
Abstract
Background:
Sperm-associated antigens (SPAGs) are 18 types of proteins, some of which play important roles in various biological functions associated with assisted reproductive technology outcomes, and are consequently important to the success of fertility programs. Despite the favorable outcomes of fecundity rates among male patients with cancer using cryopreserved sperm, the detrimental impact of freezing on cells has been noted in many studies. Cryopreservation has been thought to have adverse effects on sperm quality through disruptions in the expressions of SPAG genes. This study aimed to evaluate the effects of cryopreservation on the expressions of SPAGs genes and their transcriptome alterations in human sperm.
Materials and Methods:
A total of 12 normal ejaculations were prepared using the density gradient centrifugation procedure, and the motile sperm fractions were divided into fresh and frozen groups. In the latter, sperm samples were mixed with SpermFreeze® solution as the cryoprotectant. The cryovial of sperm suspension was first held just over nitrogen vapor and then dipped inside liquid nitrogen. After 3 days, the specimens were thawed in tap water and incubated for 2 hours for recovery. Then, RNA from sperm was extracted for SPAG gene expression analysis, using real-time polymerase chain reaction.
Results:
Our findings showed a decrease in expression of SPAG5 (p-value = 0.009), SPAG7 (p-value = 0.004), and SPAG12 (SNU13/NHP2L1; p-value = 0.039) genes during cryopreservation.
Discussion:
The results indicate that the freezing procedure could negatively affect gene expression and to some extent proteins in human spermatozoa.
Conclusion:
The alteration of SPAG expression could provide new information on the molecular correlation between cryopreservation and increased failure in intracytoplasmic sperm injection and in vitro fertilization.
Introduction
Cryopreservation is used for long-term storage of biomaterials, living cells, and tissues at liquid nitrogen temperatures (∼−196°C) to preserve their structures and functions. Cryopreservation of sperm is a mature technology now and is a successful component of assisted reproductive technology (ART). 1 Despite its widespread use, the cryopreservation technique can cause damage to sperm through induction of DNA alterations, fragmentation, and oxidation, 2 and is known to affect the motility, mitochondrial activity, and morphology of sperm.3,4
Human sperm-associated antigens (hSPAGs) include 18 types of proteins and some of them (SPAG1, 5 SPAG2, 6 SPAG6, 7 SPAG8, 8 SPAG99 SPAG12, 10 SPAG13, 11 and SPAG15 12 ) play significant roles in fertility. Their molecular weights range between 24 and 71 kDa. 13 SPAG isoforms are encoded by genes present at specific locations on different chromosomes.
SPAG proteins have critical roles in cells; they are involved in cytoplasmic assembly of the ciliary dynein arms (SPAG1), 14 signaling receptor binding (SPAG13), 15 sperm flagellar function (SPAG6, 16, 17)7,16 sperm-egg adhesion (SPAG15, 10),17,18 antimicrobial activity (SPAG11), 19 spermatogenesis (SPAG4), 20 nucleic acid binding (SPAG7, 12), and microtubule-binding and cell division (SPAG5, 8, 9).21–23 SPAG1, 2, 3, 4, 5, 6, 8, 9, 13, 16, 17, and 18 are found in the cytoplasm. SPAG7, 8, 12, and 18 are localized in the nucleus and SPAG10, 11, and 15 are found in cellular membranes (Uniprot; GeneCards).
Recently, the role of tumorigenicity in breast cancer has been attributed to SPAG5, 6, 9, and 10.6,24–26 SPAG1 is associated with pancreatic cancer, 27 SPAG8 and 9 are implicated in cervical cancer28,29 and SPAG17 is a testicular cancer biomarker. 6 Recent evidence has shown that SPAG4 is a prognostic factor in the progression of clear-cell renal cell carcinoma and glioblastoma.30,31 Wang et al. have shown that the proliferation of tumors in renal cell carcinoma can be suppressed using microRNA-200a-3p by targeting SPAG9. 32 SPAG3 and SPAG18 were first identified by Bohring et al. through two-dimensional polyacrylamide gel electrophoresis and matrix-assisted laser desorption/ionization mass spectrometry, and were reported to be heat-shock protein A2 and proteasome subunit α5, respectively. 13
Antisperm antibodies (ASAs) are proteins that lead to humoral immune response and immunological infertility through disruption of sperm fertility. Immunological infertility is caused by the binding of ASAs to the sperm membrane and consequent inhibition of sperm functions. SPAG can be recognized through their binding to these antibodies. 13 In addition, SPAGs are protein factors that are directly transmitted down generations of sperm and act at various stages, not just during embryogenesis, but in adults as well. 33
The term “transmitted down generations” has come to be used to refer to SPAGs present in most tissues, including lung, brain, colon, kidney, testis, placenta, liver, pancreas, thymus, peripheral blood leukocytes, prostate, ovary, spleen, somatic tissues, caput and proximal corpus of epididymis, epididymal epithelium, and sperm surface (Uniprot). Therefore, cryoinjuries to sperm can influence the development of future fetus and offspring derived from cryopreservation.
There have been many studies on freezing of human sperm, and only one of them has mentioned changes in one SPAG protein after the freeze–thaw process. 34 Owing to the importance of SPAGs and their critical roles in fertility, such as the roles of SPAGs4, 6, 16, and 17 in sperm tail motility, those of SPAG3, 10, and 15 in sperm-egg crosstalk, and that of SPAG3 in capacitation, for the first time, we analyzed the effects of cryopreservation on the expressions of all hSPAGs (hSPAGs 1–18).
Materials and Methods
Chemicals and ethics
All chemicals used in the study were procured from Sigma (St. Louis, MO) unless otherwise mentioned, and all techniques were part of clinical laboratory procedures.
Informed written consent was obtained from all participants for the use of their semen samples, and clinical and biological data for research purposes. All experimental protocols were approved by Royan Institute Research Ethics Board (REB) for data records (IR.ACECR.ROYAN.REC.1397.231). All methods were carried out in accordance with the ethical guidelines of the Helsinki Declaration of 1975, as revised in 2008.
The project was found to be in accordance to the ethical principles and the national norms and standards for conducting Medical Research in Iran.
Patient selection and semen collection
This study was performed with sperm from 12 normozoospermic patients who were referred to the Royan Infertility Clinic Center (Tehran, Iran). Semen samples were obtained through masturbation into sterile plastic containers after 3–5 days of sexual abstinence and left to liquefy at 37°C. The exclusion criteria for patient selection were leukocytopenia (≤1 × 106 white blood cells/mL), consumption of alcohol, use of cigarettes, and exposure to radiation or chemotherapy. After liquefaction of samples at 37°C for 30 minutes, basic semen analysis was performed according to the 2010 World Health Organization guidelines. 35 Acceptable samples had the following criteria: normal viscosity and morphology >4%, total motility >60%, 100 × 106 < sperm count <120 × 106 sperm/mL.
Study design
After primary semen analyses, two aliquots of each sample were further processed as follows: in the first stage, both aliquots in each sample were subjected to AllGrad (Lightglobal) gradient centrifugation to isolate the living spermatozoa. Then, the first aliquot was subjected to RNA extraction for real-time analysis and another sample was frozen in sperm freeze medium and thawed after 3 days. 36 After thawing, the process of real-time analysis was performed for cryopreserved sperm, as was performed on fresh sperm.
Density gradient centrifugation
To remove immotile spermatozoa, germ cells, leucocytes, and epithelial cells from the ejaculates, the Royan human sperm biology laboratory protocol was followed, and an AllGrad density gradient centrifugation (DGC) method was applied. Since spermatozoa are denser than the other floating cells, they settled at the bottom of the centrifuge tube. The precipitated spermatozoa were isolated using two-layer AllGrad DGC (45%–90%), diluted in human tubal fluid (HTF) medium containing 5% human serum albumin (HSA; Sigma) and centrifuged at 1800 g for 15 minutes in 15 mL conical tubes. 37 After this, the pellet was resuspended in 3 mL of HTF with 5% HSA and centrifuged at 300 g for 5 minutes. Phenol was removed using 1 mL phosphate-buffered saline and centrifuged for 5 minutes at 300 g to avoid phenolic contamination in RNA extraction.
Sperm cryopreservation and thawing procedure
All sperm samples used in this study (n = 12), were cryopreserved following the protocol used at Royan Institute Infertility Clinic. The cryopreservation procedure was performed according to the manufacturer's directions for rapid freezing using Sperm Freeze (Fertipro, Beernem, Belgium) as the cryoprotectant agent (CPA). One milliliter of each sample was diluted (1:0.7) with CPA in a cryotube (Nunc International, Roskilde, Denmark) for storage. After equilibration (10 minutes) at room temperature, the sample was exposed to liquid nitrogen vapor for 15 minutes, and then plunged into and preserved in liquid nitrogen (−196°C) for 3 days.
The frozen specimens were thawed for 5 minutes under running tap water. Then the cryoprotectant was removed by gradual addition of 4 mL HTF medium supplemented with 5% has followed by centrifugation (2000 g, 5 minutes). For recovering sperm samples for genome reactivation, thawed sperms were retained at 37°C under 5% CO2 38 for 2 hours, after which the RNA was extracted.
The optimal recovery time point after freeze–thawing of human spermatozoa
For evaluating the optimal recovery time point for spermatozoa after thawing, semen samples were collected from four normozoospermic individuals. The aliquots of samples were split into three equal volumes, including one fresh and two frozen groups—the two frozen groups were treated to different incubation periods as described hereunder. After 3 days of storage of the “frozen groups” sperm cells in liquid nitrogen, the samples were thawed and then incubated in a CO2 incubator. One of the frozen groups was incubated for 1 hour 38 and the other for 2 hours for recovery. Then gene expression of SPAGs1, 2, and 12 in fresh and freeze–thawed sperm samples were evaluated using real-time polymerase chain reaction (PCR) technique (Applied Biosystems, Foster City, CA). These specific SPAGs were chosen because of their roles in the signaling pathway, biosynthesis, and pre-mRNA splicing.
RNA extraction
TRIzol® reagent (Invitrogen) was used according to the manufacturer's instructions to isolate total RNA from the sperm. The quantity and quality of isolated RNA were evaluated using spectrophotometry (NanoDrop 1000; Thermo Fisher Scientific). The spectrophotometer optical density ratio for all total RNA samples was 260/280 > 1.8 and 260/230 > 2. 39
Validation through quantitative real-time PCR
cDNA was synthesized by reverse transcription (PrimeScript RT Reagent Kit; Takara, Shiga, Japan) and amplified through real-time quantitative PCR (qPCR) with the primers, shown in Table 1. Exon spanning primers for qPCR were designed for selected genes using Perl Primer Version 1.1.21 and Gene Runner Version 6.00 (Hasting Software, Inc., Hasting, NY) software. Specificity of each primer pair was confirmed using the Nucleotide Blast and Primer-BLAST online software. Experiments of qPCR were performed in triplicates, with duplicate samples and the β ACTIN housekeeping gene. Samples were normalized against the expression of β-actin as the internal control.
Primers Used for Appraisal of Expression in SPAG Genes with Quantitative Polymerase Chain Reaction
HSPA2, heat-shock protein A2; PSMA5, proteasome subunit α5; SPAG, sperm-associated antigen.
The assays were performed in 96-well plates (Applied Biosystems). Each reaction mixture (10 μL) contained 2.5 μL of SYBR Green PCR (Applied Biosystems), 1 μL of each forward and reverse primers, 2 μL of cDNA, and about 3.5 μL of bi-distilled water up to the final 10 μL volume. qPCR assays for fresh (control) and cryopreserved (test) samples for each donor were concurrently performed in the same plate and the dissociation curves were drawn to affirm the specificity of the products. The parameters used for the amplification were 95°C for 10 minutes; and subsequently at the annealing temperature, 40 cycles of 95°C for 15 seconds and 60°C for 1 minute were carried out. Analysis of the melting curve was performed to ascertain that only one peak was present per reaction; and this was confirmed by running 2% agarose gel electrophoresis of the PCR products. 40
Statistical analysis
The data were presented as mean ± standard error of the mean and p ≤ 0.05 was considered significant. Statistical analysis was performed using SPSS version 20 (SPSS, Chicago, IL). The Kolmogorov–Smirnov test was applied to assess normal distribution of values. The p-value was determined by one-way analysis of variance (ANOVA) and independent Student's t-test. Real-time PCR data used for assessment of gene expression were expressed by the ΔΔCt method with the following formula: ΔΔCT = ΔCttest−ΔCtref.
Results
Characterization of optimal recovery time point for cryopreserved sperms after thawing
SPAG1, SPAG2, and SPAG12 were considered for the determination of the optimal incubation time for recovery of the sperm cells after the freeze–thaw process. These specific SPAGs were chosen for this evaluation as they are involved in genomic functions, including the cellular signaling pathways (Uniprot). Ergo, changes in expression of these genes can be a good indicator for determining the optimal recovery time. To assign the optimal recovery time after the freeze–thaw process in human spermatozoa, 1- and 2-hour incubation times were considered (Fig. 1A–C). Statistical analysis revealed no statistically significant differences between the two recovery time periods (1- and 2-hour times); nonetheless the expression of SPAG-genes was slightly higher after the 2-hour incubation compared with the 1-hour case. In this regard, the following recovery period was evaluated at 2 hours after the thawing procedure.
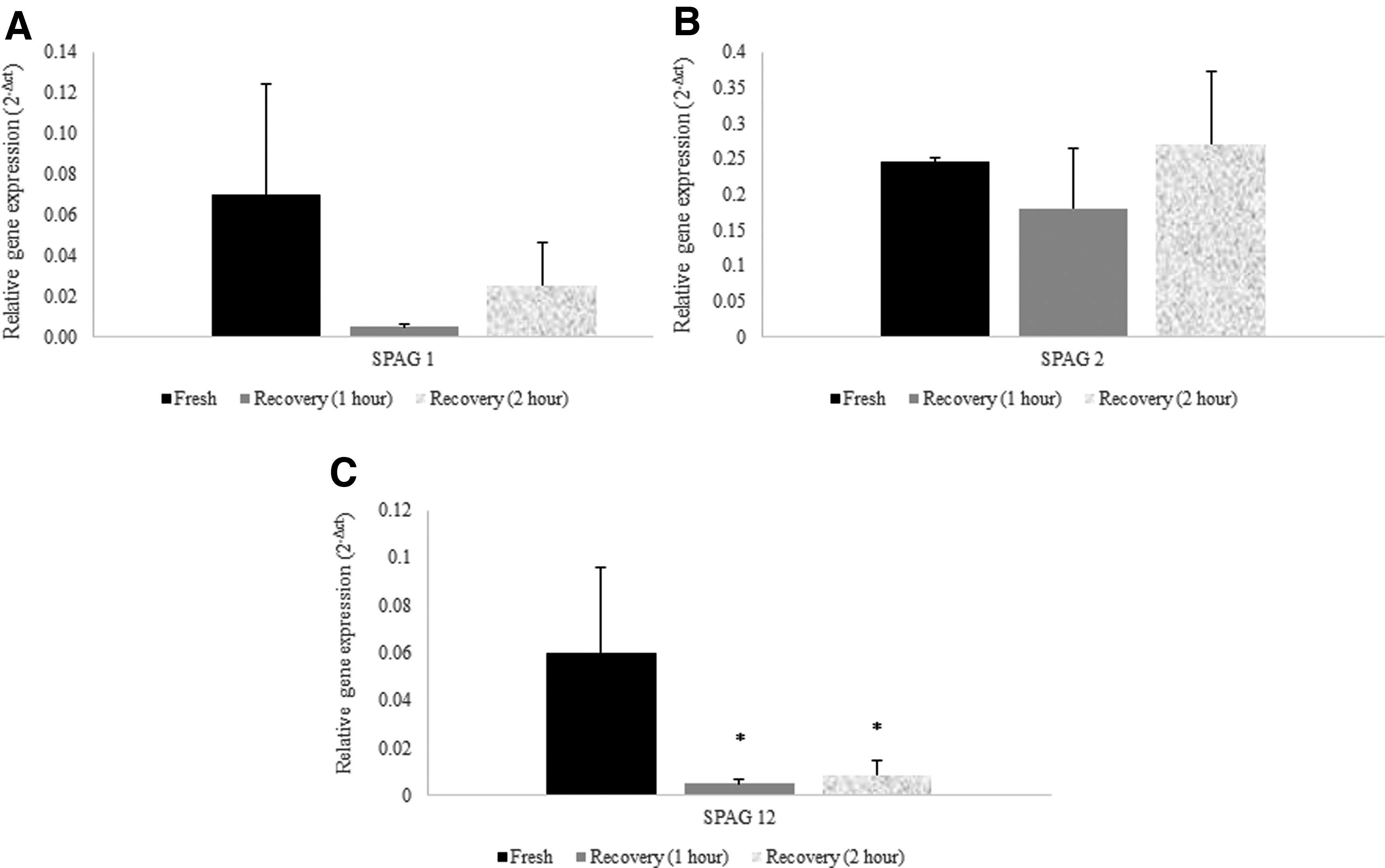
Data comparison of gene expression among fresh and freeze groups (1- and 2-hour times). Relative gene expression of SPAG1, SPAG2, and SPAG12 in human spermatozoa.
The effects of the cryopreservation procedure on sperm characteristics at two different recovery periods were evaluated using computer-assisted sperm analysis before and after freezing (Fig. 2A–G). The results, as shown in Figure 2, indicate that there is a significant decrease in total motility (Fig. 2A, p-value = 0.000) and progressive motility (Fig. 2B, p-value = 0.001) for the samples with 1- and 2-hour recovery periods, compared with the fresh group.

Effects of freeze/thaw process in two different recovery time points on motility parameters of normal human spermatozoa. Data are expressed as mean ± SEM (n = 12; p < 0.05, one-way ANOVA, post hoc Tukey test) for
Quantitative expression of SPAGs1–18 genes in spermatozoa after freeze–thaw process
Analysis of data showed significant reduction in gene expression of SPAG5, SPAG7, and SPAG12 genes in the frozen group compared with fresh (nonfrozen) group (p ≤ 0.05; Table 2). It is apparent from Table 2 that the expression levels of SPAG5 (p-value = 0.009), SPAG7 (p-value = 0.004), and SPAG12 (p-value = 0.039) were remarkably decreased.
Value of Expression Level in Each SPAG Genes in Fresh and Frozen Human Sperms
Data were presented as mean ± standard error of the mean, p ≤ 0.05.
Discussion
Many studies have shown that freezing of sperm affects gene expression due to the detrimental effects of low temperature.41,42 The purpose of this study was to determine if the freezing procedure has an adverse effect on the expression of human SPAG genes during sperm cryopreservation. SPAGs are important proteins and play roles in cell signaling, fertilization, sperm-egg adhesion, integrity of sperm tail, fertility, and motility in human sperm cells. The immunological role of the 18 types of SPAG proteins that stimulate the immune system by binding to ASAs, can lead to infertility. 13 These genes are expressed not only in sperm cells, but also in other tissues and any possible damage to these genes can potentially lead to diseases such as cancer in some tissues. 6
There have been, to the best of our knowledge, no studies on cryopreservation effects on the expression of SPAG genes in human sperm (hSPAGs 1–18). This reported research study was aimed at understanding the effects of the cryopreservation procedure on expression levels of these valuable proteins.
The results showed that a significant decrease occurs in the expression of SPAG5, 7, and 12 in frozen sperm. SPAG5 is an indispensable part of the mitotic spindle for anaphase entry. It is also involved in mitotic progression and regulation of separase activity (UniProt ID Q96R06) and plays a role in the embryonic development of the testis. 43
SPAG7 is a nucleic-acid binding protein (UniProt ID O75391). Recent evidence suggests that SPAG7 may be a possible structural element of sperm acrosome. 44
SPAG12 is a highly conserved nuclear protein that is a part of the (U4/U6. U5) tri-snRNP that binds to the 5′ stem-loop of U4 snRNA. It also contributes to pre-mRNA splicing as part of the spliceosome (UniProt ID P55769). SPAG12 has a role in fertilization and also functions in sperm tail elements and postacrosomal zone. 45 Adjustment disorder in this gene has been seen in some forms of cancer.46,47
In 1995, Mazzilli et al. reported that during cryopreservation, reactive oxygen species (ROS) are produced and this leads to a reduction in sperm motility and viability, and also has serious effects on the vital activities of cells. 48 Different types of oxidative stress affect DNA integrity by causing several clefts, cross-links, nitrogen base modifications, and even DNA strand breakage. 49 There have been relatively few recent studies on the reduction of DNA and RNA-binding proteins and their destruction due to oxidative stress and free radicals produced in the freezing process.50,51
Oxidative stress due to freezing has a powerful impact on increasing microRNAs and consequently regulates cell activity. It has been reported that microRNAs can regulate the expression of genes through various signaling pathways. 52 As noted by Guo et al., acute cold stress conditions (4°C) affect the upregulation of microRNAs in some species. 53 Zhang et al. report that miR-539 directly affects and downregulates the expression of SPAG5. 54 Therefore, it can be said that the oxidative stress after cold shock possibly increases miR-539 and downregulates SPAG5 expression.
Significant reduction of SPAG genes was observed as a result of the freezing process. Lee et al. and Roy et al. reported reduction of RNA and DNA binding proteins due to effects of ROS and destruction of these proteins by free radicals in some cases.50,51 It is possible that oxidative stress, generated by the freezing process, reduces the expression of SPAG7 and SPAG12.
Considering that SPAG12 is an important protein associated with fertility, its significantly lowered expression during the freezing process may have a negative effect on sperm fertility.
Bogle et al. analyzed the data from three individuals and concluded that SPAG17 increased in human sperm protein content after cryopreservation. 34 However, in this study, expression of SPAG17 decreased, but there were no significant differences between the two groups. The differences in the cryopreservation-thawing methods could be a reason for the discrepancy between the two studies. In addition, inducing more (or less) oxidative stress compared with another cryopreservation method can be effective as well. The other possible reason for this discrepancy could be differences in sampling. Bogle et al. examined three semen samples, whereas in this study 12 semen samples were examined. It seems that at higher sample size, individual variations are eliminated, and the results are more homogenic and reliable. The last possible explanation for inconsistency in results may be the fact that in the study by Bogle et al. protein changes were studied, whereas we evaluated the gene expression.
According to Ward 55 and Johnson et al., 56 histone-associated chromatin regions have ranged from 10% to 15% in mature spermatozoa and are poised for expression. In consonance with the strands of RNAs preserved in the mature spermatozoa, low amounts of nucleosomes present in histone-associated parts may synergistically interact to intuit primary embryonic gene expression. 56 After the thawing procedure, the sperm cells were allowed to resume their gene expression within a 2-hour recovery period, and the results showed that the expression of the target genes was better after 2 hours compared with 1 hour. Thus, it can be inferred that gene expression resumes in sperm cells, after they are thawed, and the SPAGs are expressed in mature sperms.
It is concluded that the expression of SPAGs are reduced when the temperature is reduced. The expression of SPAGs5, 7, and 12 are significantly reduced during the sperm freezing procedure. The evidence presented thus far supports the idea that cryopreservation and probable ROS may affect the histone-associated nucleosomes area. Such alterations in expression levels may have serious effects on the sperm's functions. Since these genes and derived proteins play important roles in fertilization, cleavage, implantation, fetal life, embryonic development, and possibly even postnatal lifespan, the reductions in expression may affect ART success.6,43,57
Further study needs to be done to evaluate the possible mutations of SPAG genes in cryopreserved sperms. It would be interesting to assess the effects of freeze–thawing procedure on embryos and fetuses derived from cryopreserved sperms.
Footnotes
Acknowledgments
The authors thank Mr. Vahid Esmaeili and Mrs. Elham Abedheydari for their laboratory support at Royan Institute.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
No funding was received for this article.