
Abstract
Recently, many studies have found that late embryogenesis abundant (LEA) proteins could protect cells from drought, high salinity, and other stress conditions. Because LEA proteins maintain the integrity and stability of cell membranes, LEA proteins increase the cell's tolerance to dehydration stress, and reduce the osmotic and freezing damage during freezing. Whether LEA proteins could reduce cryopreservation damage and improve cell viability remains to be confirmed. In this study, we purified the recombinant AavLEA1 proteins, examined their thermal solubility and the effect of AavLEA1 proteins on the osmotic stress of cells, and studied the effects of the AavLEA1 protein on cryopreservation of human umbilical cord matrix mesenchymal stem cells (hUCM-MSCs). We utilized three concentrations of AavLEA1 protein (0.1, 0.5, and 2 mg/mL) to cryopreserve hUCM-MSCs and analyzed cell viability and apoptosis of MSCs after slow-cooling cryopreservation. We also examined the cryopreservation effect of AavLEA1 protein on hUCM-MSCs survival with 0%, 2%, 5%, and 10% (v/v) dimethyl sulfoxide (DMSO). We found that the survival rate of hUCM-MSCs supplemented with AavLEA1 protein was significantly higher than that of MSCs cryopreserved with low concentration of DMSO solution, and the apoptosis and necrosis rates were correspondingly reduced. In conclusion, recombinant AavLEA1 protein can improve the efficiency of MSC cryopreservation, increase the hUCM-MSCs viability, and partly replace DMSO during cryopreservation.
Introduction
Owing to their self-renewal and differentiation characteristics, stem cells can effectively cure many human diseases (such as diabetes, leukemia, heart disease, organ failure, and other serious diseases).1,2 With the rapid development of regenerative medicine and precision medicine, the source of stem cells has become a clinical problem to be solved. To meet the needs of clinical and experimental research, mesenchymal stem cell (MSC) banking is a crucial issue. 3 The stem cell bank provides a stable resource for regenerative medicine and precision medicine, and solves the problem of insufficient stem cell sources in the clinic. 4
Cryopreservation is well known as an efficient technique for preserving tissues and cells, including adult stem cells. However, osmotic damage and intracellular ice crystal formation are the main causes of damage during tissue and cell cryopreservation.5–7 The addition of a cryoprotective agent (CPA) reduces freezing injury during cryopreservation. However, permeable CPAs such as dimethyl sulfoxide (DMSO), ethylene glycol, propylene glycol, acetamide, glycerol, and so on, generally exhibit cytotoxic effects on cell proliferation and development in a dose-dependent manner. Nontoxic and effective CPAs are essential to minimize the cryogenic damage and eliminate cytotoxicity.
Late embryogenesis abundant (LEA) proteins produced by nonaquatic plants and lower animals are mainly a family of proteins that are hydrophilic and inherently disordered, which make them resistant to dry environments.8–10 LEA proteins are highly hydrophilic polypeptides composed of polar amino acids. In cold and dry environments, LEA proteins could maintain enzyme activity and prevent protein aggregation against freeze-induced damage.11–13 LEA proteins can increase the tolerance of cells to dehydration stress, thus protecting the integrity and stability of cell membranes. Many studies have found that LEA proteins protect cells against drought, high salinity, and other stress conditions.11,14–16
LEA proteins are nontoxic and effective as an antifreeze desiccant that protects proteins and cell membranes during freeze-drying. In cryopreservation, whether LEA proteins could reduce cryogenic damage and increase cell survival remains to be verified. AavLEA1 protein, reported in 2002, was expressed to prevent against desiccation in the nematode Aphelenchus avenae. 17 This protein is a member of the Group 3 LEA (G3LEA) protein family, and has four repeated motifs that are composed of a loosely conserved 11-mer amino acid sequence. AavLEA1 protein is highly hydrophilic (with a hydropathy score of −1.585), and exhibits unique properties of structure and thermodynamics. 18 In this study, we purified the recombinant AavLEA1 proteins and demonstrated the effects of supplementation with recombinant AavLEA1 proteins during human umbilical cord matrix mesenchymal stem cells (hUCM-MSCs) cryopreservation.
Materials and Methods
LEA proteins purification and heat solubility assay
The plasmid of AavLEA1-pET15b was purchased from Addgene (ID: 53093). The AavLEA1 gene was cloned into the pET15b vector (Novagen) using the NdeI/BamHI restriction site. Overexpression of recombinant AavLEA1 proteins tagged with 6 × His was induced in Escherichia coli Rosetta (DE3, Novagen) by 0.5 mM isopropyl-b-
The heat solubility assay was performed as described previously. 19 The purified proteins were heated at 92°C for 15 minutes, then cooled to room temperature. After heat treatment, proteins were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis with Coomassie Brilliant Blue staining.
hUCM-MSCs culture
hUCM-MSCs were obtained as previously described. 20 In brief, after treatment with collagenase type II, the dissociated cells were filtered through a nylon mesh to remove tissue debris. The isolated hUCM-MSCs were suspended in DMEM/F-12 with 10% (v/v) fetal bovine serum (Gibco), and cultured at 37°C and 5% CO2 in a humidified incubator. Culture medium was changed every 2 days until reaching 80%–90% confluence.
Osmotic tolerance assay
Osmotic tolerance of recombinant AavLEA1 proteins was studied as previously described. 19 hUCM-MSCs were collected and maintained in phosphate-buffered saline (PBS; 275 mOsm/L) medium on ice. After centrifugation, PBS was replaced with PBS, 2 × PBS (528 mOsm/L), 3 × PBS (782 mOsm/L), and 4 × PBS (983 mOsm/L), and cells were incubated for 4 hours on ice. The recombinant AavLEA1 proteins (final 0.5 mg/L) were added into hyperosmotic solution. After hyperosmotic exposure, the metabolic activities of treated cells were measured by CCK-8 kit (Dojindo, Japan). The metabolic activities can be quantitatively measured by the absorbance at 450 nm.
Cryopreservation of hUCM-MSCs
Cryopreservation
The attached hUCM-MSCs were harvested at 80%–90% confluence. After trypsinization, the cells were collected by centrifugation at 500 g for 5 minutes, and resuspended in culture medium for further use. For the cryopreservation studies, the cells were resuspended in 1 mL of precooled (on ice for 10 minutes) DMEM/F-12 medium with DMSO (Sigma-Aldrich) and various concentrations (0.1, 0.5, and 2 mg/mL) of LEA proteins. Then the cell suspension was loaded into vials. The cooling procedure was preprogrammed by controlled-rate freezer (CryoMed; Thermo Fisher Scientific, Inc.) as −2°C/min from 25°C to 4°C and −1°C/min from 4°C to −80°C. Finally, they were stored in a liquid nitrogen tank for 2 weeks.
Thawing
Vials containing the cryopreserved hUCM-MSCs were removed from the liquid nitrogen tank and immediately placed into a 37°C water bath to thaw. Then they were gently shaken until no ice crystals remained and the cells were centrifuged at 500 g for 5 minutes. The supernatant was removed, and the cells were resuspended in PBS.
Assessment of cell viability with fluorescence staining
After thawing, the viability of hUCM-MSCs was assessed by acridine orange/ethidium bromide (AO/EB) staining kit (KeyGen BioTECH Co., Ltd, China). The cells were resuspended in 20 μL PBS and the fluorescence staining solution (1 μL) was added into the cell suspension. After incubation for 5 minutes at room temperature, the cell fluorescence images of the cells were taken for assessing the immediate cell viability.
Flow cytometry analysis of cell necrosis and apoptosis
The cell necrosis and apoptosis were analyzed by flow cytometry using FITC-conjugated Annexin V and propidium iodide (PI; Vazyme Biotech Co., Ltd, China). The PI and Annexin V were used to stain cells that died of necrosis and apoptosis, respectively. The hUCM-MSCs of postcryopreservation were centrifuged at 500 g for 5 minutes. Then the cells were resuspended in binding buffer, and stained with FITC-conjugated Annexin V and PI in binding buffer on ice (4°C). After incubation for 15 minutes in the dark, the stained samples were checked using a flow cytometer (BD FACSVerse). The results were analyzed with FlowJo software.
Statistical analysis
All data are presented as mean ± standard deviation from at least three independent runs. One-way analysis of variance paired with a Tukey multiple comparisons test was used to compare cell viability of post-cryopreservation by various concentrations of LEA protein. The individual comparisons were performed using Student's two-tail paired t-test. A value of p < 0.05 was considered statistically significant.
Results
AavLEA1 has a total molecular mass of 16 kDa, and migrates on SDS gels at a calculated mass of 18 kDa (Fig. 1A). The recombinant proteins were purified using Ni2+-nitrilotriacetate affinity resin and HiLoad 16/60 Superdex 200. The final proteins in buffer containing 20 mM Tris-HCl, pH 8.0 and 50 mM NaCl were concentrated to 10 mg/mL for cryopreservation trials. To determine the biochemical properties of recombinant AavLEA1 proteins, we examined the heat solubility of AavLEA1 proteins. Heat treatment of the purified proteins showed AavLEA1 protein was mostly recovered in the supernatant even after heat treatment (Fig. 1B), indicating that recombinant AavLEA1 proteins maintain their heat solubility in bacterial lysate. LEA proteins are proteins in animals and plants that protect other proteins from aggregation owing to desiccation or osmotic stresses associated with low temperature. To test the osmotic tolerance of hUCM-MSCs, we utilized recombinant AavLEA1 proteins to form a hyperosmotic solution on ice, and measured the metabolic activities of hUCM-MSCs. The metabolic activities of hUCM-MSCs with AavLEA1 were significantly higher than those exposed to the hyperosmotic solution (Fig. 2). The results suggested that recombinant AavLEA1 proteins improved hUCM-MSCs tolerance against hyperosmotic stress at low temperature.
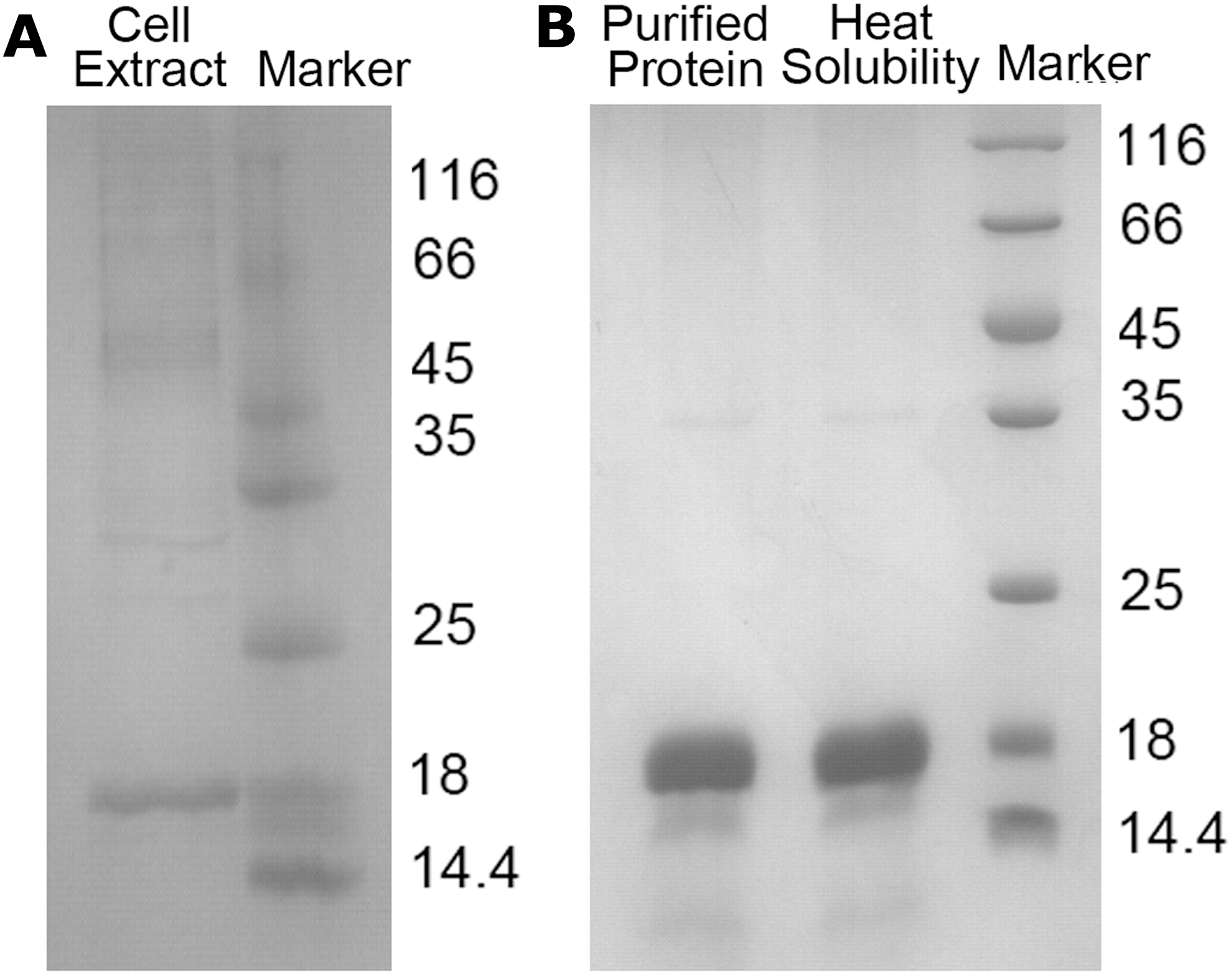
Recombinant AavLEA1 protein expression, purification, and heat soluble property.
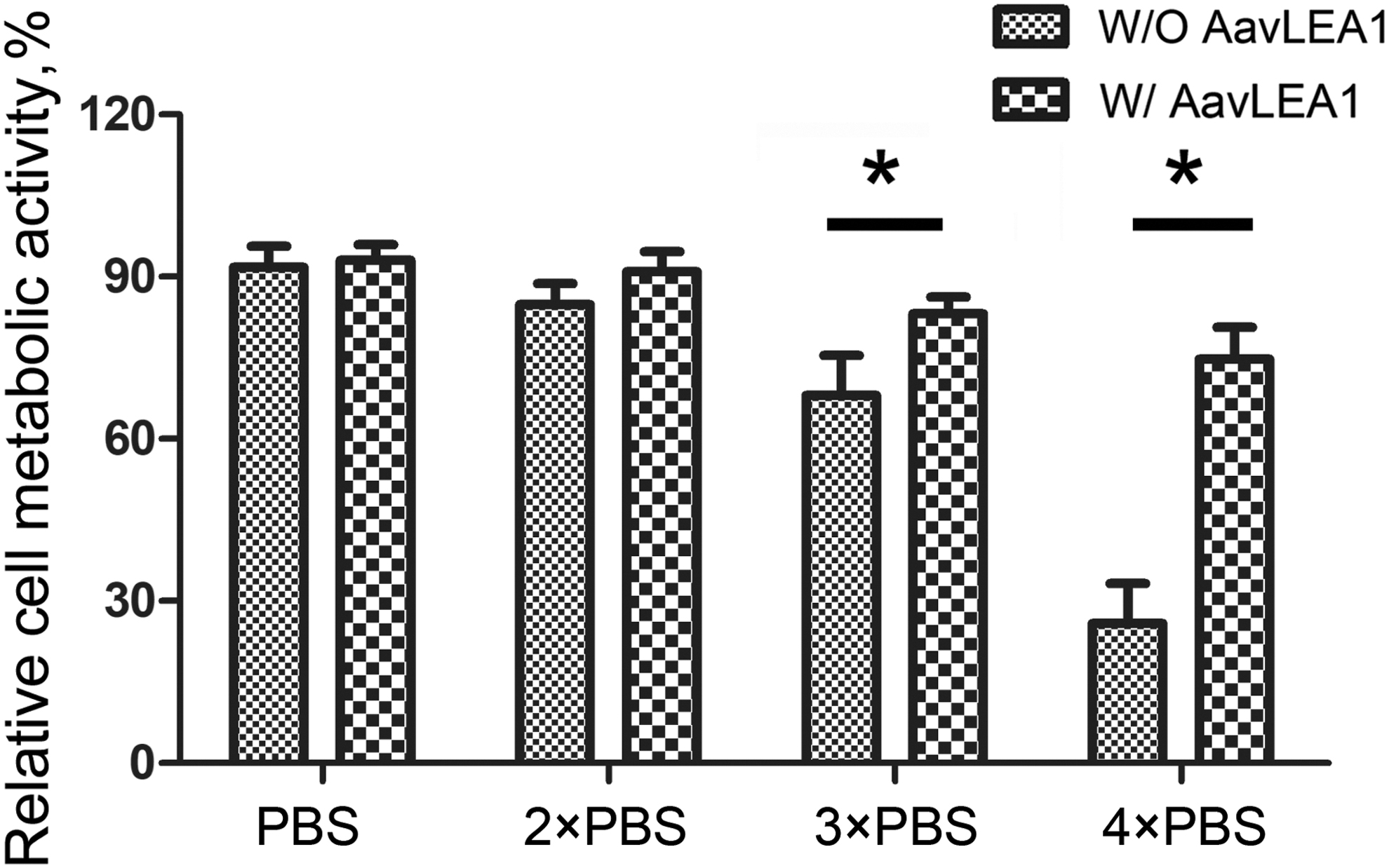
Effects of AavLEA1 proteins on metabolic activity of hUC-MSCs under hyperosmotic stress. hUC-MSCs were maintained in isotonic medium, then replaced to hyperosmotic solution (PBS, 2 × PBS, 3 × PBS, 4 × PBS) without and with recombinant AavLEA1 proteins. The relative metabolic activities of treated cells showed that recombinant AavLEA1 proteins significantly improved hUCM-MSCs hyperosmotic stress at low temperature (*p < 0.05). hUCM-MSCs, human umbilical cord matrix mesenchymal stem cells; PBS, phosphate-buffered saline.
LEA proteins could make cells more resistant to freeze-drying and dehydration. In this study, we researched the effects of LEA proteins on the hUCM-MSCs cryopreservation. After the thawing of cryopreserved hUCM-MSCs, the effects of AavLEA1 proteins on survival were assessed by fluorescence staining. Figure 3 shows representative fluorescence micrographs of the post-cryopreservation hUCM-MSCs stained to detect live/dead (AO/EB) using fluorescence probes. As expected, cell viability of MSCs cryopreserved with 2% DMSO (18.9% ± 8.4%) was low, indicated by the bright red fluorescence of the EB dye and negligible green fluorescence of the AO dye. The cell viability of cryopreservation hUCM-MSCs with 0.1, 0.5, and 2 mg/mL AavLEA1 proteins were 47.1% ± 4.8%, 53.4% ± 12.3%, and 53.9% ± 5.5%, respectively. The viability of postcryopreservation MSCs were higher with the AavLEA1 protein addition, and the viability increased as the concentration AavLEA1 protein increased. The corresponding quantitative data (Fig. 4) show the post-cryopreservation viability of the hUCM-MSCs with various concentrations of LEA proteins. There was a significant difference in the cell viability when AavLEA1 proteins were added during MSC cryopreservation, and the viability of postcryopreservation viability increased in a dose-dependent manner of AavLEA1 protein. The results indicate that AavLEA1 proteins facilitate the hUCM-MSCs viability of post-cryopreservation, and alleviate the cell damage during cryopreservation.
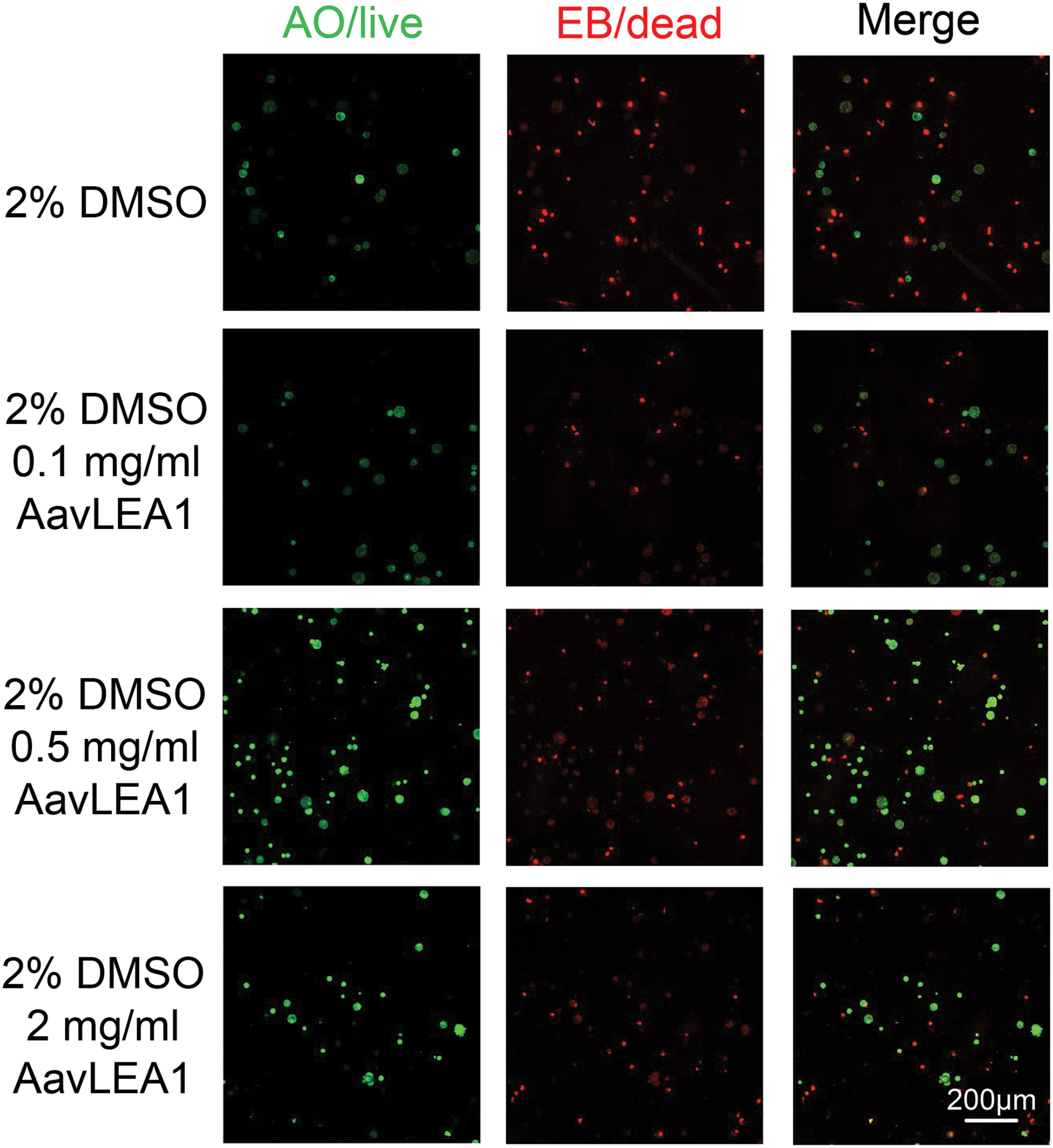
Fluorescence micrographs of hUCM-MSCs postcryopreservation with (0.1, 0.5, and 2 mg/mL) AavLEA1 proteins in 2% DMSO solution. Green fluorescence of the AO dye indicated the living cell, and red fluorescence by the EB dye indicated the necrotic cell or dead cell.
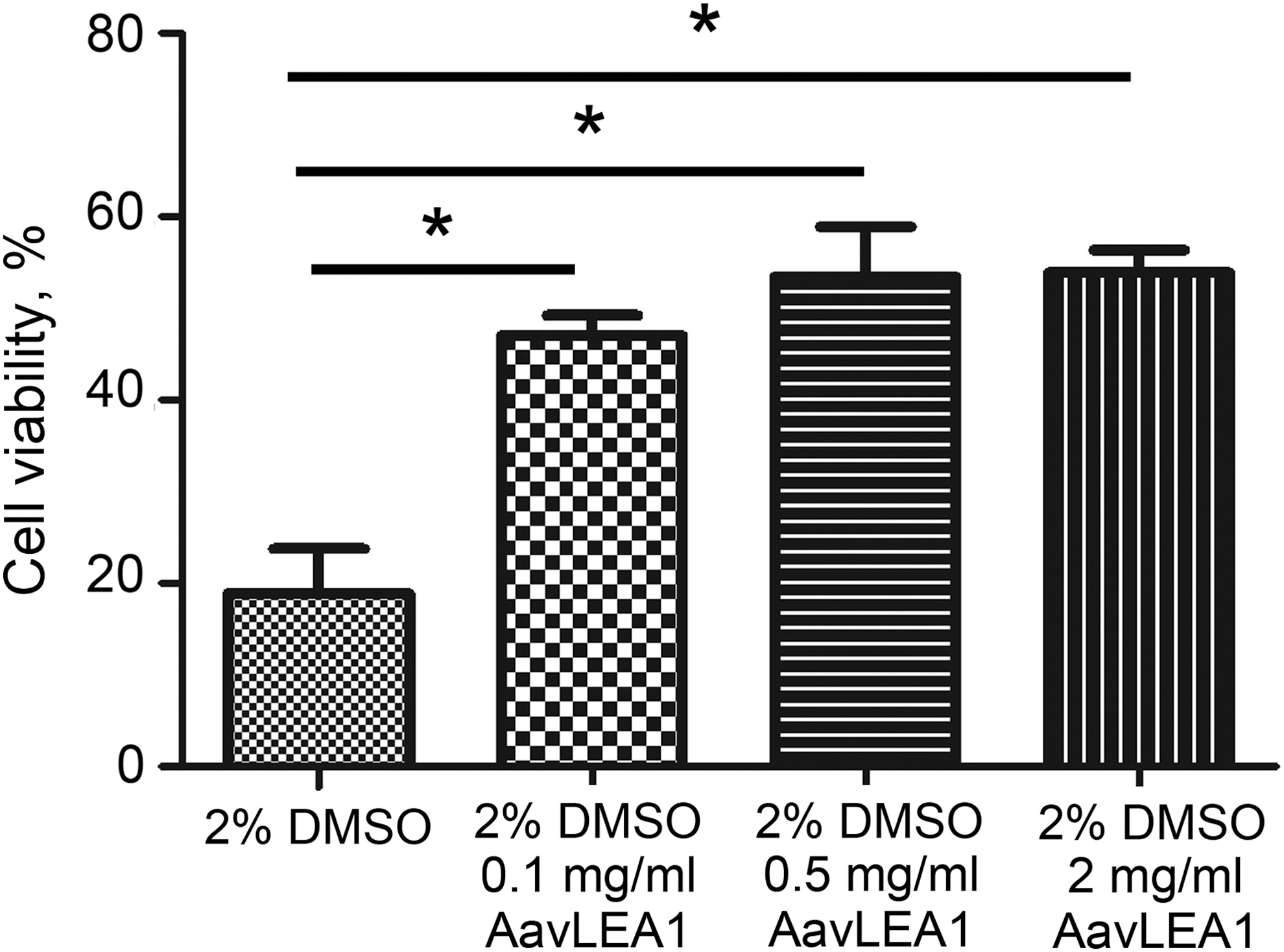
The cell viability of cryopreserved hUC-MSCs with AavLEA1 proteins and comparison cell viability of postcryopreservation by various concentration AavLEA1 proteins with 2% DMSO. Analysis results indicated the hUCM-MSCs viability of post-cryopreservation supplemented with AavLEA1 proteins significantly increased (*p < 0.05).
We further verified effects of recombinant AavLEA1 protein on hUCM-MSCs survival and apoptosis during cryopreservation. The post-cryopreservation effects on primary hUCM-MSCs were tested by flow cytometry with 0.5 mg/mL AavLEA1 proteins at various concentration of DMSO. Figure 5 provides the survival and apoptosis of hUCM-MSCs with the different concentrations of DMSO (i.e., 0%, 2%, 5%, and 10%) and 0.5 mg/mL AavLEA1 proteins. There is only 27.6% of survived cells (Annexin V−/PI−) and 66.4% of apoptotic and necrotic cells cryopreserved with only 2% DMSO. However, there was a significant increase in the live population (67.8%) and a corresponding decrease in apoptotic and necrotic cells (19.9%) with AavLEA1 proteins. The quantitative data (Table 1) show the post-cryopreservation survival and apoptosis rates. Statistical analysis indicated there were significant differences in the cell viability of cryopreservation with addition or no addition of AavLEA1 proteins (Fig. 6). The cell viability increase is not dependent on the concentration of DMSO, with addition or no addition of AavLEA1 proteins. The post-cryopreservation apoptosis sharply decreased with AavLEA1 proteins at 0% and 2% DMSO. However, the post-cryopreservation viability with AavLEA1 proteins is little more than that without LEA protein at 5% and 10% DMSO. These results indicated that AavLEA1 protein can partly replace DMSO during cryopreservation and exhibits the protective effect on the improvement of MSC cryopreservation.
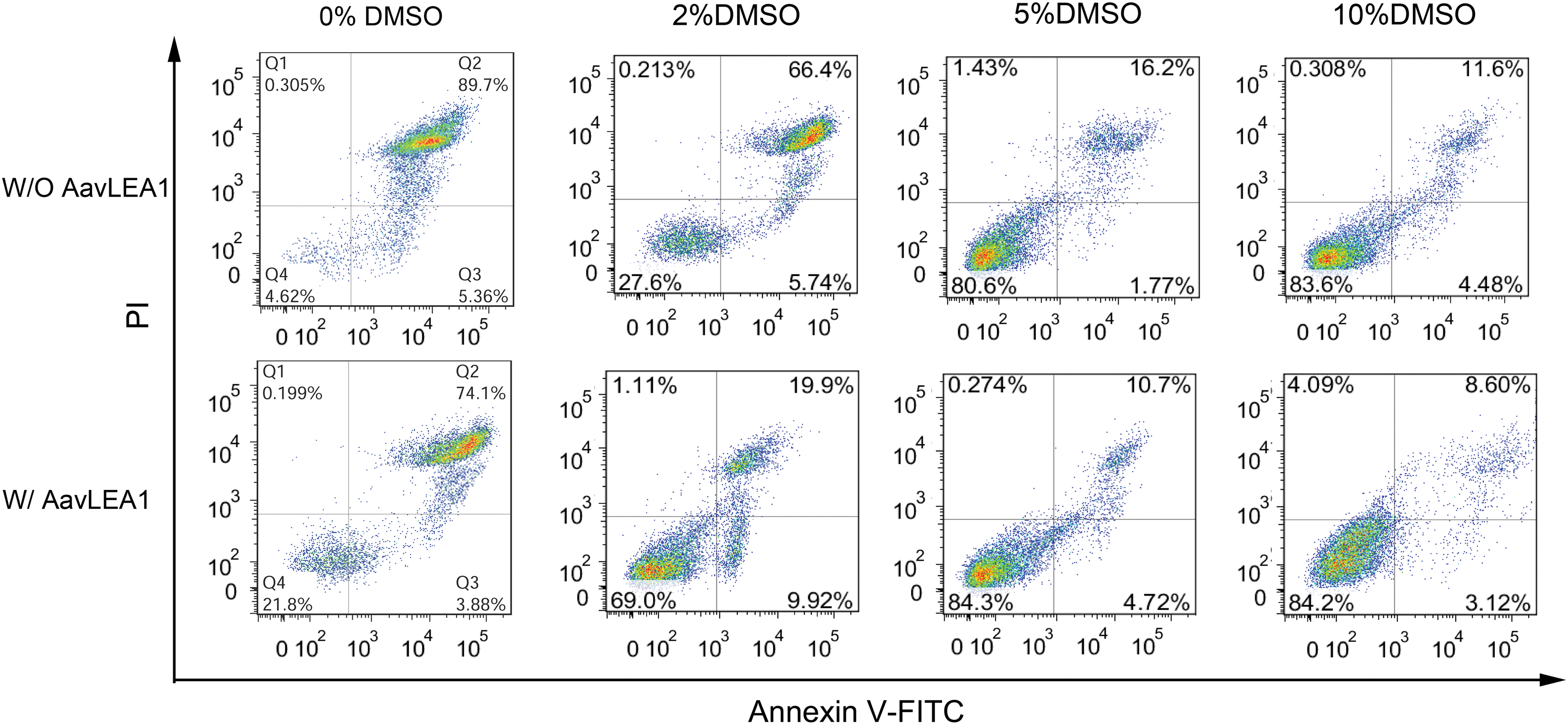
Cell apoptosis assays of cryopreservation hUCM-MSCs with LEA proteins at different concentration of DMSO by flow cytometry. hUCM-MSCs were cryopreserved with 0.5 mg/mL AavLEA1 proteins at the different concentration of DMSO (0%, 2%, 5%, and 10%). After the thawing, hUCM-MSCs were stained by Annexin V-PI for determination of cell apoptosis. LEA, late embryogenesis abundant; PI, propidium iodide. Color images are available online.

The corresponding quantitative data of the survival rate and apoptosis rate of post-cryopreservation, and comparison of survival cell rate and apoptosis cell rate of post-cryopreservation by various concentration DMSO with 0.5 mg/mL AavLEA protein.
The Survival and Apoptosis Rate of Human Umbilical Cord Matrix Mesenchymal Stem Cells Cryopreserved with 0.5 mg/mL AavLEA1 Proteins at Various Concentration of Dimethyl Sulfoxide
Discussion
Despite the fact that conventional permeable CPAs efficiently minimize the cryogenic damage during the freezing process, cytotoxicity and osmotic stress from the use of permeable CPAs are unavoidable. Thus, nontoxic and effective CPAs are especially valuable for cell and tissue cryopreservation. In this study, we extracted the recombinant AavLEA1 proteins and explored the effects of recombinant AavLEA1 proteins on hUCM-MSCs cryopreservation. We confirmed the heat solubility of recombinant AavLEA1 proteins in bacterial lysates, and investigated the effects of the recombinant AavLEA1 protein on osmotic behavior of hUCM-MSCs on ice. We have shown that supplementation of recombinant AavLEA1 protein can improve the efficiency of MSCs cryopreservation and partly replace DMSO to minimize the cryogenic damage.
During freezing, as the concentration of residual extracellular solute increases, the osmotic pressure of the extracellular solution increases. At that time, the intracellular water will flow out through the cell membrane, and the cell membrane will shrink as a result of dehydration and damage to the cell structure. In the case of low temperature and water shortage, cell membranes and proteins are damaged by osmotic pressure and low temperature. In previous studies, LEA proteins induced by dehydration could improve the tolerance of plants and lower animals to desiccation and hyperosmotic stress.14–16 The LEA gene was transfected into HepG2 and HEp-2 cells, LEA proteins were located in mitochondria and could significantly improve the resistance of cells to lyophilization and dehydration.19,21 LEA proteins were found to protect liposomes and cholesterol (similar to cell membrane) during desiccation, 22 and LEA proteins from Artemia franciscana could protect lipid bilayers during cryopreservation. 23 In our study, we applied recombinant AavLEA1 proteins and low concentrations of DMSO to hUCM-MSCs cryopreservation, and AvaLEA1 proteins exhibited protective effects on cell viability.
Owing to a highly hydrophilic polypeptide composed of polar amino acids, most LEA proteins are unstructured and flexible in solution. Many studies have shown that LEA proteins exhibit a differential modulation of the biochemical and biophysical function in desiccation tolerance, and secondary structure analysis showed that LEA proteins adopt a secondary structure when dehydrated.14,16,24,25 LEA proteins could maintain enzyme activity and prevent protein aggregation against freeze-induced damage.11–13 From in vitro studies, it has been shown that AavLEA1 attenuates the loss of enzyme activity in lactate dehydrogenase, citrate synthase, and water-soluble proteomics, and reduces protein aggregation during dehydration.11,12 According to reports, ArLEA1A and PvLEA4 have similar effects.13,14 Many studies have demonstrated that LEA proteins preferentially stabilize specific lipid bilayer membranes.22,23 FT-IR spectroscopy indicates that LEA proteins directly interact with the membrane surface, and they undergo structural transformation, predominantly forming α-helical structures in the dry state.16,26 Structural transformation of LEA proteins is involved in the protection of liposomes in the dry state. The positively charged amino acids of α-helix structures interact with the negatively charged phospholipid head groups, and the nonpolar residues face the hydrophobic core of the membrane.27,28 The α-helix of amphipathic proteins align perpendicular to the phospholipid molecules (i.e., lies flat against the membrane surface), such that the protein settles in between the phospholipid head groups. Moreover, the addition of LEA protein enhances the glass form of the liposome in a dry state and reduces its phase transition temperature from gel to liquid crystal.13,29
In view of the function of the LEA protein, it has been proposed to use the LEA protein as a “molecular barrier” to protect cell membrane during cryopreservation. This may be because of the irregularity and flexibility of the LEA protein and the ability to interact with the cell membrane during freezing and dehydration to undergo structural transformation to stabilize the protein and lipid bilayers. In this study, the metabolic activities of hUCM-MSC treated hyperosmotic solution were measured with AavLEA1 proteins. It was confirmed that recombinant AavLEA1 proteins improved hUCM-MSCs tolerance against hyperosmotic stress at low temperature. We proposed LEA proteins could protect cell membranes, and reduce osmotic pressure and cryogenic damage in cryopreservation by slow freezing. Our results indicate that AavLEA1 proteins can effectively increase the efficiency of hUCM-MSCs cryopreservation, which is of great significance for protecting mammalian cells against cryogenic damage. LEA proteins as nontoxic and effective CPAs provide a novel potential method for a large scale of cell cryopreservation.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest in carrying out this research.
Funding Information
This work was supported by grants from the Scientific Research Foundation of the Institute for Translational Medicine of Anhui Province (No. SRFITMAP 2017zhyx30), the National Key Technology Research & Development Program of China (No. 2017YFC1002004), and the National Natural Science Foundation of China (No. 81901437).