excluded from analyses due to structural changes that made measurements impossible to obtain.
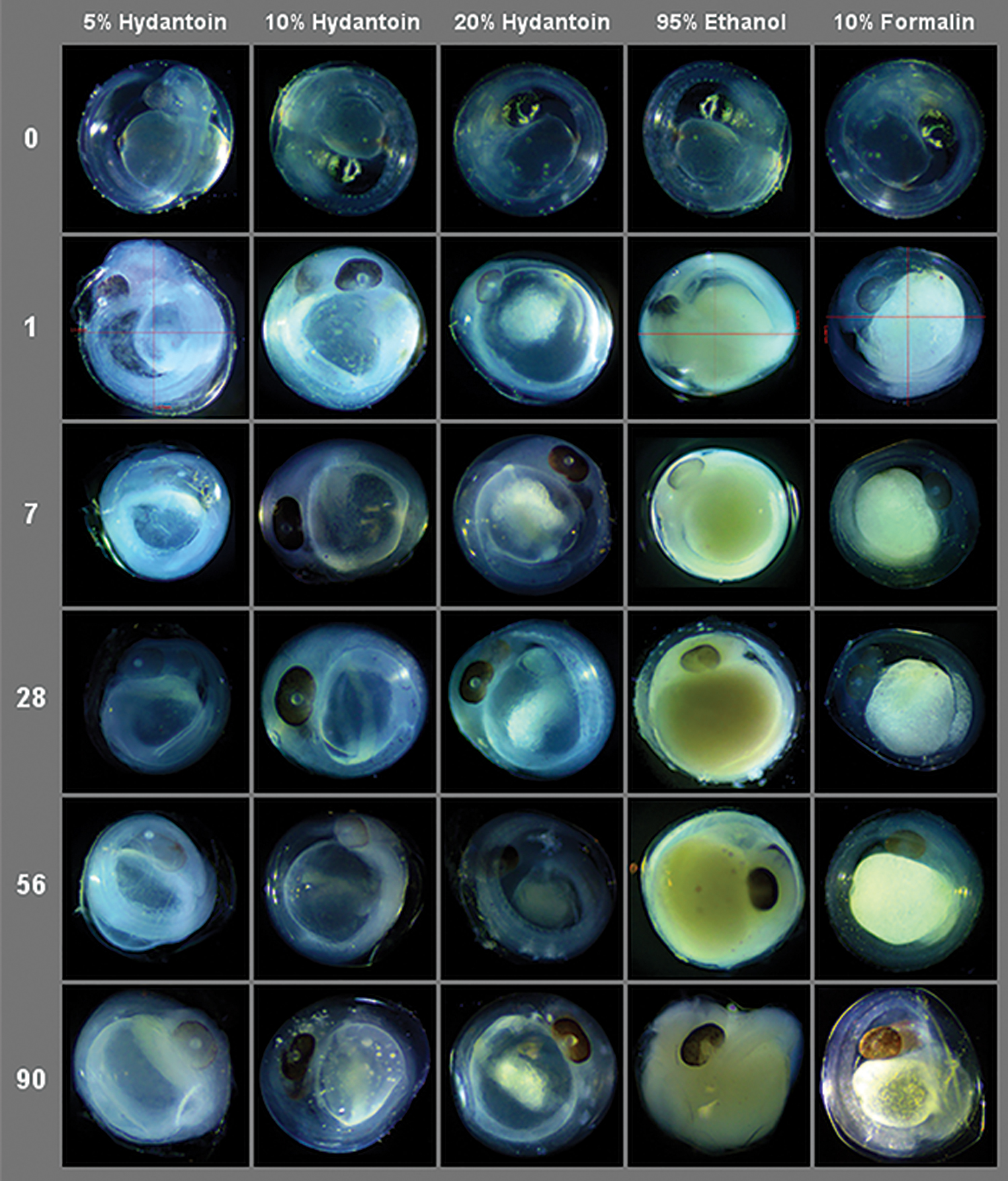
Representative images of fathead minnow (Pimephales promelas) eggs from each experimental preservative treatment across exposure times. Color images are available online.
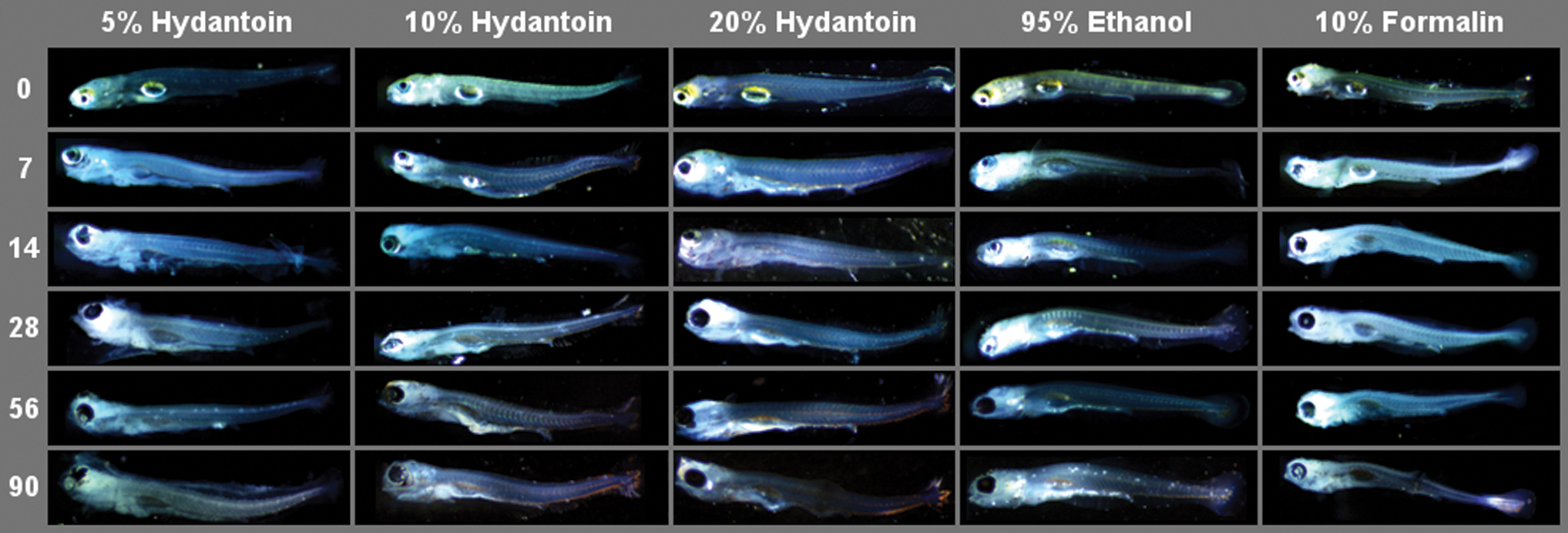
Representative images of fathead minnow (P. promelas) larvae from each experimental preservative treatment across exposure times. Color images are available online.
Effects of preservative and exposure time on mean trait length
We evaluated the effects of preservative and exposure time on egg diameter, total body length, head length, and depth using type III ANOVA. Preservative significantly influenced (p ≤ 0.05) all traits across both species, except for fathead minnow head length (F(5,112) = 0.94, p = 0.46). Time significantly influenced the larval body lengths of both species (MM: F(5,101) = 8.76, p = 6.29e-7; PP: F(5,112) = 5.09, p = 3e-4), as well as the head depth of Atlantic silversides (F(5,101) = 19.97, p = 8.46e-14) and the egg diameter of fathead minnows (F(5,111) = 3.33, p = 8e-3), respectively (Table 1).
Type III Analysis of Variance Statistics, with Various Morphological Traits as the Response Variable, and Time (Day) and Preservation Treatment as the Explanatory Variables
Significant results are in bold.
Preser, preservation treatment.
Post hoc analysis using Dunnett's test was performed to compare the five experimental treatments (EtOH, formalin, hydantoin 5%, 10%, and 20%) to control specimens. Briefly, Atlantic silverside mean egg diameter was significantly increased by all treatments (p ≤ 0.05, Table 2). For fathead minnows, only 5% hydantoin had a positive effect (mean difference from control eggs = 0.11 mm). The larval body lengths of both species preserved in EtOH did not notably differ from controls, as well as silversides preserved in 5% hydantoin; all other treatments negatively impacted total body length (Table 2).
Dunnett's Test Statistics, Comparing the Five Preservative Treatments (Ethanol, Formalin, 5% Hydantoin, 10% Hydantoin, 20% Hydantoin) with Control Specimens, with Respect to Mean Morphological Trait Measurements
Significant results are in bold.
CI, confidence interval; C, control; E, Ethanol; F, Formalin; H5, 5% Hydantoin, H10, 10% Hydantoin; H20, 20% Hydantoin.
Average larval head length did not differ from control specimens across all treatments and species. Five percent hydantoin increased mean head depth in Atlantic silversides (mean difference = 0.09 mm, p = 0.03), whereas 20% hydantoin increased fathead minnow head depth (mean difference = 0.11 mm, p = 0.03). In addition, the standardized mean differences for each morphological trait were estimated by preservative relative to control specimens using Hedge's g, and are reported in Supplementary Table S3 for use in meta-analysis.
DNA barcoding
Across species and life stages, DNA barcode identification success varied by both preservative and exposure time (Table 3). Considering all exposure times simultaneously, total barcoding success by preservative was as follows: EtOH (92%, n = 75), 10% hydantoin (82%, n = 103), 5% hydantoin (80%, n = 75), 20% hydantoin (66%, n = 62), and formalin (32%, n = 75). Across all preservative treatments, barcoding success generally declined with increased exposure time: 7 days (79%, n = 62), 14 days (84%, n = 110), 28 days (66%, n = 108), and 56 days (60%, n = 110).
Contingency Table Summarizing DNA Barcoding Success and Failure, with Both Species and Life Stages Pooled
Preservative data are across all exposure times, and vice versa for exposure time data.
Yes refers to DNA barcoding success and No to DNA barcoding failure.
Barcoding success also varied by species and life stage (Table 4). For fathead minnows preserved in hydantoin, sequencing success was typically greater for eggs compared with larvae, but the opposite was observed in Atlantic silversides (Table 4). A greater proportion of fathead minnow eggs (100%) and larvae (60%) preserved in 5% hydantoin for 56 days yielded a DNA barcode identification compared with the EtOH treatment (E = 20%, L = 60%). Five percent hydantoin Atlantic silversides (E = 80%, L = 100%) also sequenced comparably with EtOH-treated silversides at 56 days (E = 100%, L = 100%).
Contingency Table of DNA Barcoding Success (%), Organized by Preservative and Exposure Time for Both Species
“E” refers to egg and “L” is larval life stage. “NA” indicates no specimens existed for a particular category.
Ten percent hydantoin was the next most successful preservative after 5%. At 56 days postpreservation, 100% of Atlantic silverside larvae and 40% of eggs obtained a species identification, and 50% of fathead minnow larvae and 80% of eggs were successfully sequenced. One hundred percent of larvae from both species preserved in 20% hydantoin for 56 days were barcoded successfully; however, eggs were not as successful at this concentration (MM = 20%; PP = 40%).
Discussion
We explored three different concentrations of DMDM hydantoin for their ability to preserve four morphological characters and retain genetic material suitable for species identification via DNA barcoding. With respect to morphology, the 10% hydantoin solution emerged as the best candidate. Relative to the classic preservative formalin, 10% hydantoin eggs had the greatest visual clarity up to 90 days of exposure (Fig. 1). While all hydantoin treatments (as well as formalin) significantly reduced mean total larval body length in either one or both species, 10% hydantoin was the only solution of the three that did not significantly alter either dimension of the larval head capsule in any species relative to control specimens (Table 2).
The similar trait changes induced by formalin and hydantoin are potentially linked to formaldehyde, which is present in both solutions. Formaldehyde chemically reacts with amine groups to induce protein denaturation from crosslinking reactions. 10 The gradual release of small amounts of formaldehyde by hydantoin is an advantage for long-term preservation, 10 but the chemical's total useful lifetime for this purpose is beyond the scope of the present study.
Of all five preservatives, EtOH had the least instances of significant impact on mean trait lengths in post hoc testing (Table 2). EtOH-preserved larvae in particular were qualitatively reported to be preserved with the highest degree of clarity and structural integrity (Fig. 2), reflected in the results of post hoc testing for total body length (Table 2). Indeed, EtOH-based solutions have been demonstrated to preserve the osteological integrity of larval specimens better than formaldehyde-based solutions.7,22
However, EtOH-treated eggs experienced detrimental yolk sac color and overall shape changes that 10% hydantoin did not demonstrate over time (Fig. 1). The detrimental color changes observed in EtOH-preserved eggs could be related to both pH and oxidation. The pH of high-proof EtOH solutions (>7.0) can create translucent specimens via the dissolution of pigments. 22 The presence of water and oxygen in EtOH solutions can also cause melanin pigment loss. 23
Across species and life stages, DNA barcoding was generally successful for each preservative treatment (≥66% of specimens attained a species identification), with the exception of formalin (32%, Table 3). The overall lack of successful sequencing from formalin-treated specimens is unsurprising, given formalin's tendency to fragment DNA over time 24 ; however, it is worth noting that all 56-day formalin-treated fathead larvae were successfully barcoded (Table 4). While 5% hydantoin performed closer to the standard genetic preservative EtOH at longer exposure times—particularly for eggs—the ability of 10% hydantoin to better retain morphological characteristics suggests it is the preferred all-round preservative of fish eggs and larvae.
Molin and Dam 9 initially identified hydantoin's ability to preserve RNA and tissue proteins in mice. However, hydantoin's usage in biological preservation appears to predate this; Steedman 25 proposed dimethyl hydantoin formaldehyde (DMHF) as a resin for mounting small arthropods, such as crustaceans and insects. Select groups (see 26 for a typical example) utilize DMHF to mount beetle genitalia for taxonomic purposes. With respect to genetic applications, Jeffery 27 tested a hydantoin solution of unknown concentration for flow cytometry for the purpose of arthropod genome size estimation, but reported “unreliable” results.
To our knowledge, this study represents the first attempt to obtain species identifications using DNA barcode sequences from specimens preserved in hydantoin solutions. Given the good success of 10% hydantoin solution in both genetic and morphological contexts, future work should investigate the use of this preservative beyond 90 days and on taxa from other orders, such as Arthropoda. Hydantoin presents other advantages to the user, being of lower toxicity than formalin and cheaper and easier to obtain than EtOH. In conclusion, with additional, longer duration storage testing, we are cautiously optimistic that an ∼10% solution of hydantoin could be an acceptable, multipurpose preservation fluid that supports both the maintenance of morphological characters and the molecular integrity of fish eggs and larvae for fishery science.
Footnotes
Acknowledgment
The authors appreciate the applied research support and financial backing of the Electric Power Research Institute's Fish Protection Program.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Electric Power Research Institute, Fish Protection Program.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.