
Abstract
Cryoprotectants are known to have protective effects against cryodamage to spermatozoa. In this study, the cryoprotective effects of two cryoprotectants (glycerol, ethylene glycol) and cryoprotectants/trehalose combinations on frozen-thawed ram spermatozoa were investigated at the ultrastructural level. For this purpose, ejaculates collected from Konya Merino rams were pooled and diluted with a tris-based extender containing additives, including 5% glycerol, 3% glycerol +60 mM trehalose, 1.5% glycerol +100 mM trehalose, 5% ethylene glycol, 3% ethylene glycol +60 mM trehalose, and 1.5% ethylene glycol +100 mM trehalose. They were all cooled to 5°C and then frozen in 0.25 mL French straws in liquid nitrogen. The samples were thawed at 37°C and centrifuged to remove the diluents. Then, they were processed using a scanning transmission electron microscope. In the statistical analysis, the number of ultrastructurally cryodamaged and intact spermatozoa were counted in longitudinal and transverse ultrathin sections in all groups by electron microscopic examination. The amount of intact spermatozoa in the groups containing 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose was found to be higher than other groups (p < 0.05). As a result, it was suggested that the groups of 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose provided the highest protection for the ultrastructural morphology of frozen-thawed Konya Merino ram spermatozoa among the groups.
Introduction
Sperm cryopreservation has recently become widely used in reproductive biotechnology to maintain fertility. However, sperm cryodamage involves cellular damage, including changes in the membranes, mitochondria, acrosomes, and axonemes.1–5 Such cellular cryodamage adversely affects spermatozoa fertility by decreasing the viability of the cells.
Numerous studies have been conducted on the role of various cryoprotectants in cell cryopreservation.6–10 Low-molecular-weight cryoprotectants may prove to allow less damage to the spermatozoa. 11
On the one hand, it was reported that ethylene glycol reduced abnormality and improved sperm viability, acrosome integrity, plasma membrane integrity, conception rate, and pregnancy rate of bull semen compared to glycerol. 12 In another study, Awad 13 compared the cryopreservation effects of low-molecular-weight cryoprotectants (ethylene glycol) to glycerol on post-thawed computer-assisted semen analysis (CASA) sperm parameter in bull semen. They found no advantage to using ethylene glycol to replace glycerol in bull semen freezing. However, they reported that the possibility of using ethylene glycol as a permeating cryoprotectant for bull semen deserved further investigation, and these cryoprotectants should also be evaluated in extenders that contain disaccharides or cholesterol.
Trehalose is a naturally occurring sugar containing two
In this study, we used combinations of trehalose with other cryoprotectants to increase the protective efficacy of the cryoprotectants at low concentrations. We aimed to examine the effects of two cryoprotectants and cryoprotectants/trehalose combinations in different doses on cryodamage in the ultrastructural morphology of frozen-thawed Konya Merino ram spermatozoa with scanning transmission electron microscopy (STEM).
Materials and Methods
Animals, semen collection, and processing
Semen samples from six Konya Merino rams (aged between 3 and 4 years) were used in this study. The animals were housed at the Bahri Dagdas International Agricultural Research Institute and were maintained under uniform nutritional conditions. Ejaculates were collected three times a week using an artificial vagina during the breeding season. Immediately after collection, the ejaculates were immersed in a water bath (37°C) until microscopic assessment in the laboratory. Semen assessment was performed within ∼20 minutes following collection.
The volume of semen ejaculates was measured in a conical tube graduated at 0.1 mL intervals, and the sperm concentration was determined using a hemocytometer. 25 Only ejaculates with a volume of 1–2 mL sperm with >80% progressive motility and a concentration higher than 2 × 109 sperm/mL were pooled to eliminate individual differences. Seminal plasma was not removed before the extending process. Ejaculates collected at the same time were pooled for dividing and then diluting. This process was repeated seven times; thus, seven pooled ejaculates were included in the study.
A tris-based extender (Trizma 254 mM, citric acid 78 mM, fructose 70 mM, egg yolk 15%, pH 6.8) was used as the base extender. Each pooled ejaculate was divided into six equal aliquots and diluted (37°C) with the base extender containing 5% glycerol, 5% ethylene glycol, 3% glycerol and 60 mM trehalose, 3% ethylene glycol and 60 mM trehalose, 1.5% glycerol and 100 mM trehalose, and 1.5% ethylene glycol and 100 mM trehalose at a concentration of 400 million cells/mL. Diluted semen samples were aspirated into 0.25 mL French straws, sealed with polyvinyl alcohol powder, and equilibrated at 5°C for 3 hours. After equilibration, the straws were frozen in liquid nitrogen vapor, ∼4 cm above the liquid nitrogen, for 15 minutes and plunged into liquid nitrogen for storage.
Electron microscopy
After 1 month, straws were thawed individually at 37°C for 25 seconds in a water bath for electron microscopic evaluation. Samples were transferred into Eppendorf tubes and centrifuged at 2500 rpm for 5 minutes. After removal of the supernatants, samples were fixed overnight in Karnovsky solution 26 for ultrastructural analysis by STEM. The samples were then washed three times in 0.1 M cacodylate buffer and postfixed with OsO4 for 1 hour at 4°C. 27 The specimens were dehydrated in an ethanol series and then treated in propylene oxide and embedded in Araldite. The negatively stained ultrathin sections were transferred onto formvar film-coated grids and examined with Field Emission Scanning Electron Microscope (FESEM) (Carl Zeiss, Supra 40 VP) using a STEM detector.
Statistical analysis
In the statistical analysis, at least 500 ultrastructurally cryodamaged and membrane-intact spermatozoa, obtained from four straws for each group, were counted in ultrathin sections by observation with FESEM using a STEM detector. The data were statistically evaluated by chi-square test by using the SPSS/PC software package (version 13.0; SPSS, Inc., Chicago, IL). p-Values below 0.05 were considered statistically significant. This study did not involve humans so IRB approval was not needed.
Results
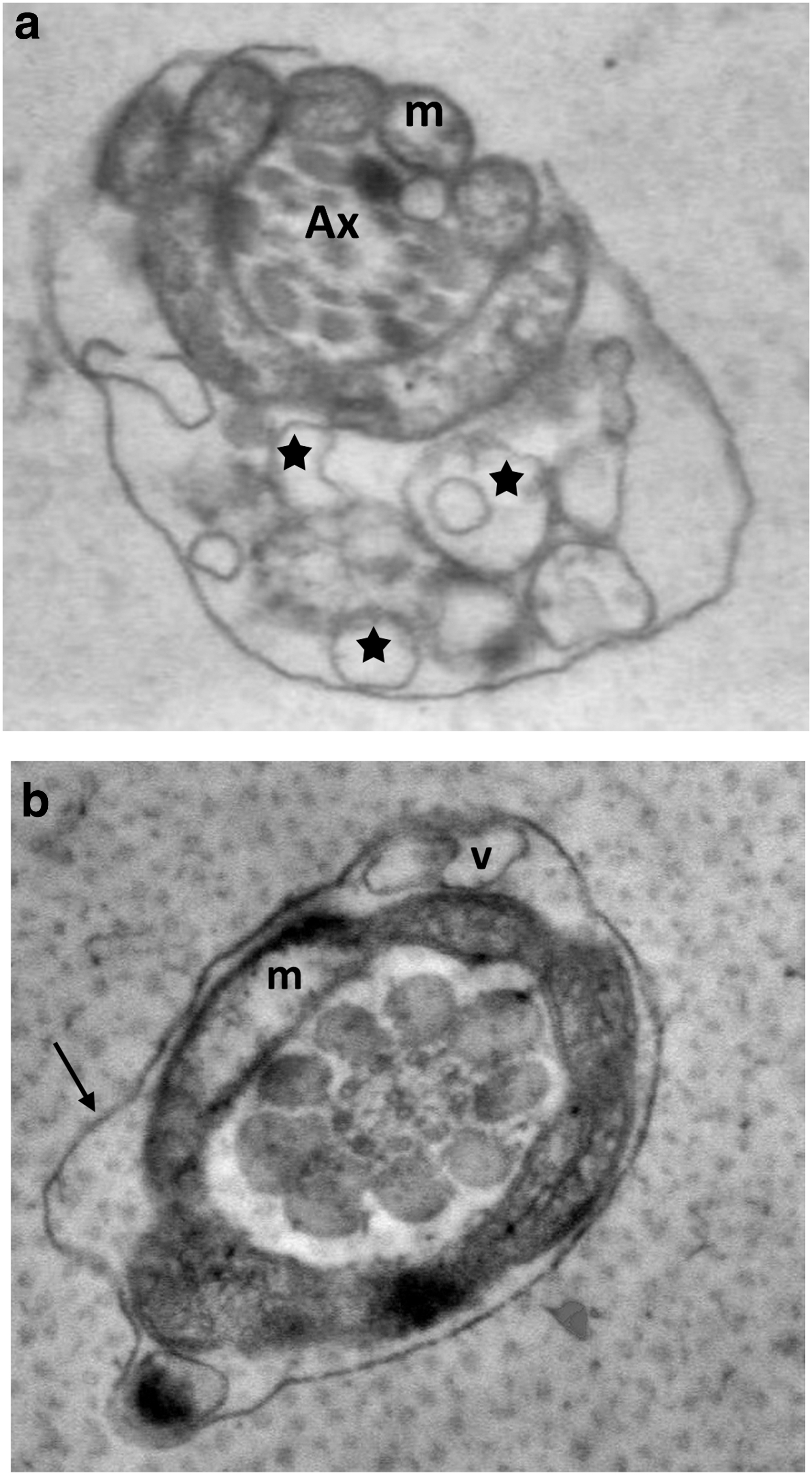
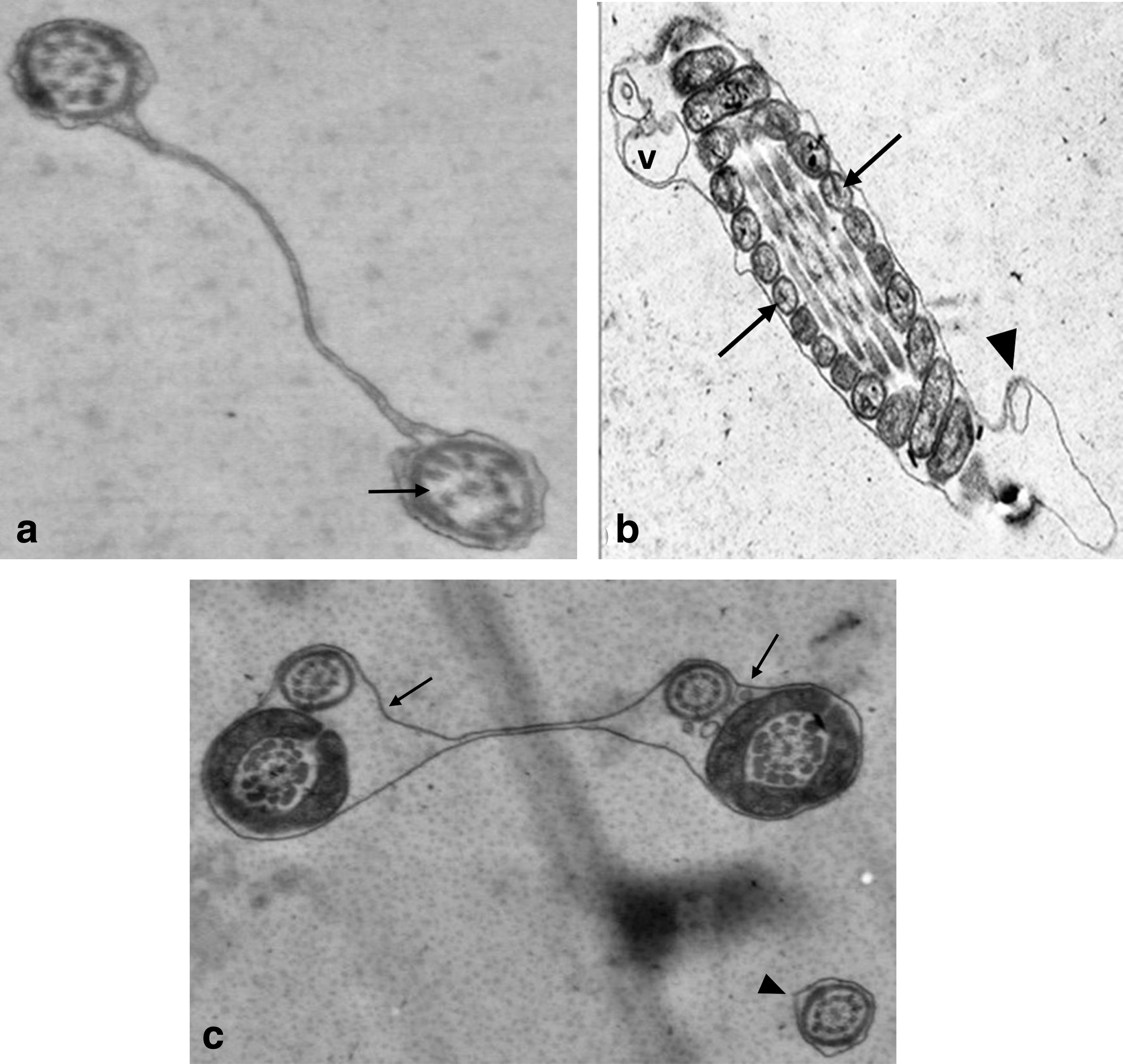
In all groups, ultrastructural cryodamage, including membrane deformations, formation of vesicles, and damaged acrosome, axonemes, and mitochondria, were observed in the STEM micrographs of the spermatozoa. The plasma membrane surrounding the sperm head showed more extensive swelling and ruffled damage (Fig. 1). In the midpiece and distal end of the sperm tails, it was separated from the underlying mitochondrial sheath and axonemal structure (Figs. 1a, 2b, and 3c).

Transverse sections of the tails
In the acrosomal and plasma membranes, blebs along the membranes were observed (Figs. 1e and 4b). In addition, an atypical acrosome reaction exhibited knob-like protrusion of acrosomal material (Fig. 4a). As for axonemes, the 9 + 2 microtubule pattern was abnormal, and therefore, differences in the arrangement of the axonemal complex and the loss of axonemal doublets were visualized in the transverse sections of the sperm tails (Fig. 5). In addition, damaged mitochondria with an electron lucent matrix and absence of cristae in the transverse (Fig. 2) and longitudinal (Fig. 3b) sections of the sperm tails and plasma membrane-derived vesicles (Fig. 2a) were observed. This cryodamage was consistent with previous ultrastructural studies of sperm cryopreservation.

Longitudinal sections of sperm heads

Tail defects, showing axonemal alterations. Note that the two axonemal structures (arrowhead) in a common MS and disorganization (dashed arrow) and absence of axonemal microtubules (arrows) × 100,000. MS, mitochondrial sheath.
In the statistical analysis, mostly membranes, and also mitochondrium (electron lucent matrix/absence of cristae), and axoneme (loss of axonemal doublets) ultrastructural cryodamage in frozen-thawed Konya Merino ram spermatozoa were evaluated in the longitudinal and transverse sections for each group. According to the statistical analysis, the amount of membrane intact spermatozoa in 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose showed the highest cryoprotective effect on the ultrastructural morphology of Konya Merino ram spermatozoa of all the groups (Table 1).
Mean Intact Sperm in Semen Supplemented with Different Cryoprotectants and Trehalose of Rams Following Freeze-Thawing
Different superscripts within the same column demonstrate significant differences among groups (p < 0.05).
Discussion
The sperm cryopreservation process induces damage and severe osmotic changes in cells due to low temperature. Damage to the cell membrane through cryopreservation leads to many changes in cellular structures and their associated functions, 28 which result in lowered sperm fertilization capacity. 29 It has been shown in many studies that cryopreservation leads to various deformations in the sperm's structure.1,30–33
In recent years, intensive studies have been carried out on the use of cryoprotectants, such as dithioerythritol, vitamin E, trehalose, and glycerol in rams,24,34–36 and cysteine, ethylene glycol, trehalose, fetuin, and glycerol in bulls37–39 against cryodamage during cryopreservation. Alcay et al. 34 demonstrated the cryoprotective effects of 6% glycerol on motility and plasma membrane integrity, and 6% ethylene glycol on acrosomal and DNA integrity in frozen-thawed ram semen.
In addition, it has been concluded that 5% glycerol and 3% or 5% ethylene glycol protect ram spermatozoa against the harmful effects of freezing and that 5% glycerol offers greater protection to the plasma membrane of the sperm. 40 Alvarenga et al. 41 reported that the percentage of motile stallion spermatozoa was higher (p < 0.05) for sperm frozen in the presence of 5% ethylene glycol. In this study, we found that the highest percentage of ultrastructurally membrane-intact sperm among the groups was in the base extender containing 5% ethylene glycol.
Malo et al. 42 reported that trehalose increases the viability of spermatozoa and in vitro fertilization parameters in the cryopreservation of boar spermatozoa. It was reported that a 50 mM concentration of trehalose resulted in the highest percentage of membrane-intact sperm in Chios rams. 43 Also, Buyukleblebici et al. 38 also demonstrated the beneficial cryoprotective effect of 25 mM trehalose and 3% ethylene glycol on acrosome morphology, and 3% glycerol on membrane integrity in bull semen. In our study, the best ultrastructural cryoprotective effect was found in 100 mM trehalose +1.5% ethylene glycol in Konya Merino ram spermatozoa.
Gutiérrez-Pérez et al. 44 determined that a mixture of 1% glycerol and trehalose (250 mM) showed the best results in sperm viability and motility in boars. One hundred millimolar trehalose or raffinose in the tris-citric acid-fructose-egg yolk extender reduced sperm anomalies in rams. 45 During the cryopreservation of boar semen, 46 the replacement of cryoprotectant glycerol (100 mM) with 100 mM trehalose significantly improved motility, mitochondrial membrane potential (MMP), and acrosome integrity. Sperm motility and MMP were considerably higher in 100 mM trehalose, whereas the acrosome integrity was substantially higher in 100–250 mM trehalose. El-Sheshtawy and Sisy 47 suggested that 50–100 mM trehalose or sucrose is more useful in the cryopreservation of bull semen.
In addition, Aboagla and Terada 20 reported that motility parameters markedly improved with the increase in trehalose concentration in goat spermatozoa. They also suggested that the cryoprotective effects of trehalose may be due to enhanced sperm membrane fluidity before freezing. Also, 100 mM trehalose showed a high cryoprotective effect on motility, mitochondrial activity, and integrity of the membrane and acrosome in boar spermatozoa. 48 Naing et al. 49 showed that the combination of glucose and trehalose improves semen quality in cryopreservation of goat semen, and 198.24 mM trehalose provided a higher contribution. In frozen ram sperm samples, 100 mM trehalose significantly increased the vitamin E level in samples compared to the controls, and 50 mM trehalose provided a high rate of motility after thawing; however, there was no significant difference in acrosome and total abnormalities. 17
Uysal and Bucak 24 reported that 100 mM trehalose has superior cryoprotective effects on motility, morphological anomalies, viability, and membrane integrity according to 50 mM trehalose in ram semen. Similarly, in our study, a high concentration of trehalose (100 mM), but in 1.5% ethylene glycol, protected the ultrastructural morphology of frozen-thawed Konya Merino ram spermatozoa. With the ultramicroscope, Aisen et al. 6 showed a significant reduction in the proportion of sperm membrane damage in the hypertonic extenders (tris-citrate modified solution) plus 76 g/L trehalose. In this study, ultrastructurally intact spermatozoa were determined in the 100 mM concentration of trehalose in 1.5% ethylene glycol using electron microscopy techniques.
It has been reported that the protective effect of trehalose is due to both to its osmotic effect and specific interactions with membrane phospholipids by rendering hypertonic media, causing cellular osmotic dehydration before freezing, and then decreasing the amount of cell injury through ice crystallization.33,50–52
Since spermatozoa are very tiny cells, observation of organelles requires higher magnifications. In this sense, electron microscopy techniques have the advantage of examining ultrastructural changes of spermatozoa after freezing and thawing processes. Ozturk et al. 53 showed that the postthaw sperm parameters (acrosome integrity, DNA fragmentation, and DNA Integrity) could be improved by the supplementation of the semen extender with trehalose, which is shown in Table 2.
Mean (±SE) Sperm Acrosome Integrity, DNA Fragmentation, and DNA Damage of Ram Semen in Presence of Glycerol, Ethylene Glycol, and Trehalose Following Cryopreservation
Source: Ozturk et al., 2020. 53
Means with different letters (a, b, c) in the same column demonstrate significant differences (p < 0.05).
In the present study, the addition of trehalose resulted in protection for the sperm ultrastructural morphology, showing a correlation with the previous study. 53 We observed that the 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose provided the highest protection for the ultrastructural morphology of cryopreserved ram spermatozoa. Thus, it can be postulated that 1.5% ethylene glycol could play a role as a cocryoprotectant with 100 mM trehalose on cryopreserved ram spermatozoa.
The cryopreservation-caused changes determined in our study were similar to the changes in some sperm ultrastructures shown in the different mammalian species.32,33,54,55 Sa-Ardrit et al. 56 showed that the freezing and thawing procedure caused structural damage, especially in the plasma membrane, acrosome, and mitochondria in elephant spermatozoa. They also reported that fluorescence and electron microscopic evaluations were potentially a powerful tool in the analysis of elephant spermatozoa after freezing and thawing. Ozkavukcu et al. 3 concluded that cryopreservation had deleterious effects on spermatozoa, especially on plasmalemma, acrosomes, and tails in human spermatozoa.
Lopez-Armengol et al. 57 showed ultrastructurally that plasma and acrosomal membranes were damaged in frozen-thawed ram spermatozoa. They reported that cryoinjury occurred principally at the plasma membrane and could be present or absent in all regions. The cryodamage that occured in the plasma membrane may be elucidated by the freeze/thaw-induced stress effect on the plasma membrane fluidity, as mentioned previously. 58 In the present study, mostly membrane cryodamage as well as mitochondrium (electron lucent matrix/absence of cristae) and axoneme (loss of axonemal doublets) cryodamage were observed by the STEM. The slightly swelled acrosome and damaged mitochondria observed in this study resembled an atypical acrosome reaction structure and damaged mitochondria with distorted cristae described in the study by Khalil et al. 59
The increased production of reactive oxygen species could explain the decrease in sperm motility, the reduced mitochondrial activity, and plasma membrane, acrosomal, 60 and axonemal 61 damages. Therefore, the sperm cryodamage that occured during the freezing process, which led to reduced fertilization capacity of sperm, is a result of the formation of reactive oxygen species. According to the deleterious effects of oxidative stress on sperm, cryoprotectant-related studies have great importance in reducing the number of cryodamaged sperm during the freezing process. It has been reported that membrane-permeating cryoprotective agents (glycerol, ethylene glycol) stabilize membranes and modulate the rate of cellular dehydration. 62 In terms of trehalose as a nonpermeating cryoprotectant, it stabilizes the cell membrane and prevents deleterious effect of cell dehydration on the plasma membrane. 63
Taken together, in the present study, the investigated cryoprotective effects of two cryoprotectants (glycerol, ethylene glycerol) and combinations of trehalose with these cryoprotectants, the best cryoprotective effect on the ultrastructural alterations in the Konya Merino ram spermatozoa was statistically found with 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose.
In conclusion, this study revealed again the importance of investigations of cryoprotectants for sperm cryopreservation. Groups of 5% ethylene glycol and 1.5% ethylene glycol +100 mM trehalose provided the highest protection for the ultrastructural morphology of frozen-thawed Konya Merino ram spermatozoa among the groups studied. Furthermore, it can be argued that, considering the importance of investigating the effects of cryoprotectants on the ultrastructural morphology of spermatozoa, this research has the potential to improve the sustainability of healthy genetic strains of various species.
Footnotes
Acknowledgments
This study was approved by Pamukkale University Animal Ethics Committee (No: PAUHDEK- 2014/014) and supported by the Scientific and Technological Research Council of Turkey (TUBITAK) (Project No: 114O642).
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The Scientific and Technological Research Council of Turkey (TUBITAK) (Project No: 114O642).