
Abstract
Oocyte vitrification, as a vital step in reproductive medicine, is strongly associated with lower development caused by cryodamaging factors, such as oxidative stress. In this study, we evaluated the antioxidative synergistic effects of Melatonin (Mel) and Resveratrol (RES) coencapsulated by solid lipid nanocarriers (SLNs) against the pure antioxidant combination (Mel+RES). In this research, the formation of Mel+RES-SLN was confirmed by Fourier-transformed infrared spectroscopy. The average mean diameter, size distribution, polydispersity index, and zeta potential of particles were measured by Zetasizer, and the morphology was evaluated by scanning electron microscopy. In addition, the encapsulation efficiency (EE%) or drug loading capacity (DL%) of the nanocapsule was determined by spectrophotometric methods. Germinal vesicle (GV)-stage oocytes harvested from 6- to 12-week-old female NMRI mice were randomly divided into seven groups for in vitro studies. In these groups, (0, 10–12 M + 0.5 μM, 10−9 M + 2 μM, or 10−6 M + 10 μM) of Mel+RES/Mel+RES-SLN were added into vitrification media. After thawing, oocytes were matured, fertilized, and cultured for 3 days. Extra/intracellular reactive oxygen species (ROS) levels were measured in in vitro maturation medium after 24 hours. Our results revealed a significant improvement in the normal morphology of warmed GV-stage oocytes, GV breakdown (GVBD) rate, Metaphase II (MII)-stage oocyte formation, fertilization rate, early embryo development, and a significant reduction in intra/extracellular ROS level when vitrification media was supplemented with the lowest Mel+RES-SLN concentration. In vitro studies also demonstrated that the highest concentration of Mel+RES-SLN was safe, without a detrimental effect on embryonic development upon treatment. In conclusion, the lowest concentration of Mel+RES-SLN supplementation in GV-stage oocyte vitrification media improved maturation, fertilization, and embryo development rate and decreased extra/intracellular ROS level through an enhanced/controlled intracellular penetration compared to the pure Mel+RES.
Introduction
Establishing oocyte banks is a vital approach in assisted reproductive technique (ART) centers, which provides a major substitute for the preservation of fertility in patients who may lose their ovarian function due to failed infertility care, radiotherapy, chemotherapy, ovarian hyperstimulation syndrome, surgery, and sperm inaccessibility on the day of oocyte recovery. 1 After the germinal vesicle (GV) stage oocyte chromosomes reside within the nucleus membrane, and the meiotic spindles have not been observed. 2 The vitrification of immature oocytes in the GV step is recommended instead of mature oocytes (MII).2,3 Despite many recent advances in this technique, the rates of viability, fertilization, and developmental competence are considerably lower.4,5 For example, the blastocyst rate of the mouse vitrified-warmed GV oocyte followed by in vitro maturation (IVM) was 17%–42.9%. 5 These decreased rates may lead to severe deleterious effects such as lipid peroxidation, DNA damage, and protein oxidation, due to impaired redox activity and raised reactive oxygen species (ROS) levels. 6 Nevertheless, the use of exogenous antioxidants (alone or a combination of several antioxidants) during the vitrification system is necessary and reasonable.7–12
Melatonin (N-acetyl-5-methoxytryptamine, Mel) is a bioactive molecule generated from the important amino acid tryptophan and was first detected in the mammalian pineal gland. 13 The evidence strongly suggests that Mel is an effective free radical scavenger13–16 and promotes preimplantation of the mouse oocyte/embryo.17,18 In addition, Mel was active in having a significant role in the production of vitrified-warm oocytes, potentially leading to better IVM. 19 It was recently reported that 10−9 mol/L Mel could improve the blastocyst development rate of vitrified mouse MII oocytes after parthenogenetic activation 20 and increase the maturation rate of the vitrified-warmed mouse GV-stage oocytes. 21
Resveratrol (3,5,4′-trihydroxystilbene, RES) is well established for its natural antioxidant effects due to free radical scavenging and its action on ROS-dependent pathways. 22 Numerous studies have also been performed on the antioxidant activity of RES in oocytes.23–25 In addition, RES supplementation in IVM phases or vitrification/warming procedures is reported to enhance the resistance of oocytes against cryopreservation injuries and improve the developmental potential of oocytes.26,27 However, there is little information about the effects of RES on the embryonic developmental efficiency of mouse oocytes after vitrification. The results of Lee et al. suggest that the combination of RES and Mel supported a synergistic increase in oocyte nuclear maturation, total blastocyst cell count, and development of SCNT embryos. 5
Although these features have prompted us to investigate that, if the combination of RES and Mel may be used as an enticing synergy through GV-oocyte vitrification, there are still some major shortfalls (such as poor and variable bioavailability, minimal biolife, fast absorption and metabolism, and low water solubility). This approach is likely to restrict the synergistic function of the exogenous application of Mel and RES as antioxidants and antiapoptotic molecules in vitrification. One of the most successful approaches to solve these difficulties is the use of nanodrug coencapsulation.28–30 Around the same time, two or more drugs may be transported to the intercellular space by a single carrier by coencapsulation. This results in “synchronizing” intraoocyte passage and biodispersal, overcoming numerous biological barriers to the ultimate optimal synergistic combination at the intracellular level. 31 Various carriers, for example, liposomes, 32 micelles, 33 and solid lipid core nanocapsules (SLNs), 34 have been used for the physical loading of drugs. Among them, self-assembled nanoparticles such as SLNs are at the forefront of the rapidly developing field of coencapsulation technology, owing to the unique strength of structure and composition. 35 SLNs are more efficient at coencapsulating various bioactive molecules with a controllable and flexible loading ratio than traditional nanoparticle formulations. 36 Finally, this progress led us to the approach of codelivery of RES and Mel by coencapsulation in SLN.
Despite recent advances in nanotechnology and its contribution to the development of new cancer treatments, 37 the use of new pharmaceutical approaches such as coencapsulation in reproductive medicine is still in its infancy. So, to the best of our knowledge, there is no study on the effects of Mel or RES loaded SLN (along or a combination) on oocyte vitrification. However, we found in our previous works that they have improved the deleterious effects of antioxidant on the mouse oocyte/embryo (unpublished). Thus, in this research, we investigated the cryoprotective effects of Mel+RES or Mel+RES loaded SLN (various concentrations) supplementation of vitrification media on the survival, maturation, fertilization rate, and embryo development to the compact-morula stage of warmed/GV-stage oocytes. Considering the ethical concerns relevant to the collection of human oocytes for in vitro fertilization (IVF) studies, we performed research on mice oocytes as a model.
Materials and Methods
Chemicals
Stearic acid, Polysorbate 80 (Tween 80), sodium tripolyphosphate solution (TPP), CH2CL2, KSOM-aa medium, Mel, and RES were purchased from Sigma–Aldrich (St. Louis, MO). Pregnant mare serum gonadotropin (PMSG) and bovine serum albumin (BSA contained 0.8% fraction V) were obtained from Biochrom (Berlin, Germany). G-MOPS and G-IVF media were purchased from Vitro life. 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) stain, Ham's F10, and mineral oil were obtained from GIBCO Invitrogen (Grand Island, NY). Buffer ingredients and all other chemicals or solvents were from Merck (Germany).
Animals
Six- to twelve-week-old NMRI female mice were purchased from the Iranian Pasteur Institute. Mice were kept in a standard room with light and temperature (12-h light/12-h dark cycles, 22 ± 2°C). Our institution's ethics committee approved all protocols regarding animals (IR.KUMS.REC.1396.665). This study was funded by the National Institute for Medical Research Development (NIMAD, 976960). Our research is supported by Kermanshah University of Medical Sciences, which is primarily involved in education or research.
Preparation of Mel+RES-loaded solid lipid nanocapsule (Mel+RES-SLN)
Melatonin and Resveratrol that co-encapsulated by SLN (Mel+RES-SLN) was prepared by a general self-assembly method. 38 Briefly, 10 mg of stearate acid and a similar amount of RES and Mel dissolved in CH2CL2 (to get the 6 mg) were added to an aqueous solution containing 0.25 mg of the Tween 80 and distilled water solution under stirring. The mixture was sonicated for 15 minutes at 80 W. The emulsion was centrifuged (18,200 g, 25 minutes; Eppendorf, 5415D, Germany) at room temperature and filtered to eliminate the unloaded RES and Mel using a 0.2 μm filter membrane and then freeze-dried and stored at −20°C for further assay. Unloaded SLN was prepared using the same procedure. The nanocapsules were tested for alterations in desired quality attributes when added to oocyte culture media.
Characterization of prepared Mel+RES-SLN
Fourier-transformed infrared spectroscopy
The chemical composition of the synthesized nanocapsules was determined using FTIR spectroscopy (model FTIR Prestige-21; Shimadzu Co., Japan). The sample spectra were recorded over the 200–4000 cm−1 scanning range, with a 4 cm−1 spectral resolution.
Basic physicochemical properties
The average particle diameter (size), size distribution, polydispersity index (PDI), and zeta potential (ZP) of the nanocapsule (Mel+RES-SLN) were determined using a Zetasizer instrument (model 3600; Malvern Instruments Ltd., Worcestershire, UK) at 25°C. Morphology and surface characteristics were evaluated with scanning electron microscopy (SEM) (KYKY-EM3200).
Drug loading and encapsulation efficiency
UV-vis spectrophotometry (Philips PU 8620) was used by our research team to evaluate entrapment efficiency (EE%) or drug loading content (DL%) as previously described 39 against a prepared calibration curve.
EE% and DL% of Mel and RES (drugs) were calculated using the following equation:
In vitro release profile
The in vitro release profiles of the Mel+RES-SLN solution were investigated by the dialysis bag method. 38 Briefly, the Mel+RES-SLN and Mel+RES (comparable to 1 mg of each one) were redispersed in 80 mL of release medium, that is, phosphate-buffered saline (PBS; pH 7.4) for the 55 hours at 37°C ± 0.5°C in the dialysis bags. At different time intervals (0.5, 1, 2, 3, 4, 5, 6, 24, 48, and 55 hours), 1 mL of the receptor phase was taken out and replaced with the same volume of fresh fluid. The quantity of Mel or RES in each sample release medium was evaluated by a UV spectrophotometer set (Philips PU 8620) at 280 nm.
Oocyte collection
Superovulation was performed intraperitoneally by injection of 10 IU serum gonadotropin (PMSG; 80056-608; VWR Science, Inc.) from a pregnant mare. Cervical dislocation of mice was performed 48 hours after the PMSG injection. Ovaries were then collected and transferred to G-MOPS media. They were washed away and destroyed the connective tissue around them. Ovaries were searched under a stereomicroscope, and the GV-stage oocytes were selected from them by a slicing method 40 under a stereomicroscope (Nikon Corporation, Japan).
Vitrification and thawing
For vitrification, oocytes were converted into an equilibrium solution comprising 7.5% dimethyl sulfoxide and 7.5% ethylene glycol in Ham's F10 medium, supplemented by 20% human serum albumin (HSA) for 10 minutes. Oocytes were then placed into a vitrification solution (Ham's F10 medium supplemented with 20% HSA, 15% ethylene glycol, 15% dimethyl sulfoxide, and 0.5 M sucrose) for 45 to 60 seconds. Finally, the oocytes were loaded onto Cryotops (Kitazato BioPharma Co., Japan) and transferred to liquid nitrogen (LN2) immediately, then stored for 40 days. 41
For warming, the Cryotops were put into a warming solution (Ham's F10 supplemented with 20% HSA and 1 M sucrose) for 60 seconds. The oocytes were put in Dilution Solution-1 (Ham's F10 supplemented with 20% HSA and 0.5 M sucrose) for 3 to 5 minutes after release from the Cryotops and finally located in Dilution Solution-2 (Ham's F10 supplemented with 20% HSA and 0.5 M sucrose). The oocytes were then put in a washing solution (Ham's F + 10% ∼ 20% HSA), rinsed thrice, and incubated in the G-IVF medium for 1 hour.
After warming, oocytes were put into IVM medium (α-MEM [alpha minimum essential medium]) for 24 hours. The oocyte's nuclear maturity steps (viability GV, GVBD, and MII) were evaluated in each group.
Sperm preparation and IVF
The cauda epididymis (of NMRI male mice at 8–12 weeks) was cut and placed into 500 μL of Hams F10 medium containing 10 mg/mL (BSA). After 30 minutes of incubation (for sperm capacitation) and microscopic sperm analysis, 1 × 106/mL sperms were transferred into each 100 μL droplets of preincubated G-IVF medium containing MII oocytes. The oocytes were assessed for the presence of female and male pronuclei after 6 hours of coincubation with sperm. The presumptive zygotes were transferred into preincubated KSOM medium, and 2-, 4-, and ≥8-cell stages (until day 3) of embryo development and embryo fragmentation were counted in each group and compared. 40
Measurement of intracellular ROS levels
The fluorescent probe, H2DCFDA stain, was used to evaluate intracellular ROS production. After IVM, 25 oocytes from each group were stained according to the modified protocol previously described. 42 Finally, under the Eclipse Ti epifluorescence microscope (Nikon Instruments, Inc., Melville, NY) all samples were visualized along with ROS filters using 450–490 and 515–565 nm excitation and emission wavelengths, respectively. For each experiment, the treatment was conducted five times, with 5–7 embryos. For each sample, the replication was conducted five times, with 5–7 embryos.
Measurement of extracellular ROS levels in culture medium
After evaluating the oocyte maturation, the maturation media without any warmed oocyte of each group and basic media (as the control medium) were collected. The levels of extracellular ROS were measured by a chemiluminescence assay using a luminol probe (5-amino-2, 3-dihydro-1, 4-phthalazinedione; Sigma, St. Louis), as mentioned previously, 43 and results were displayed in relative light units per sec. 44
Experimental design
This experiment was designed to compare the effect of Mel+RES (free combination) and Mel+RES-SLN (coencapsulated combination by SLN) added to vitrification media. The concentrations of RES (2 mM) 26 and Mel (1 nM)20,21 were set according to previous studies, and we designed seven groups: In the first group (control), nothing was supplemented to the vitrification/thawing medium; in second to seventh groups, the GV-stage oocytes were frozen in the basic vitrification medium plus different concentrations of Mel+RES (10−12 M + 0.5 μM, 10−9 M + 2 μM, or 10−6 M + 10 μM) and Mel+RES-SLN (10−12 M + 0.5 μM, 10−9 M + 2 μM, or 10−6 M + 10 μM). After 48 hours, warmed GV-stage oocytes were matured, fertilized, and cultured as mentioned above. For each treatment, the GV breakdown (GVBD) and MII-stage oocytes, subsequent embryo development and the intra/extracellular ROS level were determined as stated above.
Statistical analyses
All statistical analyses were made using the Social Sciences 20 Statistical Package (SPSS, Inc., Chicago, IL). With the Kolmogorov–Smirnov study, data normalization was verified. The information was evaluated using the one-way analysis of variance method and the post-test Tukey. All data were expressed as mean ± standard deviation. p < 0.05 was considered significant.
Results
Physicochemical characterization of Mel+RES-SLN
Table 1 summarizes the physicochemical characterization of the Mel+RES-SLN. The hydrodynamic sizes of the Mel+RES-SLN using dynamic light scattering showed an average particle size ≤200 nm and PDI ≤0.2 (Table 1 and Fig. 1), which were confirmed by SEM images (Fig. 2). The ZP analysis of Mel+RES-SLN was −23 mV (Fig. 1). Our result showed the highest EE% with values of 89% and ≥93% for Mel and RES from Mel+RES-SLN, respectively (Table 1). The DL of Mel and RES in the Mel+RES-SLN is 21% and 23%, respectively.

Physicochemical characterization of Mel+RES-SLN by Zetasizer instrument; the particle size distribution and zeta potential. Mel, melatonin; RES, resveratrol.
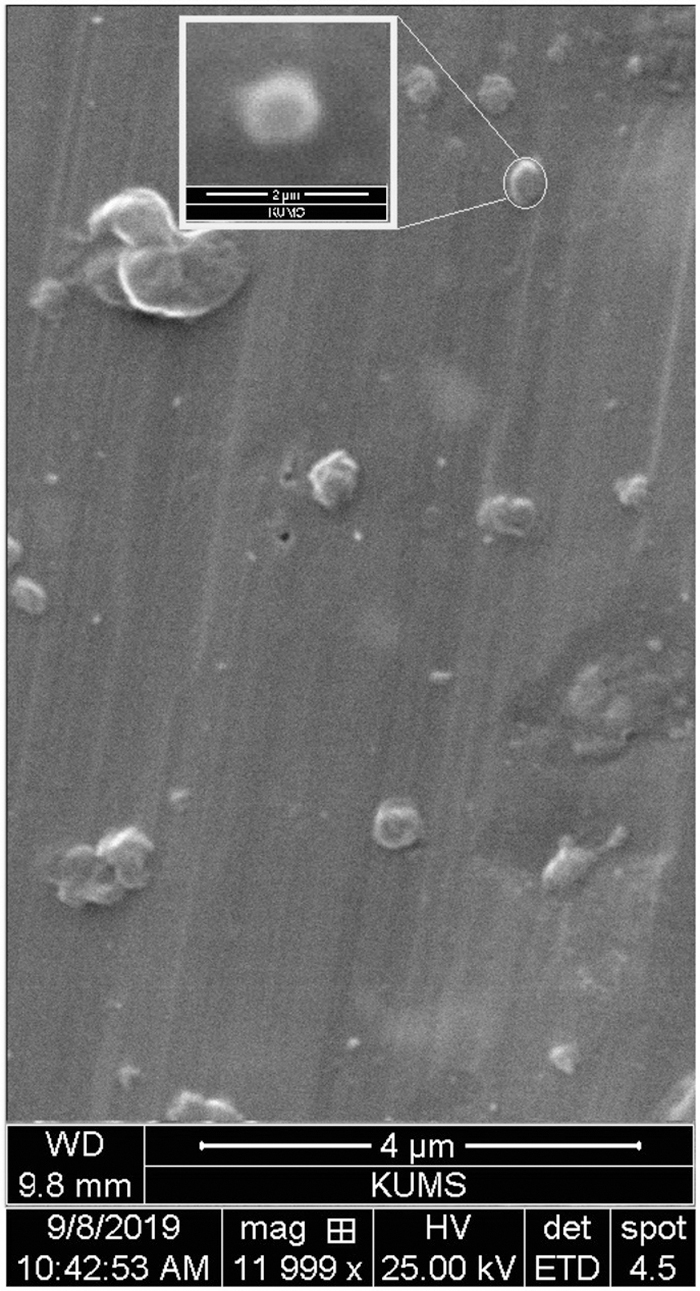
SEM image: morphology analysis of Mel+RES-SLN. SEM, scanning electron microscopy.
Physicochemical Features of the Different Nanocapsules
Data are presented as absolute values (mean ± SD).
The experiment was repeated five times.
B-SLN, “blank” solid lipid nanocapsules; DL%, drug loading capacity; EE%, encapsulation efficiency %; Mel, melatonin; Mel+RES-SLN, SLN, entrapping Mel and RES; PDI, polydispersity index; RES, resveratrol; SD, standard deviation.
FT-IR analysis
The FTIR spectra of free antioxidants (Mel or RES), B-SLN, and Mel+RES-SLN complex are presented in Figure 3. The spectrum of Mel+RES-SLN (Fig. 3a) revealed sharp bands at 2918 and 2873 cm−1, 1155, 1082, and 1033 cm−1, 3439–3400, 1618 cm−1, and the broad peak at 2500–3600 cm−1. In addition, the SLN spectrum (Fig. 3b) presented a sharp peak at 1693 cm−1, 2500–3600 cm−1, 2862–2927 cm−1, 1604 cm−1, the broad peak at 3600–2500 cm−1, and a weak peak at 3012 cm−1. In the Fourier-transformed infrared spectroscopy (FT-IR) spectra of RES (Fig. 3c), sharp peaks at 1587 and 1460 cm−1, 1627 cm−1, and 3408 cm−1 and a weak peak at 2859 cm−1 were observed. The FT-IR spectrum of the prepared Mel (Fig. 3d) indicated three major peaks at 3333 (N–H stretching), 1625 (C = O stretching), and 1210 (C–N bending). 45

FT-IR diagrams of the SLNs: Mel+RES-SLN; B-SLN; Mel; RES. FT-IR, Fourier-transformed infrared spectroscopy. Color images are available online.
In vitro drug release
The in vitro release profile of Mel and RES from the pure antioxidant combination and Mel+RES-SLN was tested by the dialysis method in PBS (pH 7.4 at 37°C) (Fig. 4). It can be seen in Figure 4 that Mel or RES absolute release percentage from Mel+RES-SLN was lower compared with Mel+RES (Fig. 4). In particular, Mel+RES-SLN released only <38% and 25% of the RES at 60 hours cumulatively, respectively. Meanwhile, Mel and RES were 100% released from the pure antioxidant combination (Mel+RES) during the first 20–25 hours.
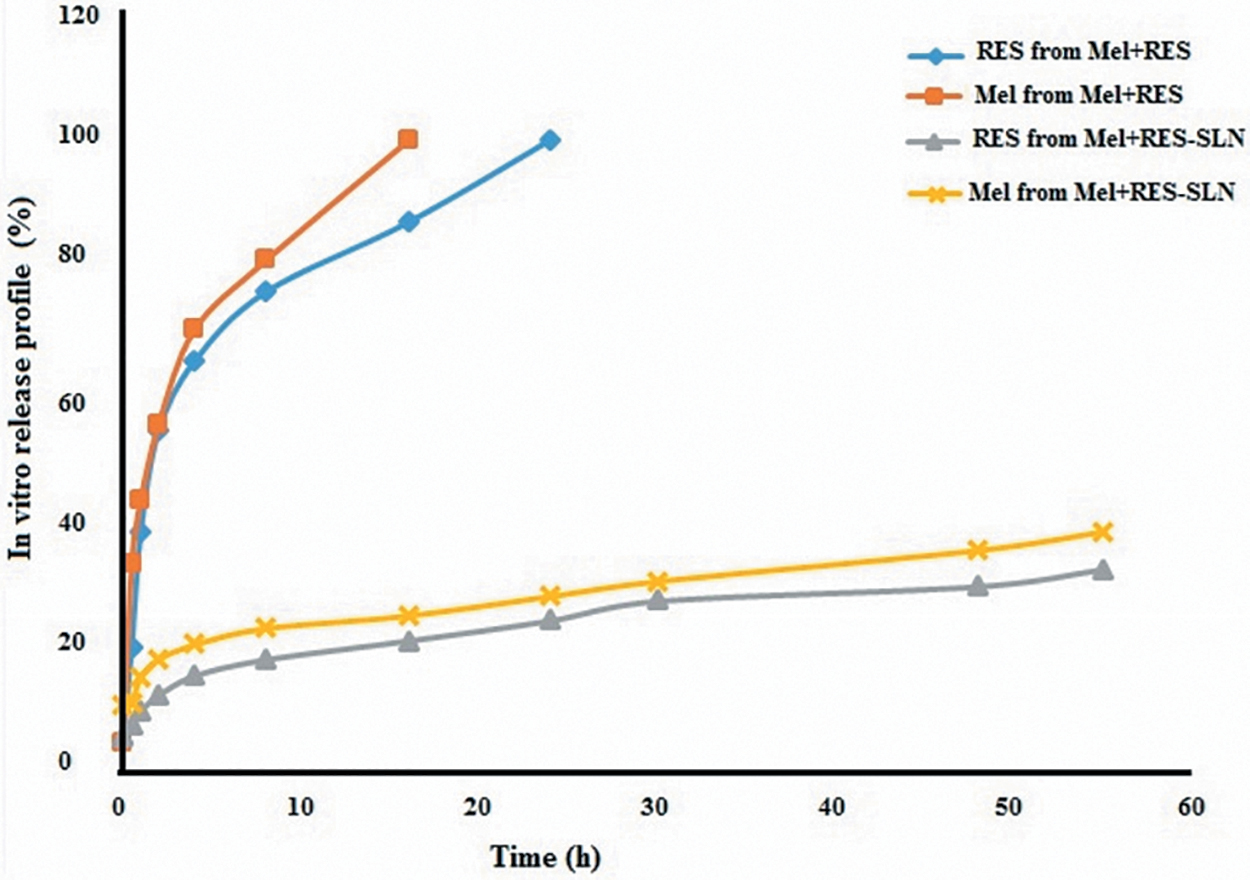
In vitro drug release profiles of Mel and RES from Mel+RES-SLN. Color images are available online.
Effects of Mel+RES-SLN and Mel+RES on MII-stage oocyte and embryo development
As shown in Table 2, supplementation of vitrification media with Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) and the medium concentration (10−9 M + 2 μM) improved significantly the normal morphology of warmed GV-stage oocytes compared with the control group (94.40% ± 1.15% and 77.46% ± 1.14% vs. 71.32% ± 1.93%; p < 0.05, Table 2). Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) produced the highest GVBD (90.37% ± 2.77%; p < 0.05) and nuclear maturation (MII) rate (86.66% ± 3.24%; p < 0.05) in comparison to other treatments (Table 2). In addition, the normal morphology of warmed GV-stage oocyte rate was significantly increased in the presence of pure Mel+RES at the medium concentration (10−9 M + 2 μM) treatment (p < 0.05) compared to the control (76.29% ± 1.69% vs. 71.32% ± 1.93%, Table 2). The rates of GVBD and MII in the pure Mel+RES (10−9 M + 2 μM) group were significantly higher than the control group (76.15% ± 1.62% and 64.23% ± 3.11% vs. 71.56% ± 2.18% and 56.86 ± 1.14; p < 0.05, respectively). However, there was no significant difference between the highest concentration of Mel+RES-SLN (10−6 M + 10 μM) and the control group regarding the normal morphology of warmed GV-stage oocytes, GVBD, and MII rates (p < 0.05). As indicated in Figure 5, RES+Tertinoin (TTN)-SLN at the lowest concentration (10−12 M + 0.5 μM) significantly increased fertilized oocytes, cleavage, and 4–8 and 8–16 cell-embryo (compact morula) formation rates and significantly decreased >10% fragmentation rate compared to control and pure Mel+RES at the medium concentration (10−9 M + 2 μM) (p < 0.05, Fig. 5).

In vitro early embryo development (a) fertilization rate; (b) cleavage rate; (c) 4-cell rate; (d) compact morula rate; and (e) ≥fragmentation rate.
The experiment was repeated five times.
Data are presented as absolute values (percentages ± SD), and different superscript letters (a–e) indicate significant differences among experimental groups (p < 0.05).
GV, germinal vesicle; GVBD, germinal vesicle breakdown; MII, metaphase of the second meiotic division.
However, vitrification medium supplementation with the highest concentration of the pure Mel+RES (10−6 M + 10 μM) decreased the fertilization rate and embryo development into 2, 4–8, and >8-cell stages, as well as an increased embryo with >10% fragmentation, (p < 0.05) (Table 2 and Fig. 5). But coencapsulation of the highest concentration of Mel+RES (10−6 M + 10 μM) by SLN had no significant toxic effects on oocyte/embryo development after warming.
Evaluation of Mel+RES-SLN and Mel+RES added to vitrification medium on intracellular ROS level
The GV oocyte that was treated with Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) had a significantly lower intracellular ROS level compared to all the other groups (Fig. 6). In addition, intracellular ROS levels were decreased in Mel+RES (10−9 M + 2 μM) groups compared to the control group (Fig. 6).

Evaluation of intracellular ROS levels in mouse GV-stage oocyte stained using DCHFDA; ROS, green. Bar, 50 μm. DCHFDA, 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA); GV, germinal vesicle; ROS, reactive oxygen species. Color images are available online.
Evaluation of Mel+RES-SLN and Mel+RES added to vitrification medium on extracellular ROS level in the culture media
As shown in Figure 7, extracellular ROS levels in maturation medium containing oocytes treated for 24 hours with Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) were significantly lower than all the other groups (p ≤ 0.05). There was a significant difference between Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) and Mel+RES at (10−9 M + 2 μM) (Fig. 7), but both had significantly decreased extracellular ROS levels compared with the control. There was no significant difference between the highest concentration of Mel+RES-SLN (10−6 M + 10 μM) group and control (Fig. 7). However, the highest concentration of Mel+RES (10−6 M + 10 μM) increased the extracellular levels of ROS significantly (p < 0.05, Fig. 7).

Effects of Mel+RES-SLN on extracellular ROS levels in vitrified/warmed GV-stage oocyte maturation medium. Different superscript letters (a–d) indicate significant differences among experimental groups (p < 0.05).
Discussion
Although GV oocyte vitrification is a key strategy for infertility treatment in patients with cancer (e.g., oocyte banking and donation), 46 there is a concern all over the world regarding cryodamage to vitrified GV-stage oocytes. 21 Despite this, antioxidant effectiveness was confirmed during oocyte vitrification, against the cryodamaging effect that led to ROS.7–12 Yet their usage has several significant drawbacks, such as poor and variable bioavailability, minimal biolife, and low water solubility. One of the most successful approaches to solve these difficulties is the usage of nanodrug encapsulation.28–30 The current study is the first to utilize the solid lipid co-nanocapsules (SLN carries two drugs) to deliver optimal doses of Mel and RES and evaluate their synergistic antioxidant efficacy during mouse GV-stage oocyte vitrification compared to the pure antioxidant combination (Mel+RES).
In this research, we supplemented vitrification solutions with various concentrations of Mel+RES-SLN or Mel+RES. Our findings showed that Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) not only was able to significantly enhance normal morphology, polar body extrusion rate (nuclear oocyte maturation), fertilization rate, and vitrified-thawed GV oocyte but also synergistically decreased intra/extracellular ROS levels better than Mel+RES. These data suggest that SLN delivers adequate amounts of both (Mel and RES antioxidants) to GV-stage oocytes more efficiently than the pure combination. 47 These beneficial effects of Mel+RES-SLN are most likely to be controlled release dependent.
The synthesized Mel+RES-SLN was confirmed by FTIR analysis with a mean particle size of ≤200 nm, as mentioned in the result section. This synthetic system could facilitate the passage across biological barriers and provide better integration for delivery of RES/Mel into the subcellular compartments such as mitochondria and nuclei. 36 Moreover, this particle's size is in the optimal range for accumulation in oocytes and embryos, owing to the enhanced permeation and retention effects. 48 In addition, the surface charge (−23 mV), a slight size distribution (PDI <0.2), spherical shape, normal distribution without aggregation (Figs. 1–4), and high encapsulation efficiency (EE% ≥ 89%) are other features of synthesized Mel+RES-SLN. All of these nanocapsule's basic physicochemical properties are likely reasons for its higher cellular uptake of Mel+RES-SLN compared with pure form (Mel+RES groups); (Fig. 5). 49
The vitrified/warmed oocytes during vitrification are extraordinarily sensitive to intra/extracellular ROS, which has played a significant role in the poor developmental potential. 11 Numerous studies have shown that intracellular ROS levels are a key mediator of the intrinsic oxidative stress pathway, which increases DNA breakdown/or cellular damage. 50 The extracellular ROS level may also be used as an indicator of slow development (≥8 cell embryo on day 3), ≥10 fragmentation, and reduced formation of compact morula. 51 Our H2DCFDA and chemiluminescence results showed that the Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) was more efficient than Mel+RES group to scavenge excessive production of intra/extracellular ROS during vitrification, which provided another explanation for improved oocyte/embryo development rates by this system (Figs. 6 and 7).
In addition, Mel+RES-SLN is safe, with up to 5 + 10 μM tested, and no cytotoxicity is associated with its supplementation in in vitro culture and IVM media observed. This is likely correlated with in vitro controlled antioxidant release owing to the fit of amphiphilic balance in Mel+RES-SLN nanoscopic structures. Only <30% of RES and Mel cumulatively were released from Mel+RES-SLN complex in 60 hours, while ∼100% of RES and Mel were released from RES+Mel over 60 hours, due to the high stability of Mel+RES-SLN with controlled leakage of antioxidants after the intracellular operation.
However, our results may provide a powerful potential application of SLN technology for other antioxidants, hormones, and molecules that are poorly soluble in water for better utility in the field of cryopreservation. However, the cell's internalization mechanisms of Mel+RES-SLN are still only partly understood. In addition, the limitation of this study is the lack of further development of oocytes by blastocyst formation and embryo transfer to the uterus. Therefore, future work must be performed on other analyses or in vivo studies before its application on human ARTs.
Conclusions
Considering all of the above, Mel+RES-SLN at the lowest concentration (10−12 M + 0.5 μM) can provide a safe synergistic antioxidative effect to inhibit intra/extracellular ROS production through an enhanced intracellular penetration of Mel or RES compared with Mel+RES (10−9 M + 2 μM). This is due to a particle size ≤200 nm, PDI ≤0.2, positive ZP, higher EE% or DL%, a monodisperse spherical shape, and the slow/simultaneous release of Mel and RES from Mel+RES-SLN compared with Mel+RES. In contrast, the present work confirms a better efficacy of dual delivery SLN than free antioxidant combination during cryopreservation of mice GV oocytes. However, future investigations are warranted to clarify the exact molecular and cellular processes of Mel+RES-SLN treatment during oocyte/embryo vitrification.
Ethical Approval
All applicable institutional guidelines for the care and use of animals were followed (IR.KUMS.REC.1396.665).
Footnotes
Acknowledgments
The writers want to thank the coworkers who helped with the tests and the collection of data. F.A., A.V.-R., M.K., and E.A. are all from Kermanshah University of Medical Sciences, where research and education are the primary functions.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by the National Institute for Medical Research Development (NIMAD, 976960). Our research is supported by Kermanshah University of Medical Sciences, which is primarily involved in education or research.