
Abstract
Cell-based therapeutics promise to transform the treatment of a wide range of diseases, many of which, up to this point, are incurable. During the past decade, an increasing number of cell therapies have been approved by government regulatory agencies in the United States, Europe, and Japan. Thousands of clinical trials based on live cell therapies are now taking place around the world. But most of these live cell therapies face temporal and/or spatial distances between manufacture and administration, posing a risk of degradation in potency. Cryopreservation has become the predominant biobanking approach to maintain the product's safety and efficacy during transportation and storage. However, the necessity of cryogenic shipment and storage could limit patient access to these emerging therapies and increase the costs of logistics. In the (bio)pharmaceutical industries, freeze-drying and desiccation are established preservation procedures for manufacturing small molecule drugs, liposomes, and monoclonal antibodies. Over the past two decades, there has been a growing body of research exploring the freeze-drying or drying of mammalian cells, with varying degrees of success. This article provides an overview of the technologies that were adopted or developed in these pioneering studies, paving the road toward the preservation of cell-based therapeutics in a dry state for biomanufacturing.
Introduction
Cell therapy is considered to be one of the most promising new drug therapeutics, which is represented by a growing list of products approved by government regulatory agencies in the United States, Europe, and Japan over the past decade (e.g., KYMRIAH from Novartis, YESCARTA and TECARTUS from Kite Pharma, and GINTUIT from Organogenesis, etc.). By now, thousands of clinical trials based on live cell therapies are already taking place around the world. Many of the conditions that are being investigated by these trials are incurable up to this point.
For cell therapy to transition from promises to products, we not only need to identify the biology that impacts the mechanism of action but to develop large-scale, cost-effective, and reproducible biomanufacturing processes, 1 thereby reducing the cost and improving patient access to these life-saving therapies. 2 Although shelf life in many early-phase trials conducted in hospital or laboratory settings is of limited concern, long-term preservation of cell therapies, especially for “off-the-shelf” cell products, may play a critical role in late-phase clinical trials and commercialization. 3
So far, cryopreservation has been the predominant biobanking approach for extending the shelf life of cells and tissues. However, cryopreservation requires a cryogenic-temperature cold chain, which is usually costly and cumbersome. The failure of such a cold chain may result in the complete loss of valuable products.
Alternatively, preserving biological samples in a dry state can provide both economic and practical advantages, such as transportation flexibility, refrigerated or ambient storage, and reduced product weight. 4 In the pharmaceutical and biopharmaceutical industries, freeze-drying and drying have been used increasingly for manufacturing small molecule drugs and biologically active molecules such as enzymes, hormones, antibiotics, and monoclonal antibodies (mAb), owing to their robust and reproducible procedures.5,6 To avoid confusion, the term “drying” or “desiccation” used in this article specifically refers to the methods of removing water from the sample through evaporation without freezing.
Freeze-drying is a low-temperature, vacuum dehydration process. As illustrated in Figure 1A, liquid water is first converted to ice by lowering the temperature of the sample at ambient pressure (path 1–2). During the primary drying (path 2–3), water is removed through sublimation as the pressure is lowered below the triple point of the system (e.g., 0.01°C and 0.6 kPa for pure water) and a certain amount of heat is also added to the system. At the end of the primary drying, there is still ∼8%–10% water remaining. The remaining unfrozen water, also known as the bound water, can be removed by desorption during the secondary drying (path 3–4) by slowly raising the temperature under vacuum, yielding a lyophilized product of long-term stability at the refrigerated or ambient temperature.
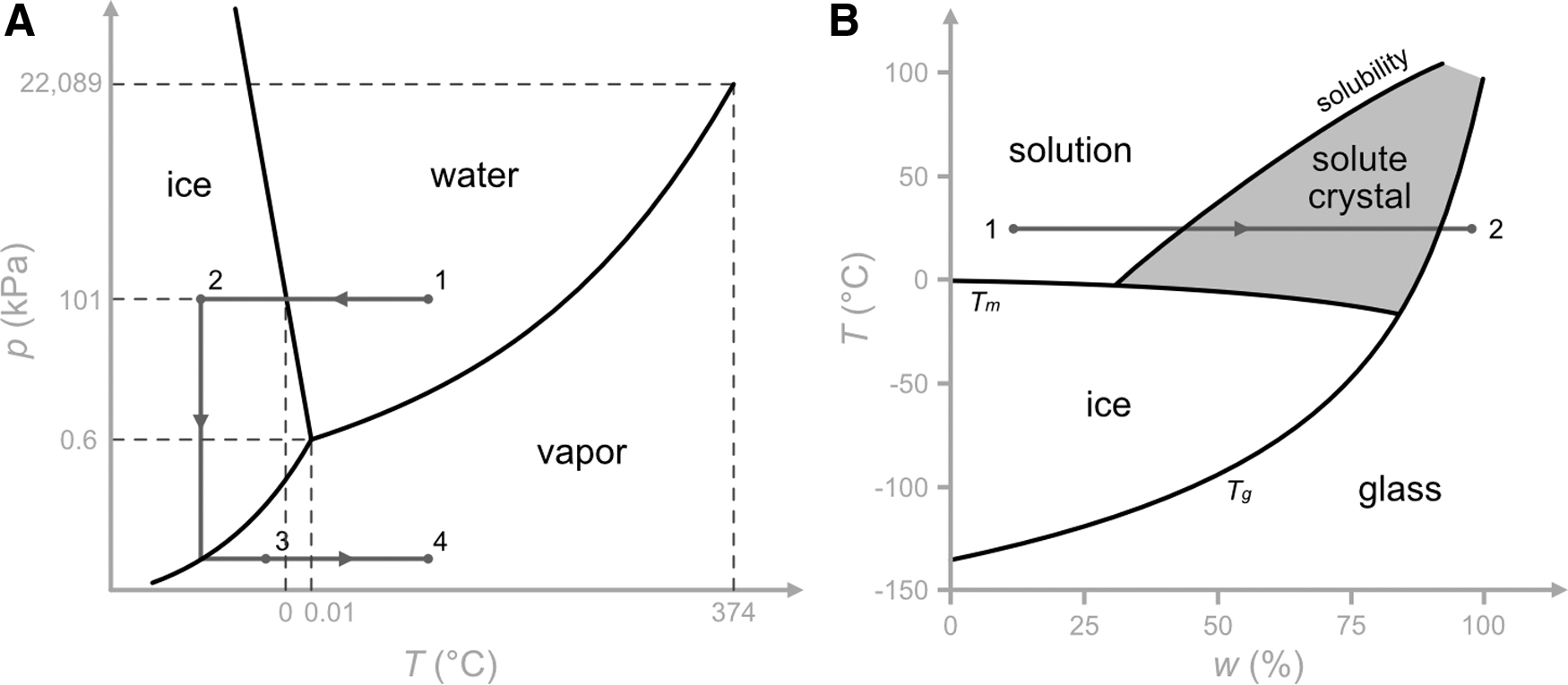
Freeze-drying and desiccation processes.
During desiccation, as illustrated in Figure 1B, water is removed from the aqueous material by evaporation at the ambient or an elevated temperature until the glass transition temperature (Tg) of the residual concentrated solution becomes higher than the processing temperature to trigger the glass transition.
To date, the majority of the applications of freeze-drying or drying of mammalian cells could only preserve the integrity and/or functionality of certain subcellular components.7–14 For example, freeze-drying of EBV-transformed B-lymphoblastoid cells could stabilize the total RNA in the cells for routine diagnostics. 15 Surface-labeled, freeze-dried lymphocytes can be used as the reference material for counting CD4+ T cells.16,17 Although it is unlikely that cells in the tissues can survive freeze-drying, freeze-dried acellular matrices can be used as biological scaffolds in regenerative medicine.18,19 Besides, spray drying was used to preserve Bacillus Calmette–Guérin (BCG) vaccines, showing no significant loss of activity after storage at the refrigerated or room temperature for up to 4 months.
To prime dry preservation and noncryogenic storage of therapeutic cells for biomanufacturing, we need comprehensive understanding of the cellular and molecular injuries associated with freezing and/or drying and full optimization and control of formulation and processing. 20 This article reviews the existing strategies that have been explored to improve the outcomes of freeze-drying or drying of mammalian cells over the past two decades. The motivation of this review was to promote continued innovation in the field that would ultimately enable the preservation of live cell therapies in a dry state.
Intracellular Trehalose Delivery
In nature, seeds of many plants, yeasts, certain insects, nematodes, and tardigrades can survive extreme desiccation (i.e., the loss of essentially all the body water) by entering a state of anhydrobiosis.21,22 The survival of many organisms in anhydrobiosis was found to be correlated with the accumulation of nonreducing disaccharides, usually trehalose or, in the case of higher plants, sucrose, in the body.23–25
The “water replacement” hypothesis proposes that trehalose can substitute for water at a low moisture content through hydrogen bonding with lipid head groups and proteins to keep their native structures in the absence of hydration.21,26,27 It was also suggested that trehalose could function as a “chemical chaperone” by working synergistically with heat shock proteins to stabilize native protein conformation during desiccation.23,28 Moreover, trehalose also has a higher glass transition temperature than other comparable sugars.
Although trehalose cannot typically permeate cell membranes, researchers have discovered the unique benefits of intracellular trehalose in helping mammalian cells, which cannot synthesize trehalose naturally, survive freezing or drying in the absence of other cryo- or lyoprotectants.29,30
Eroglu et al. 29 delivered trehalose to 3T3 murine fibroblasts and human primary foreskin keratinocytes using a recombinant pore-forming hemolytic protein, α-hemolysin. They found that a low concentration (0.2 M) of trehalose permitted post-thaw survival of >80% of murine fibroblasts and 70% of human keratinocytes after cryopreservation. In another study, Guo et al. 30 demonstrated that human primary fibroblasts expressing trehalose could be preserved in a dry state for up to 5 days. In their study, using a recombinant adenovirus vector, human primary fibroblasts were transfected with otsA and otsB genes of Escherichia coli to encode trehalose biosynthetic enzymes (i.e., trehalose-6-phosphate synthase).
Given the exceptional benefits of intracellular trehalose, a variety of approaches have been used or developed to deliver trehalose into mammalian cells, as illustrated in Figure 2. They include fluid-phase endocytosis, 31 pore-forming cell surface receptor, 4 transmembrane trehalose transporter, 32 nanoparticle mediation,33,34 ultrasonic radiation,35,36 and electroporation,37,38 which have been reviewed in the literature. 39
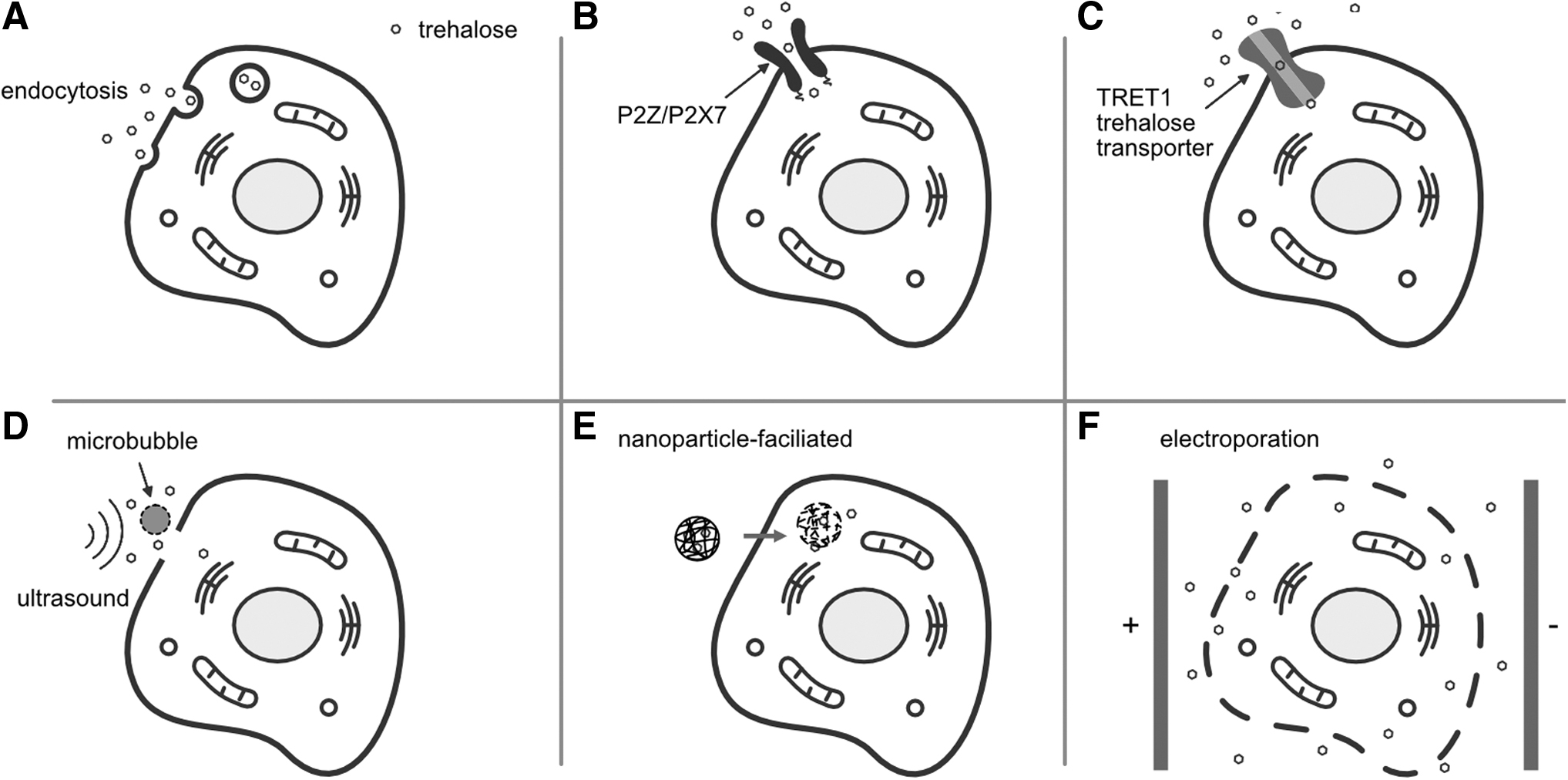
Representative techniques for intracellular trehalose delivery.
Wolkers et al. 31 freeze-dried human platelets, resulting in a postrehydration survival rate of 85%. Before freeze-drying, these platelets were loaded with trehalose through fluid-phase endocytosis at 37°C (Fig. 2A). The rehydrated platelets retained clotting properties, being able to respond to agonists thrombin, collagen, adenosine diphosphate, and ristocetin comparable with fresh platelets, indicating their potential usage for topical wound healing. 31
Buchanan et al. 4 demonstrated that hematopoietic stem and progenitor cells (HSPCs) isolated from human umbilical cord blood could be freeze-dried and stored at ambient temperature for 4 weeks when the cells were loaded with 0.2 M trehalose through membrane pore formation by the endogenous cell surface receptor (P2Z) (Fig. 2B). Their results suggested that there was no statistical difference observed for HSPCs removed at the end of the freezing step or at the end of the primary drying, compared with the fresh control. 4 When the cells were removed during secondary drying, however, there was a gradual decrease in the number of colonies formed postrehydration as the secondary drying temperature increased from 5°C to 20°C, 4 implying the importance of optimizing the secondary drying even in the presence of intracellular trehalose.
Trehalose was also introduced into Chinese hamster ovary (CHO) cells using the high-capacity trehalose transporter 1 (TRET1) (Fig. 2C) before the cells were rapidly and uniformly desiccated to a low moisture content using a spin drying technique. 32 TRET1 is a transmembrane protein originally expressed in anhydrobiotic insects induced by either desiccation or salinity stress. 40 The authors found that 62% of the cells survived spin drying in the presence of intracellular trehalose after immediate rehydration. 32 Furthermore, 51% of the spin-dried and vitrified cells survived and demonstrated normal growth characteristics after prolonged storage in liquid nitrogen. 32
Ultrasound is known to be an enhancer for drug delivery. 41 It was also found to facilitate the loading of trehalose into human platelets, with no significant change to the number and morphology of cells after ultrasonic radiation. 35 Local deposition of ultrasound energy can further perturb cell membranes and increase the permeability. Thus, gas-filled microbubbles, a type of widely used contrast agent in medical imaging, were used to serve as the deposition foci and aid in a wide range of ultrasound-mediated delivery of molecular compounds. 42
Centner et al. 36 combined ultrasound and microbubbles to induce transient pore formation in cell membranes and enable the delivery of trehalose to human erythrocytes flowing through a microfluidic device (Fig. 2D). Although this study demonstrated the feasibility of an acoustofluidic approach for intracellular delivery of trehalose, the subsequent freeze-drying of these trehalose-loaded cells only yielded <5% postrehydration recovery.
Zhang et al. 33 used poly (N-isopropylacrylamide-co-butyl acrylate), a cold-responsive polymer, to synthesize nanoparticles for the encapsulation and intracellular delivery of trehalose (Fig. 2E). These trehalose-laden nanoparticles were demonstrated to be efficiently taken up by mammalian cells by preincubation for only 4 hours at 37°C and be able to quickly and irreversibly disassemble upon cold treatment, which provides a potential trehalose delivery approach suitable for freeze-drying applications.
Late Embryogenesis Abundant Protein Expression
Although nonreducing disaccharides such as trehalose are involved in the desiccation tolerance of many anhydrobiotic organisms, trehalose does not appear to be essential to other organisms. Hengherr et al. 23 reported that certain species of tardigrades such as Milnesium tardigradum did not accumulate any trehalose in response to desiccation, whereas Echiniscus granulatus and Echiniscus testudo possessed low trehalose concentrations with no significant change in the trehalose level during the anhydrobiotic states.
Thus, nonsugar-aided strategies must be used by anhydrobiotic organisms like M. tardigradum because other carbohydrate compounds were also not detected during drying. For example, the induction of late embryogenesis abundant (LEA) proteins has also been found to correlate with desiccation tolerance.43,44 LEA proteins were originally associated with the acquisition of desiccation tolerance by plant embryos in the later stages of seed maturation. 45 In addition to abiotic-stressed plants, they were also found in bacteria 46 and invertebrates such as nematode Aphelenchus avenae 47 and brine shrimp Artemia franciscana. 48
LEA proteins are rich in polar amino acids, highly hydrophilic, and intrinsically disordered. In the fully hydrated state, LEA proteins are predominately unstructured in the form of a random coil. Upon dehydration, LEA proteins exhibit a remarkable ability to become more ordered and to develop secondary structure, as demonstrated by a previous molecular dynamics (MD) simulation study (Fig. 3).46,49
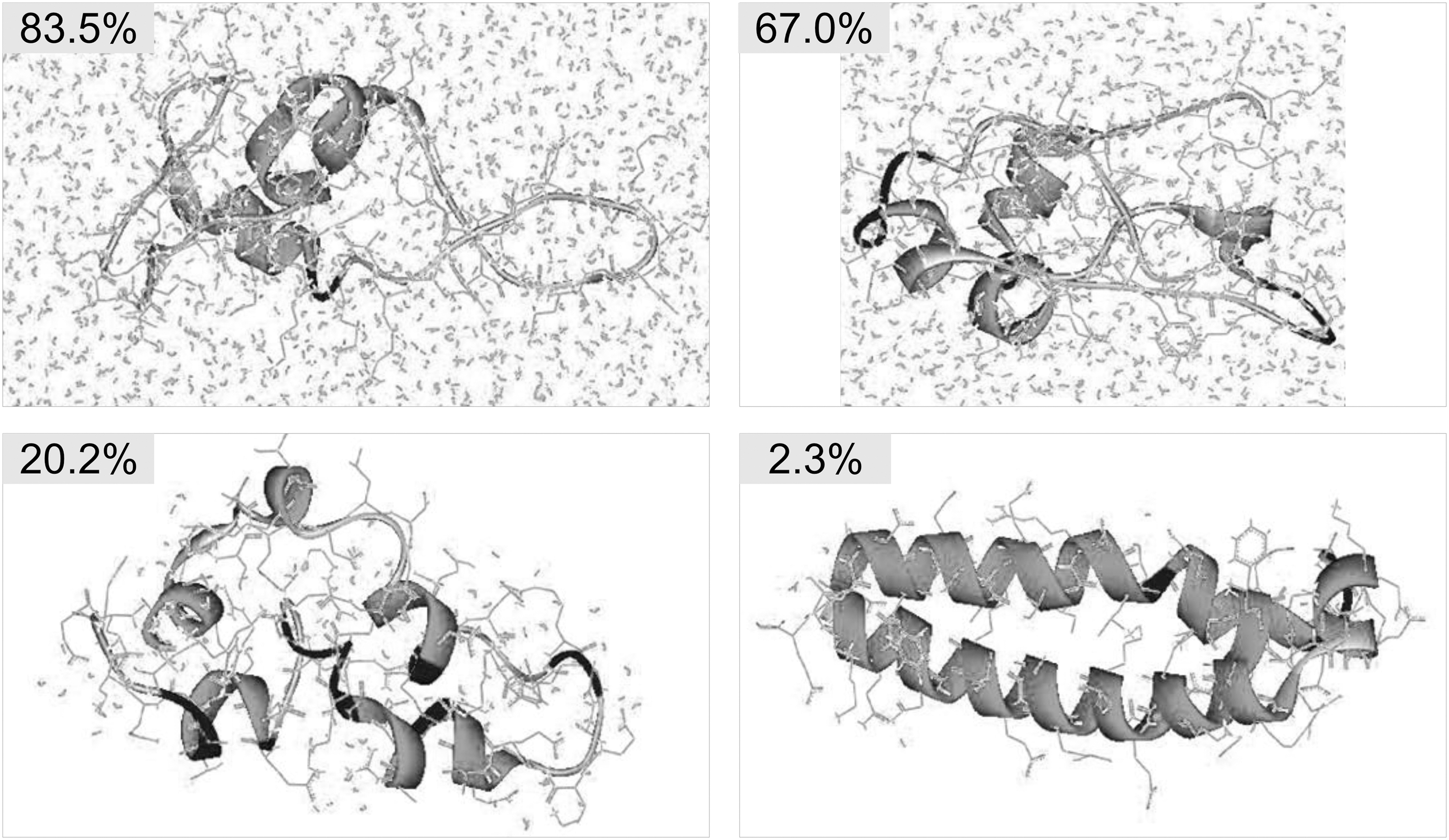
Intrinsically disordered structure of a LEA protein fragment. Molecular dynamics simulations suggested the structure of a 66-amino acid fragment of AavLEA1 in aqueous solutions at different water contents. At a water content of 83.5%, the LEA protein is fully solvated and largely unstructured. But a hairpin-like conformation with two α-helical bundles emerges in response to desiccation. LEA, late embryogenesis abundant. Images were adapted from Li and He 49 with permission from the American Chemical Society.
Wang et al. 50 investigated the effect of AavLEA1 protein, a member of the Group 3 LEA protein family, on the cryopreservation of human umbilical cord matrix mesenchymal stem cells (hUCM-MSCs) by slow freezing. They found that even the extracellular addition of AavLEA1 protein (0.1–2 mg/mL) to 2% dimethyl sulfoxide (DMSO) resulted in higher post-thaw viability (47.1%–53.9%) of cryopreserved hUCM-MSCs than 2% DMSO alone (18.9%).
Afrlea2 and Afrlea3m are another two types of LEA protein, which are naturally expressed in embryos of the brine shrimp A. franciscana. Li et al. 51 engineered a HepG2 cell line by expressing either one of these two LEA proteins. In addition, the TRET1 was also used for intracellular loading of trehalose. They found that 98% of the cells loaded with trehalose and expressing AfrLEA2 or AfrLEA3m and 94% of the cells expressing AfrLEA3m without intracellular trehalose maintained membrane integrity after the cells were desiccated and rehydrated, compared with 0% membrane integrity for the nonengineered cells. 51 The 7-day cell proliferation assay showed an 18-fold increase for cells dried with AfrLEA3m and trehalose, compared with 27-fold for the fresh control. 51
A follow-up study by Moore et al. 52 focused on the stabilizing effect of AfrLEA2 and AfrLEA3m on extrusion-made liposomes simulating the outer mitochondrial membrane, inner mitochondrial membrane, and inner leaflet of the plasma membrane. It was interesting that AfrLEA2 and AfrLEA3m provided essentially equal stabilization of liposomes during drying, regardless of the lipid compositions, although AfrLEA2 and AfrLEA3m are known to be cytoplasmic localized and mitochondrial targeted, respectively, in vivo. 52 Moreover, the enhanced protection offered by trehalose was only demonstrated on the inner mitochondrial membrane and inner leaflet of the plasma membrane but not on the outer mitochondrial membrane, which suggested that the synergetic protection offered by LEA proteins and trehalose may depend on the lipid compositions. 52
By now, the precise mechanisms of LEA proteins to protect cellular or molecular structures from dehydration are still unclear. A number of hypotheses have been proposed, including hydration buffering, 53 ion sequestration, 54 redox balancing, 55 and direct protection of other proteins or membranes. 56 Nuclear magnetic resonance relaxation studies showed that intrinsically disordered LEA proteins were able to bind a significantly larger amount of water than globular proteins. 53 But there is no direct measurement of changes in the kinetics of cellular water loss attributable to the presence of LEA proteins.
Hara et al. 57 demonstrated the binding of a variety of metal ions to dehydrin, a group 2 LEA protein. However, ion sequestration was suspected to be transient and inconsequential given that the number of chelation sites is insignificant relative to the number of total free ions present in the cells. 46 In vitro studies demonstrated that LEA proteins lacked “classical” chaperone activity but were capable of suppressing protein aggregation and inactivation under water deficit conditions. 58 Besides, Pouchkina-Stantcheva et al. 59 reported that the presence of ArLEA1B, a LEA protein from a bdelloid rotifer, could significantly decrease the gel-to-liquid crystalline phase transition temperature of dried liposomes.
Tg Enhancer and Anti-Plasticizer
Both freeze-drying and drying rely on dehydration to concentrate the aqueous material until the Tg of the remaining solution is above the target storage temperature, thus yielding a physically “solid” glassy matrix suitable for long-term storage. Because the viscosity of a glassy matrix usually exceeds 1012 Pa·s, 60 molecular movement is essentially arrested within the matrix and degradation activities can be greatly suppressed. So, formulations that have a higher Tg would provide greater storage flexibility and assure stability against unfavorable events such as temperature fluctuations.
Trehalose is perhaps the most employed excipient in freeze-drying or desiccation of biologics, in part because of its higher Tg (i.e., 100°C–120°C) than other comparable sugars.61–66 Furthermore, a number of additives have been shown to be able to further increase the Tg of trehalose-based compositions. It was shown that the addition of polyvinylpyrrolidone (PVP) or Ficoll to the mixture of trehalose and phosphate-buffered saline (PBS) increased the Tg of the desiccated mixture by 47° or 29°, respectively, compared with trehalose–PBS alone. 67 However, not all polymer molecules are able to raise the Tg of their mixtures with trehalose. Hydroxyethyl starch (HES), for instance, gradually decreased the glass transition temperature of the trehalose–HES mixture, following a Gordon–Taylor curve. 68
Freeze-dried trehalose/sodium tetraborate mixtures (with a boron to trehalose molar ratio of 1:2) were found to have a considerably higher Tg (i.e., 170°C) than that of trehalose alone, owing to the strong interaction between borate ions and trehalose molecules. 69 But it is noteworthy that the buffer concentration and pH may interfere with the borate–trehalose interaction and thus affect the final Tg, suggesting a critical role of buffering and pH in determining the glass transition temperature.
Another study showed that, for a given ratio of trehalose to phosphate, the Tg of the trehalose–phosphate mixture increased as the pH rose from 4 to 9, as phosphate changed from monobasic (
It is generally speculated that solvent viscosity is the predominant contributor to suppressing conformational fluctuations within biomacromolecules like proteins.72,73 In some cases, however, the stability of proteins embedded in sugar glasses has been proven not directly related to the viscosity or Tg, but being a function of the fast, high-frequency dynamics of the solvent matrix, which occurs on the scale of picoseconds.74–79
Cicerone et al. 74 demonstrated that adding a small amount (5%) of glycerol to sugar glasses (i.e., trehalose or lactose) substantially improved the stability of freeze-dried enzymes such as horseradish peroxidase and yeast alcohol dehydrogenase, although the final Tg of the glycerol–sugar mixture was reduced by ∼20° compared with pure trehalose.
It was later found that there was a nonmonotonic relationship between the fragility and the mass fraction of trehalose in the above mixtures, with a local minimum between 87.5% and 95% trehalose (i.e., between 5% and 12.5% glycerol), implying an anti-plasticizing, glass strengthening role of glycerol. 79 The fragility of a liquid is correlated with the fast dynamics of the glass it forms. In other words, the liquid of a smaller fragility often has suppressed fast dynamics. Therefore, the above study demonstrated that the addition of 5%–12.5% glycerol could result in a local minimization of fragility, without compromising the Tg significantly. 79
Rapid and Uniform Drying Techniques
Unlike freeze-drying, drying evaporates water from the aqueous material rapidly at the ambient or elevated temperature to outrun the solute crystallization and thereby ensure vitrification of the whole matrix, as described previously in Figure 1B. To accelerate water evaporation, for example, spray drying atomizes the liquid into droplets of high surface area-to-volume ratios and dries the resulting droplets in a hot gas stream (Fig. 4A). 80 Spray drying has been used in formulating small molecule drugs with low solubility and encapsulating active pharmaceutical ingredients such as peptides and vaccine structure.81,82 In the food industry, spray-drying of probiotic and starter bacteria enables a larger production scale than freeze-drying. 83

Nonfreezing drying techniques that can accelerate water evaporation from biological samples.
Mammalian cells, however, have rarely been the subject of spray drying. On the one hand, the shear stress generated during the atomization process could cause significant mechanical damages. On the other hand, the heat applied during the drying process can potentially result in cellular damages in part associated with changes in the physical state of the membranes, such as the gel-to-liquid crystalline phase transition, modification of the cytoplasm fluidity, and lipid peroxidation. Hence, it is necessary to decouple and mitigate the effects of atomization and drying on cell viability before achieving the successful spray drying of mammalian cells. 84
There is an obvious challenge for traditional convective-only drying approaches, such as desiccation in a low-humidity chamber. In such a case, slow evaporation usually creates a glassy skin at the liquid/vapor interface of the sample, which prevents the desiccation of the sample beyond a certain level of dryness and yields a heterogeneous water distribution across the processed sample. 85 Consequently, the compartment underneath the glassy skin remains to be a viscous liquid and cells trapped in this compartment may still degrade because the molecular mobility is not suppressed sufficiently. 85
Over the past decade, there have been a number of new drying techniques developed to assure a rapid and uniform drying in the absence of freezing or significant heating, such as microwave-assisted drying,86–88 spin drying, 85 light-assisted drying (LAD),89,90 and a surface tension-mediated drying. 91
During the microwave-assisted drying (Fig. 4B), microwaves induce dipolar rotation of polar molecules such as water. The friction among rotating water molecules generates heat. In other words, microwaves can be selectively absorbed by the water in the sample and converted into heat, accelerating the water evaporation.
A series of studies by Elliott and co-workers determined the efficiency and consistency of using microwaves to desiccate sugar-based solutions that were relevant to the drying of biologics.86–88 For example, 40 μL trehalose-based solution (0.5 M trehalose) could be dried to a moisture content as low as 0.1167 gH2O/gdw within 25 minutes, compared with 75 minutes in the absence of microwaves. 87
But when Jurkat E6–1 cells were dried in a 0.2 M trehalose solution using the microwave-assisted drying technique, the postrehydration viability decreased dramatically within the range of 0.1–1 gH2O/gdw. 88 MD simulations proposed that extended, percolating sugar clusters formed when the moisture content approached 0.1 gH2O/gdw. 88 It was hypothesized that when the cells were compressed within the void volumes in a rigidified sugar network, the mechanical shear could induce physical injury to the cells. 88
Given that microwave-assisted drying requires the optimization of microwave pulse and exposure time to prevent overheating, Chakraborty et al. 85 developed a spin drying technique that forced convective evaporation and vitrification of a thin layer of sugar-containing cell suspension by spinning the glass substrate in air (Fig. 4C). It was shown that >95% of CHO cells maintained the membrane integrity after they were spin-dried in a trehalose-based solution to 0.159 gH2O/gdw within 50 seconds. In comparison, convective drying in a controlled humidity environment could take 40–60 minutes to reach a similar moisture content. 92
Recently, Young et al.89,90 developed a LAD method taking advantage of the selective photothermolysis principle of laser treatment (Fig. 4D). The LAD method used illumination by near-infrared laser light to accelerate the water evaporation and dehydration of the sample. The wavelength was selected such that water absorbed most of the energy, whereas there was little absorption by key subcellular components such as nucleic acids and proteins.
The effect of LAD processing on protein functionality was investigated. 90 Using a 1064-nm laser at 5.0 W, the LAD of a 40-μL droplet of lysozyme formulated in a 0.2 M trehalose solution for 60 minutes yielded an end moisture content of 0.17 gH2O/gdw and resulted in no significant loss of the specific activity of lysozyme compared with the refrigerated control, although the maximum processing temperature had reached up to 44°C. It was found that increasing the wavelength could shorten the drying time but at the cost of even higher temperature excursion.
Although the newly developed drying techniques can facilitate fast and uniform drying, they have several limitations. To enable the ambient or, in some cases, refrigerated storage, it is necessary to achieve an even lower end moisture content than the levels investigated by the above studies. A lower moisture content would demand more power, longer drying time, or a smaller sample volume, which could result in detrimental damages to the cells or reduce the overall throughput.
Although the abovementioned drying techniques can produce a uniform glass that may be able to arrest molecule motions mechanically, our understanding about cellular and molecular injuries associated with these processes is still incomplete.
Besides, most of the drying methods discussed previously currently target samples of microliters, posing a scaling-up challenge if they are going to be adapted to fit biomanufacturing that always prefers large-scale batches or continuous processing.
Summary
By “suspending animation” at cryogenic temperatures, such as in the vapor phase of liquid nitrogen, cryopreservation is currently the main biobanking approach to maintain the safety and potency of cell therapies during transportation and storage. In comparison with cryopreservation, preservation of cell products in a dry state could avoid cryogenic logistics, thereby expanding patient access to these emerging therapies and reducing the overall cost for patients.
This article reviews the current efforts on freeze-drying or drying of mammalian cells. It discusses the existing protection strategies that have been shown to be able to improve the viability and/or functionality of cells after preservation under water-deficit conditions, as summarized in Table 1. Further understanding of the biology of freeze-drying and drying of mammalian cells, continued innovation in cryo- and lyoprotective strategies, and improvement of current techniques' scaling-up capability and compliance with current good manufacturing practice could ultimately enable the preservation of cell therapies in a dry state, bringing both economic and practical benefits to the patients, health care providers, and biopharmaceutical companies.
Technologies Enabling Freeze-Drying or Drying of Biologics and Their Applications, Benefits and Challenges
cGMP, current good manufacturing practice; LEA, late embryogenesis abundant.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The author received no financial support for the research, authorship, and/or publication of this article.