
Abstract
This study investigated the effect of sample storage duration on the quantification of oxidative stress markers in the gastrocnemius, heart, and brain of mice submitted to a maximum swimming exercise. Thiobarbituric acid reactive substances (TBARSs), protein carbonyl derivatives, total antioxidant capacity (TAC), and the activity of superoxide dismutase (SOD) and catalase (CAT) were quantified in fresh tissues and in samples stored at −80°C for 1, 3, or 6 months, from exercised (n = 13) and nonexercised mice (n = 13). Except for protein carbonyl derivatives in the heart, the exercise resulted in the modification of all markers in all fresh-evaluated samples (p < 0.001). The storage duration did not modify the effect of exercise on protein carbonyl derivatives and TAC. TBARS was stable for 3 months in the gastrocnemius and for 1 month in frozen heart and brain. Accordingly, the exercise effect on TBARS levels observed in fresh samples was absent in the gastrocnemius frozen for 6 months (p = 0.98) and in the heart and brain frozen for 3 months (p = 0.07 and 0.28, respectively
(Color images are available online).
Introduction
Exercise of sufficient volume and intensity can lead to increased reactive oxygen and nitrogen species (RONS) production in the contracting skeletal muscle and in other organs and tissues.1–3 As RONS are short-lived compounds, their direct measurement is generally not feasible. Thus, indirect indicators, such as oxidative-modified lipids and proteins, are frequently quantified. 4 In this context, many studies show increased levels of oxidative stress biomarkers, such as thiobarbituric acid reactive substances (TBARSs) and protein carbonyl derivatives, after an acute bout of exhaustive exercise, in skeletal muscle, heart, brain, and liver, among others.5–8 RONS produced during exercise are involved in the activation of redox-sensitive signaling pathways that mediate some of the skeletal muscle adaptive responses to exercise.9–11 However, excessive RONS production can result in oxidative stress, which is involved in reduced muscle force production and fatigue during exercise. 12 RONS effects are controlled by antioxidant mechanisms, including the enzymes superoxide dismutase (SOD) and catalase (CAT), as well as nonenzymatic antioxidant compounds. 13 Upregulation of the antioxidant mechanisms is one of the most frequently observed adaptive responses to exercise training,6,11,14 which increases the ability to control RONS oxidative effects.
To evaluate how exercise affects tissue redox status, reliable measurement of oxidative stress biomarkers is mandatory. Biomarker quantification is strongly affected by preanalytical sample handling and storage procedures.15–17 Most often, in biological assays, tissue samples are frozen at ultralow temperatures (−80°C to −160°C) before the quantification of the biomarker of interest. Although it has already been shown that the stability of different biomarkers can be jeopardized by long-term storage,17–20 most studies generally do not provide information about the sample storage duration until the analysis. There is also limited information about the effect of storage duration on the stability of oxidative stress biomarkers. It has been demonstrated that SOD and CAT activity in the kidney are reduced after 28 days of storage at −70°C, although this procedure did not affect the SOD activity in the liver. 20 However, SOD activity was reduced in liver samples stored at −80°C for 60 days, 18 which suggests that the effects of storage duration on the stability of antioxidant enzymes are both tissue and enzyme specific. However, to our knowledge, no study has been conducted to date to investigate whether the stability of biomarkers during sample storage can be affected by different physiological conditions.
In this study we investigated whether sample storage duration can alter the quantification of oxidative stress markers in different tissues from mice submitted to a maximal swimming protocol. The markers of interest were quantified in fresh excised tissues and in samples stored at −80°C for 1, 3, or 6 months, from both exercised and nonexercised mice. We hypothesized that the longer the tissue storage duration, the greater the interference with the evaluation of the exercise effect on the oxidative stress markers. We also proposed that the effect of storage duration would be tissue and marker specific.
Methods
Animals and exercise protocol
Swiss male mice (n = 26), 8 weeks old, were housed in a temperature-controlled room (22.0°C ± 2.0°C) with a 12-h light/12-h dark photoperiod. Filtered water and standard rodent chow were supplied ad libitum. The animals were handled in accordance with the principles of Law 11.794 of October 8 of 2008 that established the protection of animals used for experimental and other scientific purposes in Brazil. All procedures performed in the present study were in accordance with the Ethics Committee on the Use of Animals of Universidade Federal dos Vales do Jequitinhonha e Mucuri (CEUA/UFVJM, protocol 042/2015).
The animals were randomly distributed into two groups: exercised (EX, n = 13, body mass = 39.0 ± 2.2 g) and nonexercised control (CON, n = 13, body mass = 38.9 ± 2.7 g) mice. Animals were maintained in separate cages (820 cm2), four animals per cage. For adaptation to the aquatic environment, mice from both groups swam during five consecutive days (water temperature = 32.0°C ± 1.0°C), 10 minutes/day (between 8 and 9 a.m.), with no workload.
Two days after the adaptation period, mice from the EX group were submitted to a maximum progressive swim exercise, as previously described. 21 Briefly, animals swam until exhaustion, with an initial workload of 2% body mass, attached as a metal hook to the mouse's tail. Workload was increased by 2% every 10 minutes, without exercise interruption, by the addition of a metal ring to the metal hook attached to the animal's tail. Exhaustion was defined as the point at which the mouse remained below the water surface for more than 4 seconds. Time to exhaustion and maximum workload achieved were registered for each animal. Control mice did not perform any exercise but were kept immersed in water, at the level of the scapular region, for 30 minutes.
Animals were euthanized by cervical dislocation immediately after either maximum exercise or water immersion, and the tissues (gastrocnemius muscle, heart, and brain) were quickly excised and kept on ice bath for processing.
Tissue fragmentation and processing
After being removed, the brain was quickly washed in ice-cold phosphate buffered saline (PBS, 0.15 M, pH 7.4), and the cerebellum and the brainstem were excised. The hemispheres were separated from the olfactory bulb, and a sagittal cut was made in each one. Thus, the brain was divided into four fragments with a scalpel, to include the same structures (isocortex, including the somatomotor areas, thalamus regions, hypothalamus, and hippocampus) in all fragments. After removing the pericardium, the heart was quickly washed in ice-cold PBS and sagittally cut into four fragments. Both gastrocnemius, from the left and right limbs, were excised, quickly washed in ice-cold PBS, and each one was cut into two fragments, both covering the medial and the lateral portion of the muscle.
All tissue fragments were kept on an ice bath for 1 hour before being freshly processed or frozen. After that, one fragment of each tissue was immediately processed for the oxidative stress marker analysis (fresh tissue [FT]), as described below. The other fragments were immediately transferred from the ice bath to storage in an ultrafreezer (−80°C), without any cryoprotectant solution. Tissue fragments were kept frozen for 1 (TM1), 3 (TM3), or 6 months (TM6) before being processed. Immediately before processing, frozen tissues were removed from the ultrafreezer and thawed on ice (for ∼15 minutes).
Tissue fragments were homogenized in a Potter-Elvehjem tissue grinder, with ice-cold PBS, on an ice bath. After homogenization, samples of fresh and frozen/thawed tissue fragments were divided into two aliquots. One of them was centrifuged at 5000 g for 5 minutes at 4°C, and the supernatant was used for the quantification of the TBARS and the nonenzymatic total antioxidant capacity (TAC). The pellet was used to measure the content of protein carbonyl derivatives. The second aliquot was centrifuged at 10,000 g for 10 minutes at 4°C, and the supernatant was used for the quantification of the SOD and CAT activity. The Bradford method 22 was used for tissue protein quantification.
Analyses over time, for all groups, were conducted by the same evaluator, to avoid inter-rater variability. In addition, the same materials, reagents, and equipment were used throughout the analysis.
Markers of oxidative damage to lipids and proteins
The reaction between thiobarbituric acid and malondialdehyde (MDA), the main product of lipid peroxidation, was used to estimate the oxidative damage to lipids. 23 The procedure was conducted as described elsewhere. 24 Briefly, tissue homogenates were incubated with 0.8% thiobarbituric acid, for 90 minutes at 90°C. Absorbance at 532 nm was used for TBARS quantification, through the MDA (1,1,3,3-tetramethoxypropane; Sigma) standard curve. The results are expressed as MDA equivalents per milligram of protein. Measurements were performed in duplicates.
The quantification of protein carbonyl derivatives in tissues, as a marker of protein oxidative damage, was performed as previously described, 25 based on the reaction with 2,4-Dinitrophenylhydrazine (DNPH). Briefly, proteins in samples were precipitated with 10% trichloroacetic acid and then incubated with DNPH (10 mM) in 2 mM HCl, at room temperature, for 30 minutes, protected from light. The protein precipitate was washed twice with ethanol/ethyl acetate (1:1) and then dissolved in 6% sodium dodecyl sulfate. Samples were centrifuged (10,000 g, 10 minutes, 4°C), and the supernatant was evaluated at 370 nm, considering the DNPH molar extinction coefficient of 22,000 M−1 cm−1. The results are expressed as nmol of carbonyl derivatives per milligram of protein. Measurements were performed in triplicates.
Antioxidant capacity
The assay to determine SOD (EC 1.15.1.1) activity was performed according to the Marklund and Marklund's method, 26 as described elsewhere. 27 The inhibition of pyrogallol autoxidation was monitored at 420 nm, during 4 minutes, at 37°C. SOD activity was expressed as units per milligram of protein. One unit of enzyme was considered as being the amount that caused pyrogallol autoxidation inhibition by 50%.
To determine CAT (EC 1.11.1.6) activity, the decay of hydrogen peroxide absorbance at 240 nm was monitored for 1 minute, at 25°C. 28 CAT activity was expressed as millimoles of hydrogen peroxide decomposed per minute, per milligram of protein (ΔE/min/protein mg).
The homogenate's TAC was estimated by the ferric-reducing antioxidant power (FRAP). 29 The reduction of the complex Fe3+-ferric-tripyridyltriazine (TPTZ) to Fe2+-TPTZ, at acid pH, by nonenzymatic antioxidants of the tissue homogenate determines the antioxidant capacity of the sample. The assay was conducted as previously described. 27 Tissue nonenzymatic TAC was expressed as FeII equivalents per protein milligram, determined from the standard curve of known concentrations of FeSO4.
SOD and TAC measurements were performed in duplicates and CAT in triplicate.
Intra-assay and interassay variation coefficients
Intra-assay coefficients of variation (CVs) were calculated from each duplicate for TBARS, TAC, and SOD and for each triplicate for protein carbonyl derivatives and CAT for all assays. The following values were obtained: 2.6%, 5.4%, 5.0%, 5.6%, and 7.0% for TBARS, TAC, SOD, protein carbonyl derivatives, and CAT, respectively. The interassay CVs were calculated from the standards of each trial, and the values were 4.8%, 3.4%, 1.6%, 2.5%, and 1.1% for TBARS, TAC, SOD, protein carbonyl derivatives, and CAT, respectively.
Statistical analyses
Statistical analyses were conducted using GraphPad Prism (v.6.00; GraphPad Software, San Diego, CA). The Shapiro–Wilk test was used to test data normality. Two-way analysis of variance was used for groups and storage time comparisons. Significant differences between means were determined by Tukey (main storage duration effect and intragroup comparisons—CON versus EX, in each sample condition) or Sidak (main exercise effect and intergroup comparisons—CON and EX in frozen versus fresh samples) post hoc tests. Alpha value ≤0.05 was adopted.
Results
Oxidative stress markers were quantified in fresh (FT) and frozen (TM1, TM3, and TM6—1, 3, and 6 months frozen, respectively) tissue fragments (gastrocnemius, heart and brain) from Swiss mice submitted to a maximum swimming exercised (EX), as well as control nonexercised, mice (CON). The exercised animals reached a mean maximum workload of 3.2 ± 0.7 g (8.3% ± 2.1% of body mass), and the mean time to exhaustion was 2078.0 ± 600.0 seconds. The animals of the control group were maintained for 30 minutes (1800.0 seconds) in the water, under the same temperature conditions of the exercise group (32.0°C).
Markers of oxidative damage to lipids and proteins
TBARS concentration was reduced by storage duration in all tissues examined (main time effect, p < 0.0001), and a significant interaction between the storage duration and exercise on TBARS concentration was also observed (p = 0.005, 0.018, and 0.029 for gastrocnemius, heart, and brain, respectively) (Fig. 1). TBARS concentration was higher in the fresh gastrocnemius of exercised mice (EX FT) compared to the control nonexercised animals (CON FT) (Fig. 1A). This exercise effect on TBARS concentration was also observed in gastrocnemius frozen for 1 (TM1) and 3 months (TM3), comparing EX and CON groups, in each storage condition. However, in 6-month frozen samples (TM6), no difference was observed for TBARS concentration between the CON and EX groups. This indicates that 6 months of storage jeopardize the integrity of the gastrocnemius for TBARS quantification and, therefore, the evaluation of the maximal exercise effect on TBARS in this tissue.
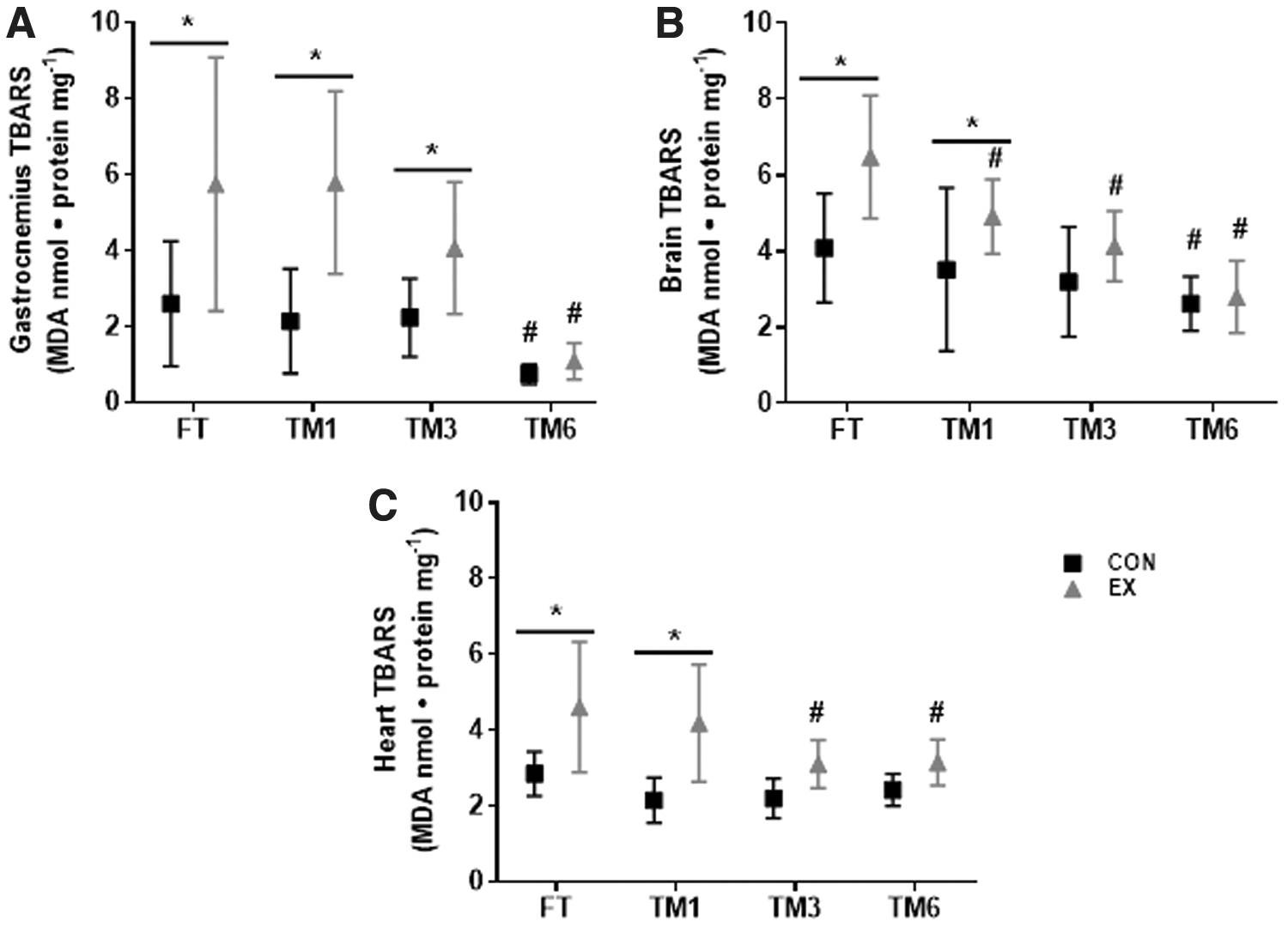
Effect of exercise and storage duration on TBARS concentration in the gastrocnemius
Similar results were observed for brain and heart TBARS quantification (Fig. 1B, C). Although TBARS levels were greater in EX versus CON, in fresh analyzed tissues, and also in tissues frozen for 1 month, no difference was observed between EX and CON in TM3 and TM6. This indicates that the exercise effect on TBARS levels, observed in fresh samples, was lost after sample storage for 3 and 6 months, in both brain and heart. In brain specifically, TBARS levels in samples frozen for 1 month (TM1) were higher in EX versus CON, despite reduced TBARS concentration in EX TM1 compared to EX FT (Fig. 1B). Therefore, the storage of brain and heart samples for 3 months or more interferes with the assessment of the maximal exercise-induced lipid peroxidation. Of note, only in the heart of nonexercised CON mice, TBARS concentration after 6 months of storage (TM6) was not different, compared to FT (TM6 CON versus FT CON) (Fig. 1C).
The storage duration effect on protein carbonyl content quantification was dependent on the tissue analyzed (Fig. 2). Protein carbonyl content was higher in the gastrocnemius of exercised animals (p < 0.0001, EX FT versus CON FT), and storage duration had no effect on the concentration of this marker in either CON or EX groups (Fig. 2A). In the brain, protein carbonyl content was also greater in the EX versus CON group, in freshly analyzed tissue (FT), as well as in tissues frozen for 1, 3, and 6 months, despite the reduction of this marker in brain samples of exercised animals frozen for 3 and 6 months (EX TM3 and EX TM6), compared to EX FT (Fig. 2B). Thus, freezing for up to 6 months did not jeopardize the evaluation of the exercise effect in the gastrocnemius and brain protein carbonylation, despite the storage duration effect. In the heart, different from gastrocnemius and brain, exercise had no effect on protein carbonyl content, either in fresh or frozen tissue, independent of the storage duration (Fig. 2C, EX versus CON, in each tissue condition). However, storage duration affected heart protein carbonyl content stability, as this marker was greater in EX TM3 and EX TM6 heart samples, compared to EX FT (p < 0.5 and p < 0.01, respectively) and also in CON TM6 compared to CON FT (p < 0.05).
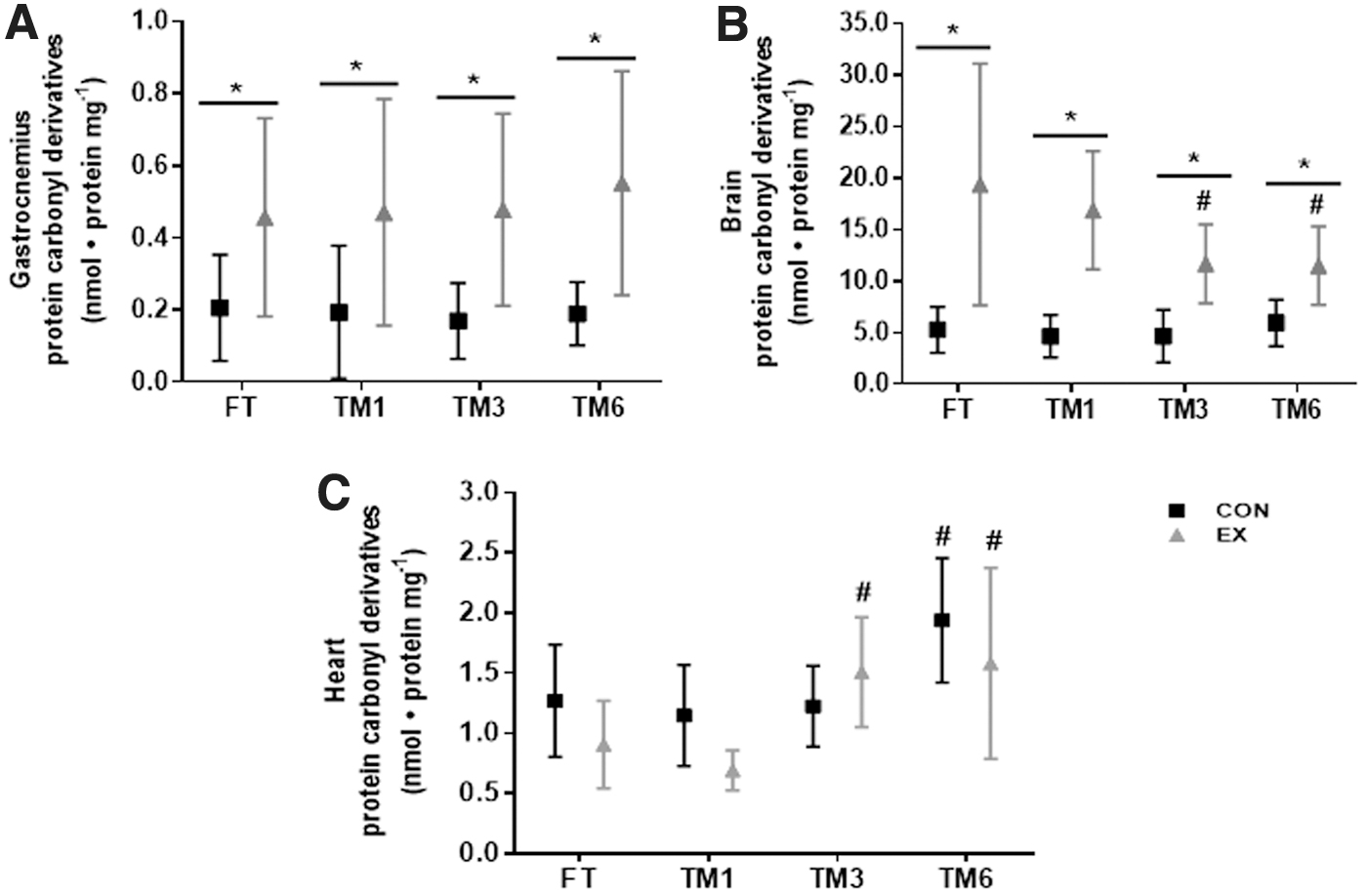
Effect of exercise and storage duration on protein carbonyl content at the gastrocnemius
Antioxidant capacity
Sample storage reduced CAT activity in the gastrocnemius and heart (p < 0.0001, main time effect), and interaction between the storage duration and the exercise effect was observed (p < 0.0001) (Fig. 3). CAT activity was higher in the gastrocnemius (Fig. 3A) and heart (Fig. 3B) in fresh samples of exercised animals (EX FT) compared to nonexercised CON group (CON FT). The enzyme activity was reduced in samples frozen for 1 month or more in both CON and EX groups, and thus, no differences were observed in CAT activity between EX and CON groups in samples frozen for 1 month or more. These data indicate that the storage of the gastrocnemius and heart for 1 month interferes with the assessment of the CAT activity response to the maximal exercise. We were unable to quantify CAT activity in the brain, probably because the amount of sample used was insufficient for the assay.

Effect of exercise and storage duration on CAT activity in gastrocnemius
SOD activity was also reduced by storage duration in the gastrocnemius, heart, and brain (main time effect, p < 0.0001), and a significant interaction between the storage duration and exercise on SOD activity was observed (p < 0.0001, 0.0032, and 0.0001, for the gastrocnemius, heart, and brain, respectively) (Fig. 4). In all the three tissues, SOD activity was greater in EX versus CON, in freshly evaluated samples (EX FT versus CON FT). This exercise effect was still observed in brain samples frozen for 1 month (EX TM1 versus CON TM1, p < 0.0001) (Fig. 4B) and also in 1-month frozen heart samples (p < 0.01), despite lower SOD activity in both CON TM1 and EX TM1 compared to FT (Fig. 4C). However, in the gastrocnemius, SOD activity in EX group was lower in samples frozen for 1 month or more, compared to EX FT, and thus, the exercise effect on gastrocnemius SOD activity was abolished by sample storage (Fig. 4A). No difference was observed in SOD activity for CON in the gastrocnemius, throughout the storage period tested (Fig. 4A). Storage duration equal or superior to 3 months also abolished the exercise effect on heart SOD activity observed in FT, as no difference was observed between CON and EX groups in both TM3 and TM6 (Fig. 4C). Heart SOD activity in TM1, TM3, and TM6, in both CON and EX groups, was lower compared to their respective groups in FT. Similar results were observed in the brain: lower SOD activity in samples frozen for 3 months or longer, in both exercised and control mice, compared to the data obtained in freshly evaluated tissue (Fig. 4B). Therefore, in the brain sample storage for 3 months or longer also abolished the exercise effect on SOD. These findings show that sample storage affects the quantification of SOD activity in response to maximal exercise in the gastrocnemius, brain, and heart in a tissue specific manner.
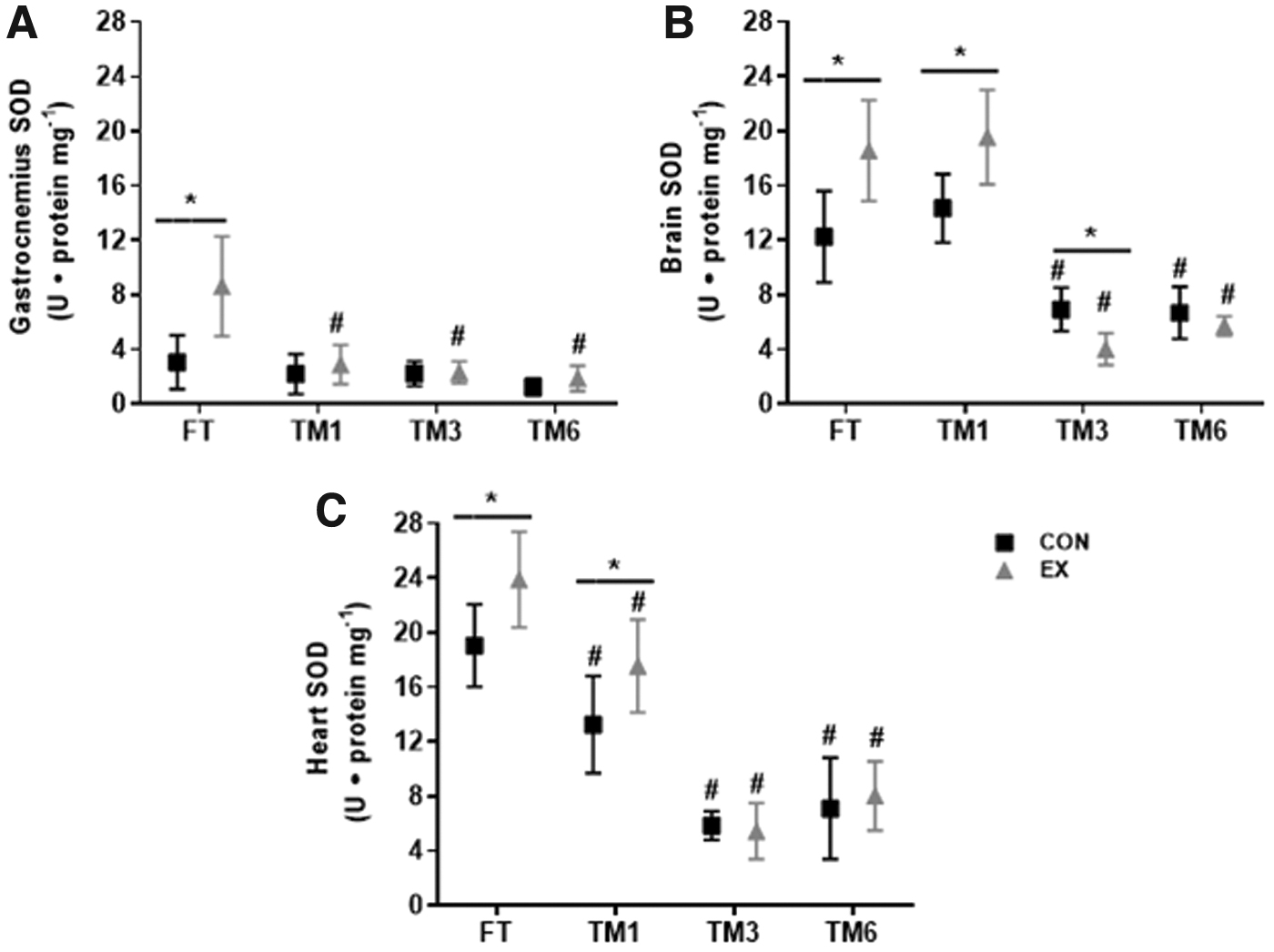
Effect of exercise and storage duration on SOD activity in gastrocnemius
The TAC of gastrocnemius, heart, and brain was lower in the exercised animals (p < 0.0001, main exercise effect) compared to the nonexercised CON group, regardless of the sample storage duration (Fig. 5). In the gastrocnemius, TAC was greater in EX versus CON, in all situations (FT and frozen samples), and thus, sample storage duration had no effect on TAC (main time effect, p = 0.65) (Fig. 5A). The same was observed in the brain and heart. TAC was lower in EX compared to CON in all conditions (fresh and frozen tissue samples), despite lower TAC in TM6 in EX and CON, compared to EX FT and CON FT in both tissues (Fig. 5B, C). Thus, sample storage duration did not affect the evaluation of the exercise on gastrocnemius, heart, and brain TAC.
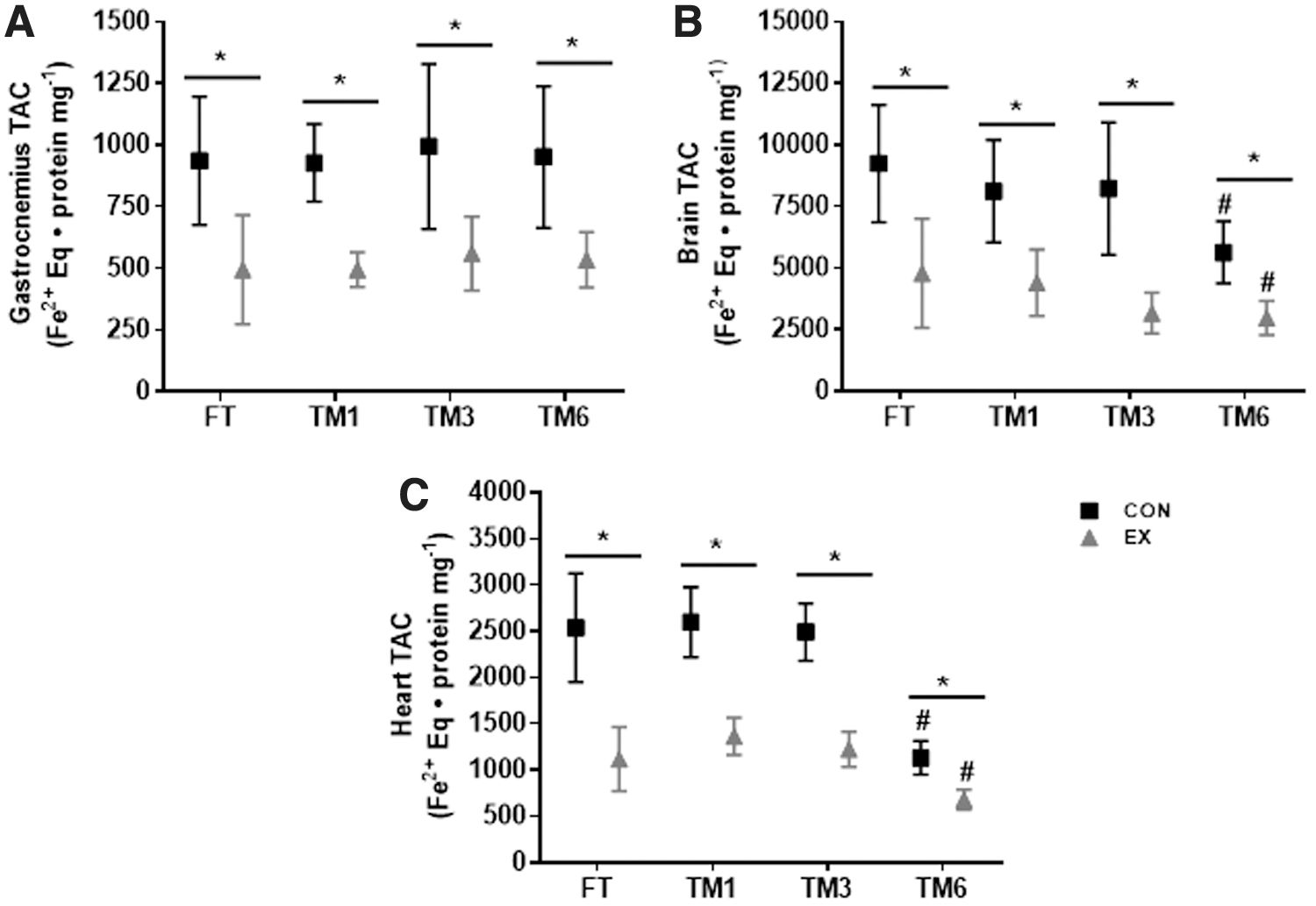
Effect of exercise and freezing on total nonenzymatic antioxidant capacity of gastrocnemius
Discussion
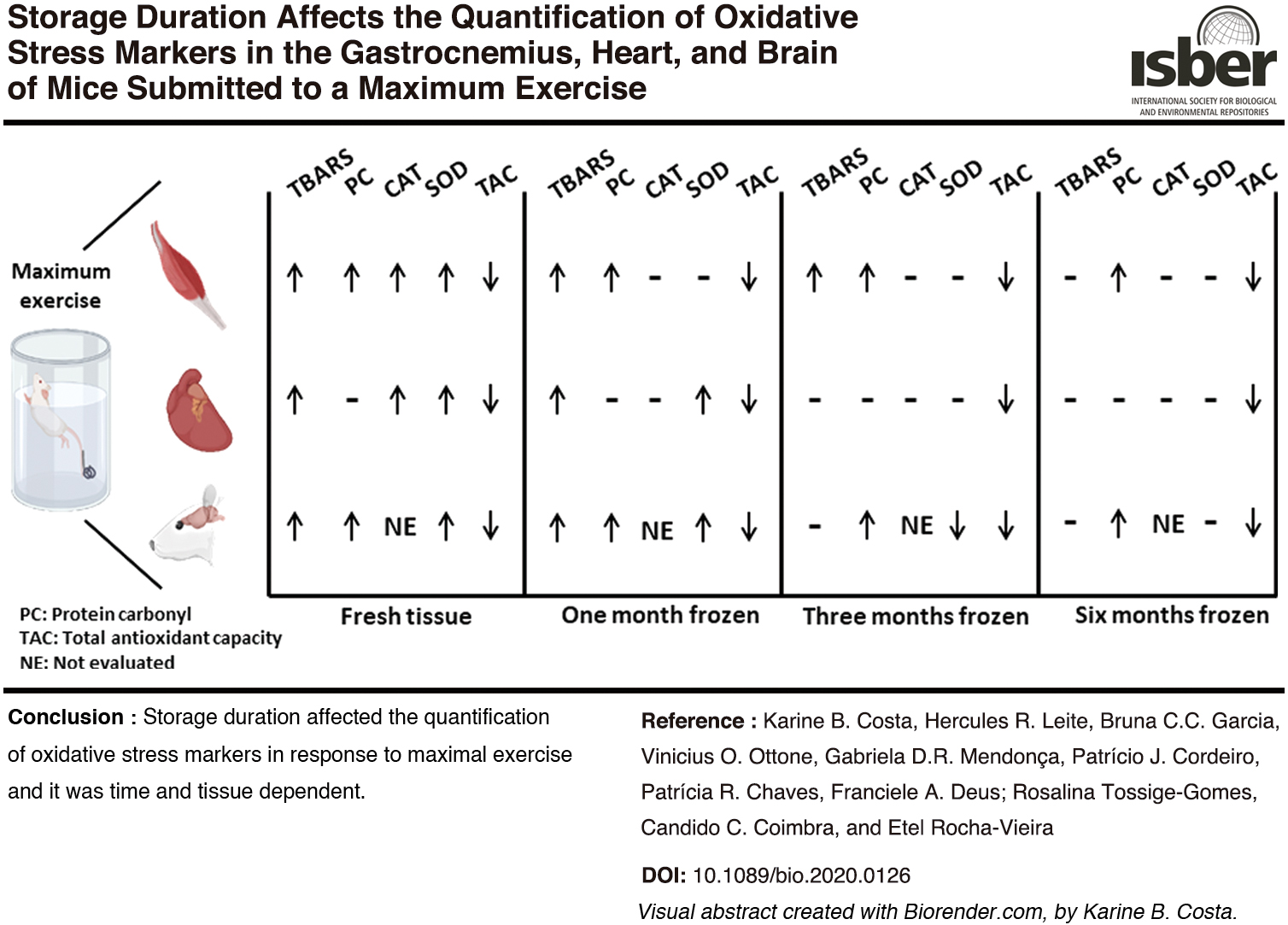
In this study we investigated whether sample storage duration may interfere with the quantification of oxidative stress markers in the gastrocnemius, heart, and brain of mice submitted to a maximum swimming protocol. Maximum exercise led to alterations in the redox status of all tissues, and the stability of all markers evaluated was affected by storage duration. However, storage duration interfered with the evaluation of the exercise effect on TBARS concentration and the SOD and CAT activity, and the effect was tissue specific (Table 1).
Effect of Storage Duration on the Quantification of Redox Biomarkers in the Gastrocnemius, Heart, and Brain in Response to Maximal Exercise
† = freezing interference on maximal exercise response; — = no freezing interference on maximal exercise response.
CAT, catalase; FT, fresh tissue; Gastroc., gastrocnemius; SOD, superoxide dismutase; TAC, total antioxidant capacity; TBARS, thiobarbituric acid reactive substance; TM1, treatment 1 month of frozen; TM3, treatment 3 months of frozen; TM6, treatment 6 months of frozen.
Some studies have previously shown that the SOD and CAT activity in the liver and kidneys of rats are reduced in samples frozen for 1 to 2 months,18,20 but these studies are limited by the tissues examined, the 2-month maximal storage duration, and none investigated whether the response of these markers to physiological and nonphysiological stimuli could be affected by sample storage duration.
As previously reported, maximum exercise resulted in increased TBARS concentration in fresh-analyzed gastrocnemius, heart, and brain.21,30,31 We observed that TBARS concentrations were not stable in frozen tissues, and therefore, the quantification of the maximum swimming effect on lipid peroxidation was affected. This, however, was time and tissue specific. Frozen gastrocnemius samples are stable to the quantification of the exercise-induced TBARS response for up to 3 months, while heart and brain samples should be analyzed up to 1 month after freezing at −80°C. These tissue-specific differences can be related to the magnitude of the exercise response: in the gastrocnemius the exercise promoted a twofold increase in the TBARS concentration, while in the brain and heart this response was 40% lower. In addition, intrinsic differences in tissue composition may account for the stability of its analytes. 32
Maximum exercise also resulted in increased protein carbonyl derivative levels in the gastrocnemius and brain, but not in the heart,8,30,33 and the exercise effect was not affected by sample storage duration. Some may consider some of our observations, together with the TBARS data, contradictory to the idea that exercise is beneficial for health, including brain health.34,35 However, some aspects of the exercise physiology are worth mentioning. First, a single bout of exercise is generally related to some degree of tissue oxidative damage, an effect that is necessary to trigger many physiological adaptations to exercise that are related to improved health, including adaptations of the tissue antioxidant capacity.1,36,37 Second, we intentionally used in this study a model of maximum exhaustive exercise, in which mice swam until they were not able to maintain muscular activity anymore and sank. Maximum, unaccustomed exhaustive exercise is known to induce extensive oxidative damage to tissues, as seen here, that can be detrimental to tissue function, despite the induction of compensatory antioxidant mechanisms.1,21 Finally, the brain is very sensitive to oxidative stress because of its high metabolic activity, high density of oxidizable substrates, and relatively low antioxidant defense. 38 Together, all these aspects can explain the high degree of oxidative damage to this tissue reported here.
The stability of protein carbonyl derivatives, under basal conditions, in rat brain and liver frozen at −80°C for 2 months was previously reported. 18 Carbonylation is an irreversible form of protein modification, and this biomarker is very stable, compared to lipid peroxidation products.39,40 Now we show the stability of this marker to freezing also in response to the exercise stimulus. We suggest that protein carbonyl derivatives should be the marker chosen to investigate the exercise effect on oxidative stress when samples cannot be freshly analyzed, or they need to be frozen before the analysis.
Freezing can lead to protein aggregation, denaturation, and loss of the tertiary structure,41,42 and studies have shown reduced CAT and SOD activity in frozen tissues.18,20 We observed a drastic reduction in the activity of both enzymes in frozen tissues, and therefore, the quantification of the exercise effect on the activity of SOD and CAT was affected. SOD stability to storage duration is tissue specific. In the gastrocnemius, the exercise effect in SOD activity was abolished in 1 month, while in the heart and brain this effect was abolished in samples frozen for 3 months or more. The early deterioration of SOD activity in the gastrocnemius can be due to a lower concentration of SOD in this tissue, as it is suggested that the freezing damage is inverse to the enzyme concentration.43,44 Thus, our findings indicate that fresh samples are the best option for the quantification of SOD and CAT activity in response to the maximum swimming exercise. It will be important to show whether the exercise effect on SOD and CAT activity can be preserved in samples frozen for less than 1 month.
Different from the antioxidant enzymes, the quantification of the TAC response to maximal exercise was not affected by sample storage duration. As observed for protein carbonyl derivatives, this may be a consequence of the stability and/or freezing resistance of the compounds evaluated in the assay, which include uric and ascorbic acids, among others. 29 Our findings show that the analysis of TAC by the FRAP assay, as well as protein carbonylation quantification, can be a choice when the experimental design does not allow the quantification of oxidative stress markers in fresh samples.
For some of the biomarkers evaluated, the storage duration effects were manifested earlier in the samples of the exercised animals (TBARS levels in the heart and brain) or it was observed only in the exercise condition (protein carbonyl derivatives in brain and SOD activity in the gastrocnemius). The interaction between storage duration and the exercise may be a consequence of the physiological alterations imposed on tissues by the maximal exercise. Exercise alters not only tissue redox homeostasis but also several other processes, including metabolic and ionic homeostasis. 45 Maximum exercise also leads to dehydration and increased internal temperature and substrates depletion, 46 which may affect the stability of some biomarkers during the storage period. This can explain why in some situations the storage duration effect was dependent on the exercise condition. In addition, our findings show that the experimental situation to which the tissues are exposed can modify the biomarker stability, and therefore, this must be considered during the planning of an investigation, even when the stability of the biomarker under basal conditions is known.
Some procedures were followed in this study to assure that our findings are mainly due to the storage duration and not to other preanalytical conditions. The samples were carefully sectioned into four equal parts and frozen and thawed only once at the moment of the assays. We chose to conduct the analyses in frozen tissue fragments instead of homogenized samples, as it has been suggested that samples frozen in aqueous solution are more vulnerable to degradation and oxidative modifications. 18 This procedure was also adopted to minimize pH alteration and consequent selective crystallization, ice-water interface formation, and eutectic crystallization, factors that may affect the stability of the biomarkers evaluated.47,48 Moreover, proteins in solution have a short-term storage stability.44,47,49 In addition, the tissues were fractioned into four segments, under low temperature conditions (ice bath), as the probability of artifactual proteolysis occurrence, sample degradation, and loss of its analytical utility is greater when the sample is fractionated in a large number of fragments or it is submitted to many freeze–thaw cycles. 50
Conclusion
In conclusion this study shows that the quantification of TBARS, SOD, and CAT activity in response to the maximum swimming exercise is affected by sample storage duration. As a result of our study the following recommendations are proposed: nonfrozen, freshly excised samples should be prioritized for the quantification of CAT and SOD activity; TBARS should be quantified in the gastrocnemius frozen for up to 3 months and in the heart and brain frozen for up to 1 month; TAC and protein carbonyl derivatives should be the markers of choice for long-term frozen samples. These recommendations can contribute to more accurate and reproducible studies in the field of exercise physiology and oxidative stress markers.
Footnotes
Acknowledgments
The authors acknowledge the Centro Integrado de Pós-graduação e Pesquisa em Saúde at the Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, Minas Gerais, Brazil, for providing equipment and technical support for experiments.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The work was supported by the Brazilian Funding agencies CNPq, Fapemig, and Capes (grant number 2004/2015).