
Abstract
Freeze-drying is one of the sperm preservation methods leading to the long-term preservation of sperm genetic material. Our main goal of this study was to evaluate the effect of the trehalose freeze-drying method on sperm motility, viability, morphology, acrosome, and DNA integrity compared with a standard protocol without trehalose. Twenty-five normozoospermic samples were included in this prospective study. Direct swim-up was used for sperm preparation. An experiment was performed on freeze-dried samples containing trehalose (0.2 M), and the results were compared to that without trehalose. The sperm parameters, including count, motility, morphology, viability, acrosome reaction, DNA denaturation, and DNA fragmentation, were evaluated before and after freeze-drying in both groups. The spermatozoa were totally immotile after freeze-drying in both groups. Sperm viability, acrosome integrity, and nondenatured sperm DNA were significantly higher in the trehalose group in comparison with that of without trehalose group. Nonfragmented sperm DNA showed an increasing trend in the trehalose group compared to the group without trehalose. While freeze-drying significantly reduced normal morphology, the addition of trehalose did not affect this parameter. The results of this study showed that trehalose can attenuate the detrimental effects of freeze-drying on human sperm parameters.
Introduction
Sperm cryopreservation is routinely performed worldwide in in vitro fertilization clinics. One of the limitations of sperm cryopreservation methods is the need for a permanent source of liquid nitrogen. It is difficult to transport liquid nitrogen from production centers, and liquid nitrogen containers may be broken or contaminated. 1 Sperm freeze-drying has been introduced as a method of preserving the genetic source. 2 Freeze-drying does not require liquid nitrogen or dry ice and is an option for the preservation of viral samples and can omit the risk of cross-contamination. In addition, it is less expensive to maintain the freeze-dried sperm cells at room or ambient temperature, making the transfer of freeze-dried sperm cells from one place to another easier, and the cells are easily rehydrated.3–6
In this method, the integrity of the genetic material could be preserved for a period of more than 1.5 years at 4°C. 4 It was recently shown that freeze-drying could be considered as a sperm preservation method to produce genome-edited animals. 7 Despite the advantages, damage to the sperm membrane and thus reduced viability are among the limitations of this procedure. While freeze-drying has been studied in human spermatozoa, the sperm cells became immotile and died after freeze-drying in all the media. Damage to the sperm membrane has been proven by electron microscopy studies as well. 8 More studies should be performed to modify the freeze-drying method for permanent and successful preservation of human dried spermatozoa.
It would be worthwhile to develop a new freeze-drying buffer medium for better results. This medium must be adapted to the characteristics of different species and improving the composition of the buffer or adding an antioxidant can be effective. Trehalose is a natural sugar that contains two glucose molecules and is an impermeable cryoprotectant. Its presence on both sides of the plasma membrane is required as an effective cryoprotectant. 9 This molecule maintains proteins and membranes in a physical state, as in the case of complete dehydration, 10 prevents crystallization, and provides stability during drying by reacting with biological structures. 9
One reason for the decrease in sperm quality is the crystallization of intracellular ice during cryopreservation, which is minimized in isotonic environments with nonreducing sugars. 11 Due to this dehydration, the volume of water inside the cell decreases and therefore there will be less intracellular ice formation, which is beneficial for sperm because intracellular ice formation causes cell death and reduces fertility.12,13 The protective effects of trehalose on DNA integrity of boar freeze-drying sperm have been shown previously. 14 In addition, it was shown that evaporative drying of the mouse spermatozoa using trehalose maintained at room temperature for 2 years has the potential to produce live offspring. 15 Recently, Wakayama et al. showed that trehalose improves the tolerance of freeze-drying mouse spermatozoa to extreme temperatures. 16
To the best of our knowledge, this is the first study that evaluates the effect of trehalose in freeze-drying of human spermatozoa on sperm motility, viability, morphology, and integrity of acrosome and DNA in comparison with standard protocol.
Materials and Methods
Chemicals
All the reagents used were purchased from Sigma (Sigma-Aldrich).
Samples
Twenty-five normozoospermic specimens were included in this prospective study. The semen samples were collected by masturbation after 3–5 days of abstinence. The sperm parameters were evaluated according to WHO guidelines. 17 Sperm preparation was done by the direct swim-up method. The sperm samples were divided into two groups of trehalose and without trehalose, and freeze-drying was performed. In this study, the effect of 0.2 M trehalose on sperm parameters after freeze-drying was estimated. 2 This study was approved by the Institutional Review Board (IRB) and Ethics Committee of Tarbiat Modares University (IR.MODARES.REC.1397.262).
Freeze-drying
Freeze-drying was performed using a modified standard protocol, as previously described. 18 First, after swim-up, 100 μL of the sperm pellet (70 × 106 mL−1) was resuspended in a 400 μL buffer containing 1 mM ethylene diamine tetraacetic acid (EDTA) and 10 mM Tris-HCl at room temperature. Then, it was packed in cryovials of 2 mL before being transferred to a freezer of −20°C and −80°C for 6 and 24 hours, respectively. After that, the samples were incubated in a precooled freeze-dryer chamber (Alpha 1–4 LSC; Martin Christ, Germany) with a cycle at a condensation temperature of −50°C and 0.22 bar pressure for 48 hours. The lyophilized samples were stored in vacuum-sealed bags for 1 month at 4°C and then rehydrated through resuspension in 100 μL of deionized water at room temperature for further analysis. 19 In the trehalose group, 0.2 M trehalose was added to freeze-drying buffer, and all stages were the same.
Analysis of sperm characteristics
Sperm motility, viability, and morphology
Sperm motility was assessed based on the WHO classification. 17 In this report, sperm that were actively moving linearly or in a large circle, regardless of velocity, were classified as progressive. All other patterns of motion with a lack of progress, for example, swimming in a small circle, belonged to sperm with nonprogressive motility. Immotile spermatozoa did not have any moves. At least 200 spermatozoa at six fields were evaluated by two expert operators. Membrane integrity and sperm viability were evaluated by the eosin-nigrosine staining method. 17 The sample was well mixed and inserted into the microtube; the equivalent volume of eosin-nigrosine dye was added. After 30 seconds, 10 μL of the mixture was removed and a smear prepared. At least 200 sperm cells were evaluated (1000 × ).
Live sperm cells were identified with white heads, and spermatozoa with red or dark pink heads were considered as dead sperm cells. Sperm morphology was evaluated with the Papanicolaou staining method. 17 A smear was prepared on the slide and, after being dried, immersed in 90% ethanol solution for cell fixation and dehydration. The slides were then resuspended in 70% ethanol and distilled water to enter the water-soluble hematoxylin dye. This color makes the core blue. For decolorization of the cytoplasm, slides were immersed in acidic ethanol 4 to 8 times. To reduce the acidity, slides were placed in running water and subsequently they were placed in 70% and 90% ethanol, respectively. This dehydration allows the Orange G/EA50 dye to enter the ethanol solution.
The slides were again resuspended in 90% ethanol for EA50 green staining, in which this color, along with the previous color, resulted in a pink cytoplasm. Abnormalities in the head, including shape and size of the head, vacuoles, number of heads, size of acrosome, and abnormalities in mid piece, including thickness, insertion, excess residual cytoplasm, and abnormalities in the tail, including length, number, shape, and excess residual cytoplasm, were checked with brightfield light microscope at × 1000 magnification with oil immersion. At least 200 sperm cells were checked for head, mid piece, and tail abnormalities.
Acrosome reaction by double staining
To fix the sample, 3% glutaraldehyde was mixed with an equal ratio of sperm. After centrifugation, the pellet was mixed with distilled water. The smear was prepared and dried. The staining was performed with Bismarck brown (37°C, 10 minutes) and Rose Bengal (room temperature, 20 minutes). At least 200 spermatozoa were evaluated for acrosome reaction (1000 × ). Sperm with brown or light pink heads did not respond to acrosome reaction considering intact acrosome. Those with darker or conical heads had lost their acrosomes. 20
DNA denaturation by acridine orange test
The smear was first prepared from the sample and then was immersed in Carnoy's solution (methanol:glacial acetic acid, 3:1) for 2 hours at 4°C. Acridine orange was added in the darkroom and observed by fluorescent microscope and a 460 nm filter. At least 200 spermatozoa were examined (1000 × ). Sperm cells with healthy DNA were seen in green, and the denatured sperm cells were seen in yellow to red. 20
Sperm chromatin dispersion test
The sperm sample was mixed with low melting agarose and was placed on a slide already covered with normal agarose. After putting on a coverslip, it was incubated for 5 minutes at 4°C. Then the slide was kept in 0.08% HCl in the dark for 7 minutes. The slide was immersed in lysis buffer (distilled water, sodium dodecyl sulfate, NaCl, Tris-HCl, EDTA, and mercaptoethanol) for 25 minutes, which leads to the breakdown of disulfide bonds and the removal of proteins. After being washed with distilled water for 5 minutes and dehydrated by 100%, 90%, and 70% alcohols, it was dried. Finally, it was dyed in Wright's solution. Thus, the outer DNA loops and halo formed around the nucleus. The dispersion of these loops and the amount of halo indicated the extent of damage. At least 200 spermatozoa were evaluated (1000 × ). The spermatozoa with medium and large haloes were considered normal, and spermatozoa with a small halo and no halo were considered abnormal cells. 21
Statistical analysis
The data were reported as mean ± standard deviation and median (maximum-minimum). The Shapiro–Wilk test was used to measure the normative distribution of data. Kruskal–Wallis test with Dunn's multiple comparison test was used to analyze the data between the experimental groups. Independent-samples t-test and Mann–Whitney U test were used for comparison of different morphological abnormalities between two groups wherever appropriate. p-Value was considered ≤0.05.
Results
The characteristics of semen included in this study are shown in Table 1. The freeze-drying process totally immobilized spermatozoa in both groups. In addition, sperm normal morphology was reduced after freeze-drying, and addition of trehalose had no effect on preserving the normal morphology. Addition of trehalose improved sperm viability, acrosome, and DNA status in freeze-dried sperm. Sperm viability decreased after freeze-drying in both groups, but in the presence of trehalose, a significant increase was observed compared to the freeze-drying without trehalose. The rate of intact acrosome and nondenatured DNA after freeze-drying was significantly increased in the trehalose group compared to the without trehalose group. The DNA fragmentation occurred after freeze-drying, and the addition of trehalose was attempted to maintain DNA fragmentation but the difference was not significant (Table 2). In a subanalysis, after rehydration, we compared different morphological defects between two groups. Our data showed that there were no significant differences regarding head, mid piece, and tail abnormalities between the two groups (Table 3). Figure 1 shows sperm viability, acrosome reaction, and DNA fragmentation.
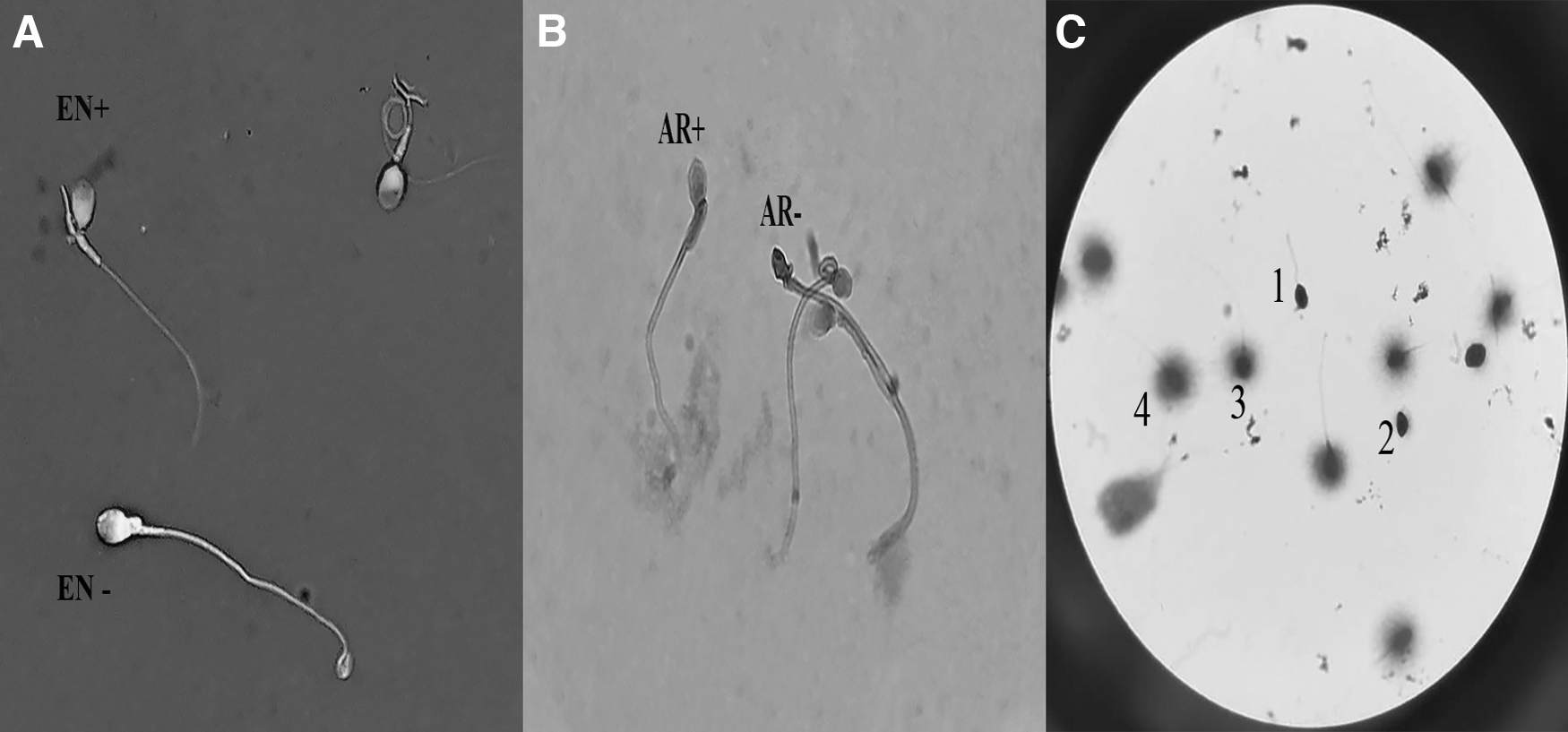
Sperm viability, acrosome reaction, and sperm DNA fragmentation.
Semen Characteristics Included in This Study
SD, standard deviation.
Comparison of Sperm Parameters Between Different Groups
Data as mean (%) ± standard deviation, median (maximum–minimum).
Numbers with the same letters have a significant difference. a,bp ≤ 0.0001; cp ≤ 0.05.
Comparison of Different Types of Morphological Defects After Rehydration Between Two Groups of Freeze-Drying With Trehalose and Without Trehalose
Data as mean (%) ± standard deviation.
Mann–Whitney U test was used.
Discussion
In this study we evaluated the effect of adding trehalose to the freeze-drying solution of human sperm in comparison to a standard protocol. The beneficial effects of trehalose on animal sperm have been previously described, but to the best of our knowledge this is the first study that evaluates the effect of trehalose on freeze-drying of human sperm. Our results showed that the freeze-drying process impaired sperm motility, and addition of trehalose had no effect to maintain sperm motility. In a similar study, freeze-drying caused the overall lack of motility in the rehydrated samples that indicated severe damage to the plasma membrane of the spermatozoa. 19 It was shown that trehalose improves sperm motility after the freeze–thaw process. 22 Trehalose, as a nonpermeable cryoprotectant and a nonreducing disaccharide, extracts the intracellular water leading to reducing the risk of intracellular ice formation. The effect of physical conditions, including cell swelling and shrinkage and chemical environment in freeze-drying, results in damage to the integrity and structure of plasma membrane, which is irreversible for sperm motility. Intracytoplasmic sperm injection must be used for rehydrated sperm, so motility is not important to be maintained after freeze-drying. As the distilled water is used for sperm rehydration in freeze-drying, the significant decrease in sperm viability in both groups after rehydration is not surprising which was in line with previous study. 19 Our data (not shown) showed that rehydration with culture media did not change the sperm parameters, concluding that the sperm damage had already happened (i.e., in the drying steps). According to our data, in the presence of trehalose, sperm viability was significantly higher in comparison with the standard protocol with no trehalose (61.6 ± 27.64 vs. 4.26 ± 5.72, respectively) (Table 2). The freeze-drying of human spermatozoa showed a general lack of motility and viability in human spermatozoa, indicating that the cell membrane was severely damaged by this process. 19 According to El-Sheshtawy et al., using 50–100 mM trehalose maintains bull sperm viability after thawing. 23 Due to the lack of a specific carrier of trehalose in mammalian cells, trehalose remains in the extracellular space after adding as a cryoprotectant agent. 24 Water replacement theory suggests that the hydroxyl groups of trehalose can replace hydrogen bonding. 9 It was shown that trehalose prevents protein denaturation and inhibits oxidation of unsaturated fatty acids.25,26 It is hypothesized that trehalose penetrates the membrane, and it forms a hydrogen bond with the polar heads of the membrane phospholipids. The interaction between trehalose and head groups of phospholipid bilayers under dehydration conditions decreases the gel to liquid transition leading to a reduction in probable damages during rehydration. 27 In addition, trehalose produces osmotic pressure that probably induces cellular dehydration, reduces the formation of intracellular ice crystals, increases membrane fluidity, and stabilizes the biological structure during drying.9,23,28,29 Indeed, the freeze-drying has two steps of primary and secondary drying. In the primary drying phase, the water is removed from the solid parts and is physically separated, and the sample is converted to a solid phase and the remaining water is removed in the secondary drying phase induced by vacuum. In addition, it is suggested that trehalose sequesters the water molecule around the biomolecules resulting in a decrease in the hydration radius and increase compactness and stability during the process. 30 The normal morphology of freeze-dried sperm in the present study indicated a significant decrease compared to the fresh group; even the addition of trehalose to the freeze-drying buffer failed to maintain the normal sperm morphology. A variety of morphological abnormalities were observed in freeze-dried sperm, of which tail abnormalities accounted for the highest percentage. Gianaroli et al. 19 showed preservation of head morphology without increasing vacuole content compared to the fresh group. There was also a significant increase in tail malformations, and curved tails had the highest proportion of this anomaly. When sperm is returned to distilled water, it leads to torsion and flexion of the tail around the sperm head. In contrast, the change in water content during cell dehydration may lead to torsion of the tail, which is evident in the rehydrated spermatozoa. Indeed, the rehydration process was the same between two groups, and tail coiling was seen in both groups (p > 0.05). More importantly, the rate of head abnormality was the same between the two groups. Our data (not shown) regarding the rehydration of spermatozoa with culture media showed that the rate of head abnormality did not change suggesting that the head morphology does not depend on the osmolality of rehydrated medium. Our results showed that freeze-drying impairs acrosome integrity, but the rate of intact acrosomes was significantly higher in the trehalose group compared to the routine procedure. The acrosome is a part of sperm that is highly affected by freeze-drying in all media regardless of semen status or storage temperature. Martins et al. 31 reported that freeze-drying medium containing trehalose maintains the acrosome integrity of bovine sperm. The protective effect of trehalose on acrosome integrity may be dose dependent. It was also shown that trehalose decreases osmotic sensitivity and inhibits acrosome reaction induced by lysophosphatidylcholine in ram spermatozoa. 32 After freeze-drying of dasyurid marsupial sperm using 0.27 M trehalose, ∼80% of spermatozoa had a normal acrosome. 33 In addition, it was shown that acrosomal contents were retained after freeze-drying of primate sperm using trehalose. 34 Recent findings of the effect of trehalose on tolerance of freeze-dried sperm against extreme temperature suggest that trehalose protects the integrity of acrosomes. 16 Drying induces irreversible damages to sperm critical components that are necessary for motility. Our data showed that using trehalose in freeze-dying could not prevent the damages leading to motility loss but could prevent damages leading to impair viability and acrosome status. It was shown that sperm DNA integrity is maintained after freeze-drying. 19 Our data showed that the DNA integrity was better maintained after freeze-drying in the presence of trehalose. In a similar study, it was shown that adding trehalose has an increasing trend to protect DNA integrity in freeze-dried dog sperm using sperm chromatin dispersion test. 2 However, Ito et al. recently showed that trehalose did not affect the rate of DNA denaturation in zygotes produced from freeze-dried mouse sperm. 35 It was shown that the rate of birth after injection of freeze-dried sperm into oocytes is significantly lower compared to fresh spermatozoa, due to DNA damage. 4 DNA damage has been attributed to mechanical stress, heat shock, ice crystal formation, and action of endonucleases released during freeze-drying and oxidative stress that occurs after rehydration. Using calcium chelators (e.g., EDTA and EGTA) can inhibit the endonucleases that compromise DNA integrity during freeze-drying. 1 Adding trehalose to the medium improves the integrity of boar sperm DNA after freeze-drying but does not promote fertility and blastocyst development. 14 The immediate effect of trehalose on freeze-drying medium has been shown from morphology of dried cells seen by scanning electron microscopy. 36 It seems that trehalose, as a good glass former, molecules bond to DNA and form hydrogen bonds among themselves leading to the formation of a glassy and viscous medium, which prevents the rate of DNA denaturation. 36 Trehalose preserves DNA structure with replacing water and formation of hydrogen bonds with DNA. 30 In addition, the antioxidant capacity of trehalose has been shown in a recent study. 37 Our limitation was the lack of comparison of fertilizing ability of spermatozoa between the two groups. It is suggested that a follow up study should evaluate the effect of trehalose on rates of fertilization and embryo development following freeze-drying of human sperm. In conclusion, trehalose can maintain sperm viability, acrosome, and DNA integrity of human spermatozoa during freeze-drying. The efforts to introduce the optimum protocol for freeze-drying are ongoing and seems that adding trehalose could ameliorate the detrimental effects on freeze-drying on human sperm parameters. It was shown that freeze-drying can inactivate viruses in the biological products and has beneficial effects to inactivate viruses in bone transplantation.38–40 Freeze-drying as an alternative method for preservation of human spermatozoa can be translated to the clinic in the future especially for viral infected specimens. More studies evaluating the clinical outcomes should be performed for verifying this hypothesis.
Footnotes
Acknowledgment
This article is extracted from M.Sc. thesis of Elaheh Shahmoradi. The authors thank Tarbiat Modares University for supporting this project.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by Tarbiat Modares University.