
Abstract
Vitamin E is one of the most powerful antioxidants for prevention of cell damage resulting from cryopreservation, but its efficacy for cryopreserving brown trout sperm is still unclear. In this work, the protective effect of vitamin E on quality, fertilizing capacity, and DNA damage of brown trout (Salmo trutta macrostigma) sperm after cryopreservation was evaluated. Sperm samples were diluted at the ratio of 1:10 with three different extenders (E): (E-I): 300 mM glucose, 10% egg yolk; (E-II): 33.3 mM glucose, 5.1 mM NaCl, 0.5 mM NaHCO3,, 15% DMA; and (E-III): 61.6 mM NaCl, 134.2 mM KCl, 1.9 mM CaCl2, 0.8 mM MgCl2, 2.3 mM NaHCO3 in distilled water. Each extender was supplemented with 10% DMSO and different concentrations of vitamin E at 0.1, 0.5, and 1.0 mM. Spermatozoa frozen without vitamin E (0 mM, control) and fresh sperm were also used. After dilution, the sperm was aspirated into 0.25 mL straws, frozen 3 cm above the liquid nitrogen (LN2) surface, and plunged into the LN2. Cell motility, viability, fertilization, and eyeing were determined in post-thawed samples. DNA damage was determined by the comet assay after cryopreservation. Supplementation of 1 mM vitamin E to all extenders exhibited the best cryoprotective effect in terms of sperm motility, duration of motility, viability, fertility, and DNA integrity against cryopreservation damage, compared with 0.1, 0.5, and control group (0 mM) (p < 0.05). The highest post-thaw motility (62.4% ± 0.36%), fertilization (48.2 ± 0.84), and the lowest DNA damage (7.245%) were obtained with the extender-II including 1.0 mM vitamin E (p < 0.05). Consequently, vitamin E positively affected the motility parameters, fertility, and DNA integrity, and the results suggest the addition of extenders with vitamin E as an antioxidant for the cryopreservation of brown trout sperm.
Introduction
Cryopreservation is a long-term storage technique that is efficiently used as an important biotechnological tool for the aquaculture industry. Its benefits can be summarized as follows: synchronization of the availability of the gamete year-round, reduction of the nursing cost of broodstock, promotion of breeding techniques for broodstock management, and development of the hatchery management by protecting sperm of the selected individuals in cryobanks.1,2 Thus, the improvement of effective cryopreservation methods is very important to facilitate the use of cryopreserved sperm for large-scale larvae production.
Salmo trutta macrostigma is one of the important freshwater fish species in terms of biodiversity and its economical value. Unfortunately, its population declines day by day because of illegal practices and heavy fishing pressure, degradation of spawning habitats, and river damming.3,4
Cryopreservation of salmonid sperm has been widely investigated because of its high commercial and recreational value in terms of the food sector and fishing. On the contrary, the sperm cryopreservation process, despite its various advantages, causes cellular damage that can reduce semen quality such as motility, membrane stability, DNA integrity, and also it impairs its fertilization capacity and embryo development.3–6 The main cause of the damage in sperm during the cryopreservation process is oxidative stress owing to the inconsistency between reactive oxygen species (ROS) and antioxidant activity of sperm cells.
Naturally, sperm cells are protected against oxidative stress reactions with antioxidant components of the seminal plasma. On the contrary, dilution of sperm causes a reduction of seminal plasma ingredients and an increase in the susceptibility of sperm cells to oxidative stress during the cryopreservation process. 7 It has been proven that by reducing the oxidative damage it is possible through supplementation of some substances, which have antioxidant properties, to the cryosolutions, the sperm cells can be protected against damaging effects of ROS. 8 However, antioxidant effectiveness can vary among species and is dependent upon its concentration and structure.
Cabrita et al. 7 reported that cryosolutions containing vitamins with antioxidant properties decreased DNA fragmentation in gilthead seabream (Sparus aurata) when compared with that in European sea bass (Dicentrarchus labrax). From these results it is known that vitamin E (α-tocopherol) is one of the natural antioxidants, which has the ability to penetrate the sperm membrane and protect the sperm cells against the detrimental influence of free radicals. 9 In addition, it can neutralize lipid peroxidation and also decrease oxidative damage. 10
However, to the best of our knowledge, there is a lack of information concerning whether vitamin E could improve the quality and decrease DNA damage of sperm after the cryopreservation process in brown trout. In this framework, the objective of this study was to evaluate the influence of three different extenders (ionic and sugar based) with different vitamin E concentrations (0.1, 0.5, and 1.0 mM) on post-thaw quality, fertilization ability, and DNA damage after cryopreservation of brown trout semen.
Materials and Methods
Ethical approval
This study was approved by the Local Animal Experiment Ethics Committee of the Mustafa Kemal University (1005-M-0111) in Antakya, Hatay.
Animals
Brown trout gametes were obtained from 2- to 3-year-old broodstock at the Kılıç Bafa Aquaculture Farm (Kahramanmaraş, Turkey) in January. Mature male (386.2 ± 5.7 g, n = 20) and female (427.5 ± 4.2 g, n = 10) brown trout were kept in 500 L indoor tanks, which were supplied with constant hatchery water (8.2°C ± 1.24°C; 10.6 ± 1.5 mg/L O2) flow at rate of 2.0 L/s. Before sampling, individuals were anesthetized with 15 mg/L quinaldine (Sigma) for a few minutes.
Collection of gametes
For sperm collection, the external urogenital pore of the mature males was wiped dry with a towel to avoid contamination with water, urine, and feces. Then sperm was collected by gentle abdominal massage into 50 mL sterile glass tubes separately. The sperm-containing tubes were placed into a styrofoam box including crushed ice (4°C ± 1°C) until evaluation for quality in terms of motility, density, pH, and color. For egg collection, mature females were wiped dry and stripped by gentle abdominal massage. The eggs were collected into a 1000 mL round-bottomed bowl. The collected eggs were evaluated for homogenous shape, color, and size, and used for fertilization within 30 minutes of stripping.
Sperm cryopreservation
The motility of selected sperm samples were evaluated with the aid of an activation solution (AS) (45 mM NaCl, 5 mM KCl, and 30 mM Tris-HCl, pH 8.2). 11 For this aim, 1 μL of sperm suspension was placed on a glass slide and activated by adding 100 μL AS, and sperm motility was determined using a CCD video camera (model SPC-2000P; Japan) mounted on a phase-contrast microscope at 100 × magnification (BX43; Olympus, Tokyo, Japan). The observed video records were scanned to evaluate the percentages (%) and duration (s) of motility at least three times for each sample. Sperm motility was evaluated as the percentage of cells exhibiting progressive forward movement, whereas the duration of motility was evaluated until forward movement stopped.
Spermatozoa density was determined according to the hemocytometric method. For this aim, sperm were diluted at a ratio of 1:1000 with Hayem solution (35.2 mM Na2SO4, 17.1 mM NaCl, 1.8 mM HgCl2, 200 mL bicine) and density was determined using a 100 μm deep Thoma hemocytometer (TH-100; Hecht-Assistent, Sondheim, Germany) at 400 × magnification with an Olympus BX50 phase contrast microscope (Olympus) and expressed as spermatozoa × 109 mL−1 (three replicates). 12 Standard pH electrodes were used to measure sperm pH, and semen color was evaluated visually within 30 minutes after sperm collection.
Sperm obtained from 18 males exhibiting >80% motility were individually split into three subsamples and each of them diluted at a ratio of 1:10 (v:v) with three different extenders (glucose and ionic based) composed of (E-I) 12 : 300 mM glucose, 10% egg yolk; (E-II) 13 : 33.3 mM glucose, 5.1 mM NaCl, 0.5 mM NaHCO3, 15% DMA; and (E-III) 14 : 61.6 mM NaCl, 134.2 mM KCl, 1.9 mM CaCl2, 0.8 mM MgCl2, 2.3 mM NaHCO3 in distilled water. After dissolving vitamin E in 50 mL ethanol, 15 each extender was supplemented with 10% DMSO and vitamin E at 0.1, 0.5, and 1.0 mM concentrations separately. The selected concentrations of vitamin E were based on previous studies [7, 24]. Control groups for each extender were supplemented only with 10% DMSO.
Diluted sperm was equilibrated at 4°C for 10 minutes and then drawn into 0.25 mL straws (IMV, France) and sealed with polyvinyl alcohol (PVA). Sperm concentration was ∼12.5 × 106 in each 0.25 mL straw. Before freezing, diluted sperm were maintained in a cool chamber at 4°C to obtain isothermal conditions. Sperm samples were frozen 3 cm above the liquid nitrogen (LN2) surface in a styrofoam box for 10 minutes and then the frozen samples in each experiment were plunged into LN2. Nine straws per sperm sample were frozen. After 1 week storing of samples in LN2, rewarming was performed in a water bath at 30°C for 20 seconds. After this, samples were examined in terms of motility, viability, fertilizing capacity, and DNA integrity.
Sperm analysis
To evaluate the frozen–thawed sperm samples, a 10 μL drop of the sperm dilution was placed on a microscope slide, 20 μL (AS) was added and then the sperm suspensions were thoroughly mixed for 2 seconds. The motility and motility duration of spermatozoa were immediately recorded for 1 minute after activation using a CCD video camera (CMEX-5, the Netherlands) mounted on a phase-contrast microscope (100 × ; BX43; Olympus). Sperm motility was evaluated from sperm performing progressive forward movement, whereas motility duration was determined by counting the time from activation until it stopped moving. 11 At least five straws were used for evaluation of each parameter and it was performed three times for each treatment.
To assess the percentage of viable sperm, eosin-nigrosin preparations were formed according to the method defined by Björndahl et al. 16 and a total of 300 sperm cells were counted on each slide at a 1000 × magnification. Sperm showing partial or complete purple colorization were considered nonviable and only sperm showing strict exclusion of the stain were considered viable.
Fertilization ability of cryopreserved sperm
The good quality eggs pooled from four females were divided into batches of about 100 eggs and fertilized with 100 μL frozen–thawed sperm or 25 μL fresh sperm (sperm-to-egg ratio: ∼ 5 × 106:1) in 500 mL round-bottomed bowls. Next, the fertilization solution (125 mM NaCl, 20 mM Tris-HCl, 30 mM glycine, pH 9) was added onto the eggs in a ratio of 1:2 (fertilization solution/eggs).17,18 The eggs were slightly stirred for 30 minutes and rinsed with hatchery water (8.2°C ± 1.2°C; 10.6 ± 1.5 mg/L O2) at the end of stirring, and finally transferred into the labeled vertical incubators supplied with flowthrough hatchery water. Eggs were incubated ∼40 days until the eyed stage. The fertilization (number of 4-cell stage embryos/number of total eggs) rates were determined under a stereomicroscope at 20-fold magnification. Three replicates were performed for each treatment with frozen–thawed and fresh sperm.
Evaluation of sperm DNA damage
DNA damage of sperm cells was explored using the single cell gel electrophoresis (COMET) assay that was generally carried out under neutral conditions. The performed method was similar to several protocols with few modifications that were applied to previous studies.19,20 First, diluted sperm samples were centrifuged at 300 g for 10 minutes at 4°C. Seminal plasma was removed and remaining sperm cells were washed with PBS, then adjusted to a concentration of 1 × 105 spermatozoa/mL. For this aim, each microscope slide was precoated with a layer of 1% normal melting point agarose in PBS and thoroughly dried at room temperature. Next, 100 μL of 0.7% low melting point agarose at 37°C was mixed with 10 μL of the sperm samples and dropped on top of the first layer. Slides were allowed to solidify for 5 minutes at 4°C in a moist box.
The coverslips were removed and the slides were immersed in freshly prepared cold lysis buffer with 2.5 M NaCl, 100 mM Na2-EDTA, 10 mM Tris, 1% Triton X-100, 10% DMSO, and 40 mM dithiothereitol (pH 10) for 1 hour at 4°C. Then, the slides were incubated overnight at 37°C in 100 μg/mL proteinase K and added to the lysis buffer. The slides were removed from the lysis buffer, drained, and placed in a horizontal electrophoresis unit filled with fresh alkaline electrophoresis solution, containing 300 mM NaOH and 1 mM EDTA (pH 13), for 20 minutes to unwind the DNA. Electrophoresis was performed for 20 minutes at room temperature at 25 V and was adjusted to 300 mA. Then, the slides were washed with a neutralizing solution of 0.4 M Tris, pH 7.5. After neutralization, the slides were stained with 50 μL of 2 μL/mL ethidium bromide and covered with a coverslip. The whole process was performed under dim light to prevent further DNA damage.21,22
The images of 100 randomly chosen nuclei were analyzed visually. Observations were made at a magnification of 400 × using a fluorescent microscope (BX51; Olympus). Damage was detected by a tail of fragmented DNA that migrated from the sperm head, causing a “comet” pattern, whereas whole sperm heads, without a comet, were not considered as damaged. All data were expressed as a mean percentage undamaged sperm heads ± SEM.
Statistical analysis
All analyses were repeated three times for each treatment. Results were presented as mean ± standard deviation. Motility values were normalized through arcsine transformation and differences among the parameters were tested with two-way analysis of variance (extender × vitamin E). Correlation analyses were performed to determine the interactions between tested extenders and vitamin E concentrations. Duncan post hoc test was implemented for all comparisons among the treatments at level of p < 0.05. All statistical analyses were carried out using SPSS 17.0 software package program.
Results
Sperm quality parameters
In fresh sperm, the mean percentage (%) and duration (s) of motile spermatozoa were 80.55% ± 13.81% and 47.22 ± 13.15 seconds, respectively. Mean spermatological properties of fresh sperm are given in Table 1. The effect of different extenders containing different concentration of vitamin E on post-thaw progressive motility, motility duration, and viability parameters of frozen–thawed sperm are given in Figures 1–3. Increasing the vitamin E concentration in all extender types caused an increase in all mentioned post-thaw quality parameter values in comparison with those that not containing vitamin E (0 mM) (control group) (p < 0.05). The highest post-thaw average motility, motility duration, and viability values were attained with the extender-II containing 1 mM vitamin E concentration. Statistical differences were significant among the treatments in terms of motility, duration, and viability of frozen–thawed sperm (p < 0.05).
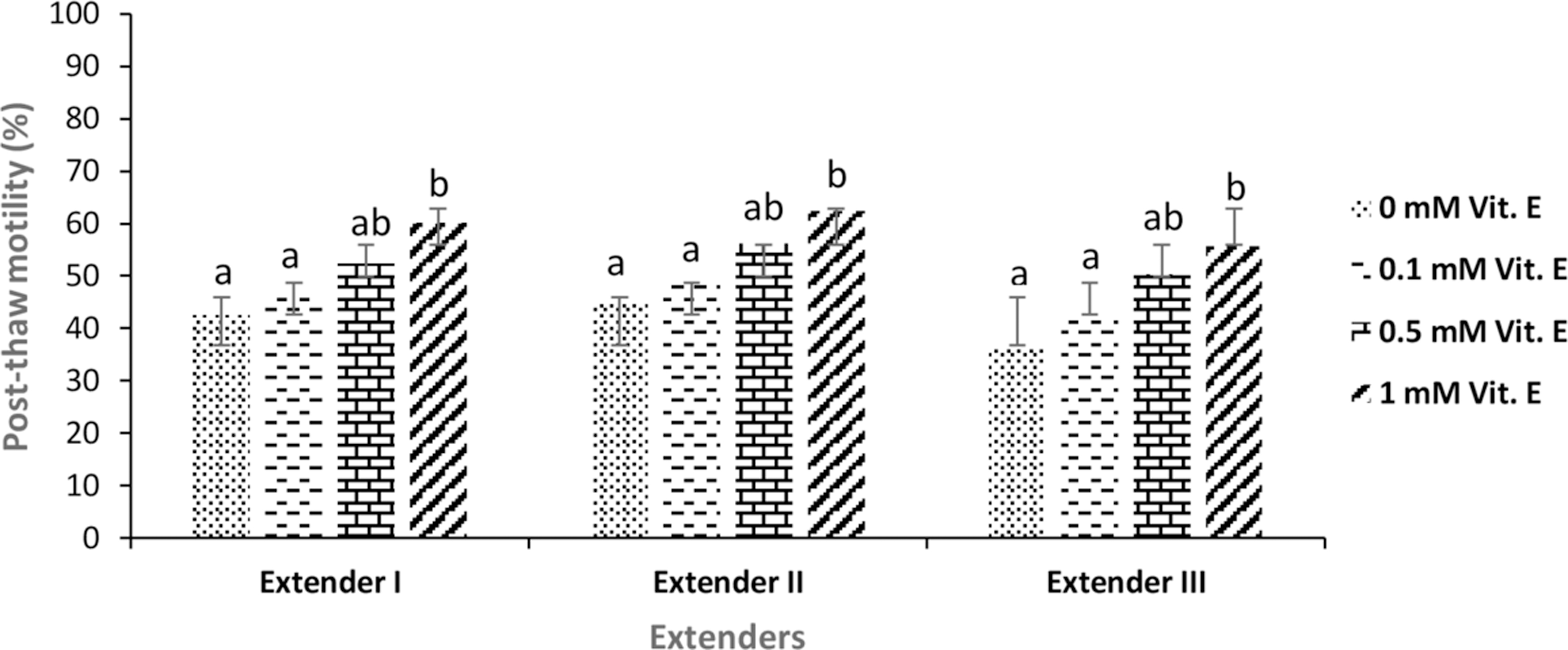
The mean post-thaw motility (%) of frozen–thawed brown trout (Salmo trutta macrostigma) sperm. Different letters indicate differences among treatments (ANOVA, p < 0.05, n = 9). ANOVA, analysis of variance.
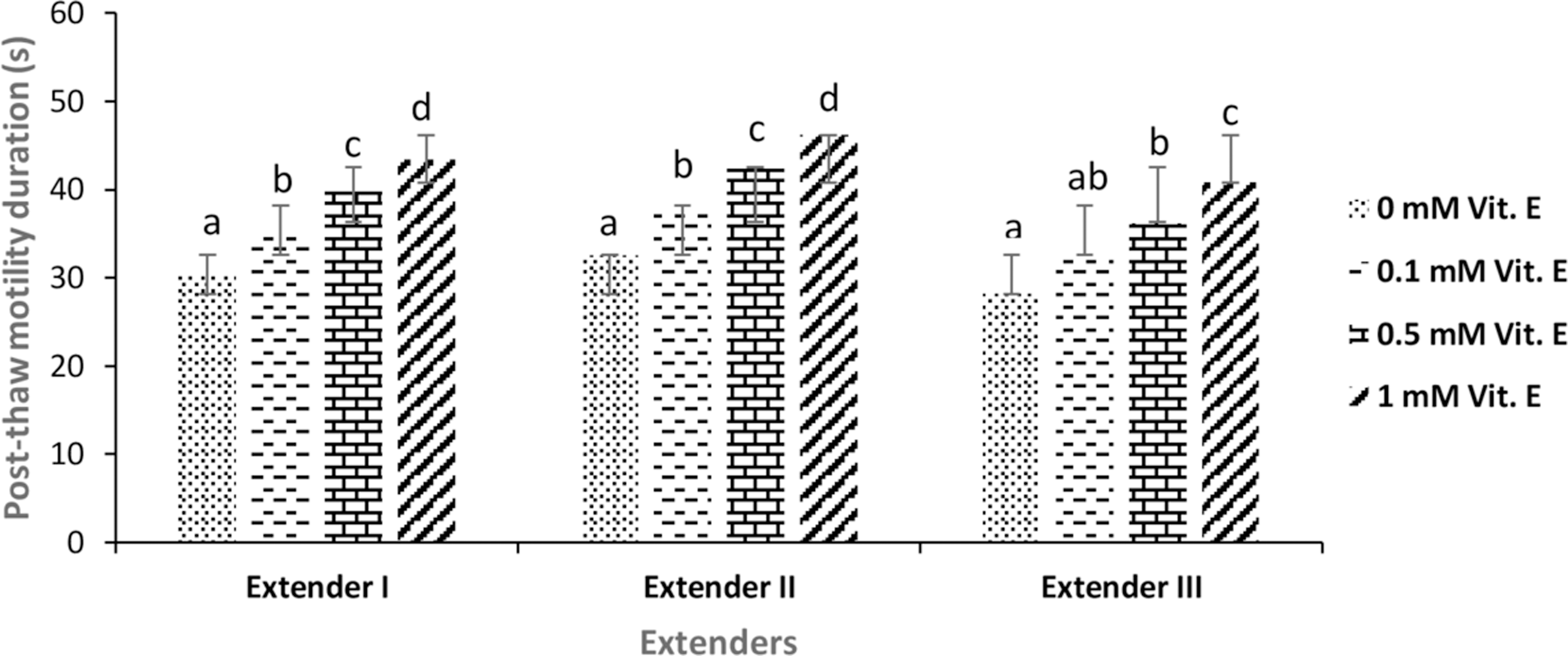
The mean post-thaw motility duration (s) of frozen–thawed brown trout (Salmo trutta macrostigma) sperm. Different letters indicate differences among treatments (ANOVA, p < 0.05, n = 9).

The mean post-thaw viability (%) of frozen–thawed brown trout (Salmo trutta macrostigma) sperm. Different letters indicate differences among treatments (ANOVA, p < 0.05, n = 9).
Mean Spermatological Properties of Fresh Brown Trout (Salmo trutta macrostigma) Sperm (n = 20)
Fertility
Increasing the vitamin E concentration in all extender types caused an increase in post-thaw fertility values in comparison with those that not containing vitamin E (0 mM) (p < 0.05). In addition, higher values were obtained with the extender-II in terms of all vitamin E concentrations. Among the cryopreserved samples, the highest average fertilization rate (48.2% ± 1.2%) was obtained using the extender-II containing 1 mM vitamin E (Fig. 4). Statistical differences were significant among the treatments in terms of fertility of frozen–thawed sperm (p < 0.05).
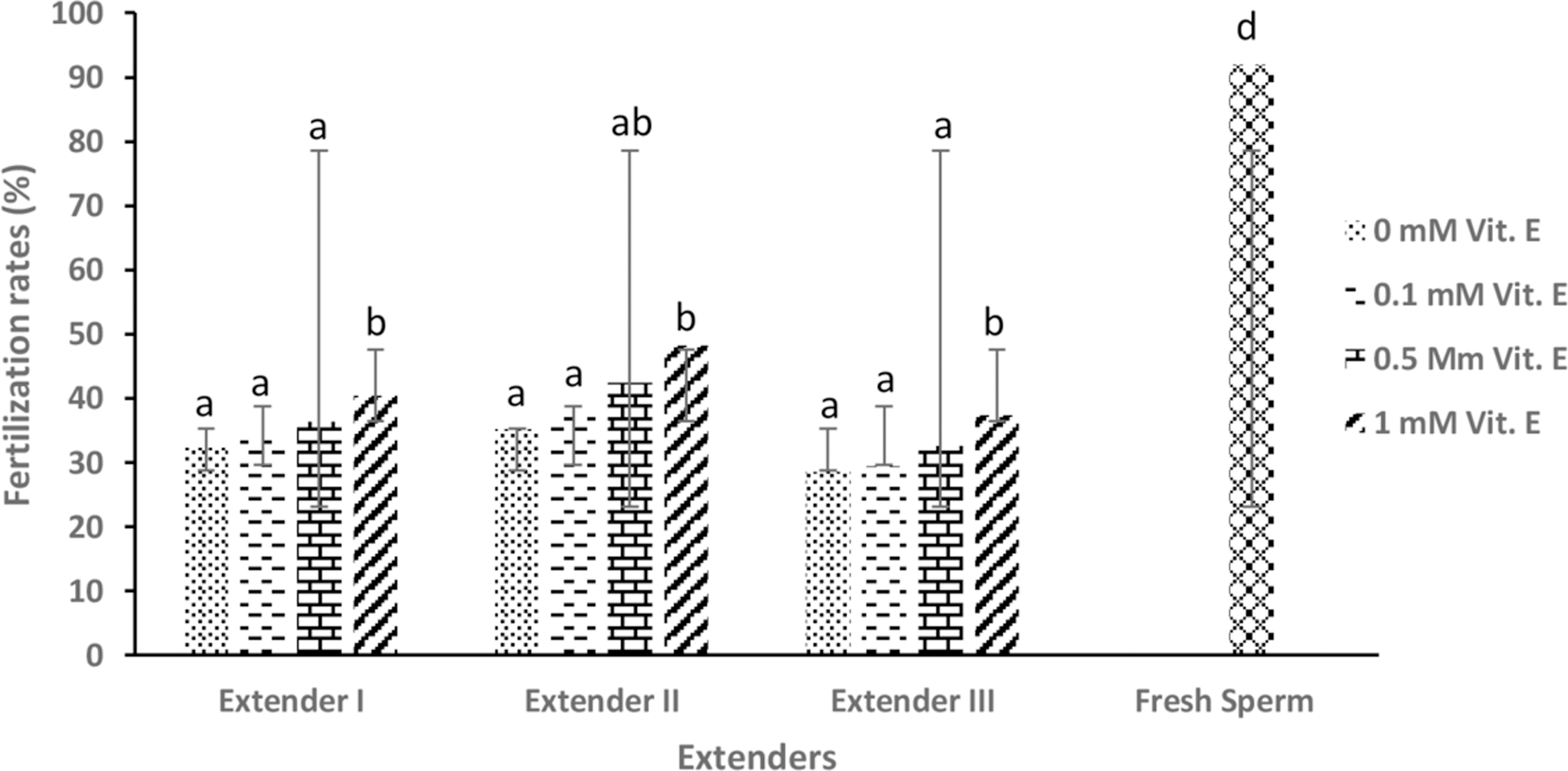
The mean post-thaw fertilization rates (%) of frozen–thawed brown trout (Salmo trutta macrostigma) sperm. Different letters indicate differences among treatments (ANOVA, p < 0.05, n = 9).
DNA damage
DNA damage of brown trout (Salmo trutta macrostigma) sperm cryopreserved with four different vitamin E concentrations and three different extenders are presented in Table 2. There were significant differences among the treatment groups (p < 0.05). Sperm cells frozen–thawed in the absence of vitamin E (0 mM) showed the highest DNA damage among all extenders. The extenders supplemented with vitamin E decreased the DNA damage. In the comet test, all extender groups containing vitamin E ranging from 0.1 to 1.0 mM produced lower DNA damages than that of the extenders not containing vitamin E (p < 0.05).
DNA Damage of Frozen–Thawed Brown Trout (Salmo trutta macrostigma) Sperm
Different letters indicate differences among treatments (ANOVA, p < 0.05, n = 9).
ANOVA, analysis of variance.
Discussion
Vitamin E (α-tocopherol) is a fat-soluble vitamin and also one of the most common antioxidants. Although the importance of vitamin E supplementation is addressed in many other studies, the majority of these studies have indicated the nutritional value of vitamin E supplementation. 23 However, several studies have reported its protective effect on sperm quality in domestic animals in recent years.24–26 On the contrary, studies about its protective effect on post-thaw sperm quality and DNA integrity are limited in aquatic species.
In this study, the effect of different extenders and vitamin E concentrations on post-thaw motility and fertilizing ability of cryopreserved brown trout sperm were compared. It was demonstrated that supplementation of the extenders with vitamin E increased the post-thaw motility, viability, duration, and fertilization rates depending on the extender type. In contrast, other researchers have shown that supplementation of vitamin E to the freezing medium caused a decrease in the motility rate in Pagrus major. 27 In addition, this vitamin did not affect the sperm motility rates of Prochilodus lineatus, 28 Sparus aurata, or Dicentrarchus labrax.7,29 These results differ from that of this study, suggesting that vitamin E is more effective for brown trout. These differences could be owing to the fish species or even to other conditions used such as different concentrations of vitamin E, cryoprotectants, and their interactions with the vitamin E extender.
It is well known that motility is a significant feature in evaluating fresh and post-thaw sperm quality. Consistent with our results, Ubilla and Valdebenito 30 found that post-thaw motility, fertilizing capability, and storage period increased when using vitamin C in extenders in rainbow trout sperm cryopreservation. According to Martinez-Paramo et al. 29 supplementation of cryosolutions with α-tocopherol and ascorbic acid improved sea bass post-thaw sperm motility. On the contrary, Cabrita et al. 7 indicated that post-thaw motility did not significantly increase with the addition of ascorbic acid and tocopherol to the cryosolution in gilthead seabream and European seabass sperm. The findings of this study indicated that there were significant improvements in the quality of frozen–thawed brown trout sperm resulting in higher fertilization rates in response to vitamin E additions to the extenders.
The findings of this study also revealed a lower motility of frozen–thawed brown trout sperm than fresh ones, which might be owing to a decrease in the ATP stores resulting in low mitochondrial activity, damage in cell membrane integrity, or serious cryogenic DNA damage occurring during the cryopreservation process. The assessment of the DNA situation of frozen–thawed sperm is particularly important in terms of commercial-scale usage or for cryobanking. After cryopreservation, DNA fragmentation has also been reported in rainbow trout, 31 seabream, 5 sea bass, 32 Nile tilapia, 33 and paddlefish. 34 The probable physiological reason for this situation may be the insufficient natural antioxidants in seminal plasma, which can promote the creation of free radicals. The positive effect of Vitamin E is that it deactivates the negative effects of ROS with a consequent improvement of the thawed semen quality.
Furthermore, a decline in TAS (total antioxidant potential) in frozen–thawed samples, which shows a decrease in antioxidants, naturally exists in the seminal plasma of gilthead seabream, as was reported by Martinez-Paramo et al. 35 This situation may have occurred because of reducing the capacity of sperm to withstand further cryodamage in the absence of antioxidants such as vitamin E in the cryosolution. The results of this study reveal decreasing DNA damage in cryopreserved brown trout sperm with an increasing concentration of vitamin E in the extenders used. This situation can be explained by the observed reduction in DNA damage, which may be owing to the decrease in ROS activity when vitamin E is added as an antioxidant.
The findings of this study are in agreement with the results of Massaeli et al. 36 who indicated that vitamin E was rather effective in the prevention of lipid peroxidation reactions in the cell membranes. In addition, according to some studies of vitamin E supplementation, it improved post-thawed sperm motility, viability, membrane integrity of goat, rooster,37,38 and human sperm DNA integrity, 24 which is in agreement with our findings.
Conclusion
In conclusion, the findings of this study demonstrated that supplementation of vitamin E to extenders significantly improved sperm quality in terms of motility, motility duration, viability, DNA damage, and fertility in brown trout. Overall, the addition of vitamin E to the freezing extenders is advisable to optimize brown trout sperm cryopreservation. However, further research is required to select the best concentration for this purpose.
Footnotes
Acknowledgments
The authors thank the staff of the Kılıç Bafa Aquaculture Farm.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was funded by İskenderun Technical University Scientific Research Fund (Project No: ISTE BAP-2017-YP-04).