
Abstract
Estradiol is a steroid hormone excreted from the female gonads, mainly during the pre-estrus. However, the potential effects of estradiol are yet to be explored on sperm parameters through cryopreservation. In this study, we supplemented estradiol, 3 and 5 μM, in the goat semen extender and assessed the sperm parameters after a freeze-thawing process. Sperm motility was assessed using the computer-assisted sperm analysis system. Sperm viability and membrane integrity improved using both 3 and 5 μM concentrations of estradiol. The highest rate of progressive motility was observed in the 3 μM estradiol group. However, a higher concentration of estradiol (5 μM) reduced the progressive motility. Then, we were interested to see if the supportive effect of estradiol on sperm motility is mediated through the intracellular concentration of calcium ionophore. We supplemented the semen extender with 1 and 10 mM ethylenediaminetetraacetic acid (EDTA) and showed that 1 mM has no adverse effect on progressive sperm motility. Then, estradiol (3 μM) was supplemented with or without EDTA (1 mM) into the semen extender. Individual EDTA treatment improved the progressive sperm motility compared to the control group. However, in the presence of estradiol, EDTA treatment reduced the progressive motility compared to the individual estradiol group. This indicated a considerable interaction between estradiol and EDTA for progressive sperm motility. Indeed, EDTA reduced the supportive effects of estradiol on sperm cryopreservation parameters. These results indicated that induction of higher progressive sperm motility in response to estradiol is a calcium-dependent process, as the EDTA did completely abrogate the estradiol-mediated effect.
Introduction
Despite significant advances in the process of sperm cryopreservation, the technology still faces some challenges such as reduced sperm motility and survival, damaged structure of sperm plasma membranes or organelles, and premature acrosomal reaction, as well as overproduction of reactive oxygen species (ROS) during the freeze-thawing process. 1
17-beta-estradiol (estradiol) is a steroid hormone excreted from the female gonads mainly during the follicular phase of the estrus cycle. During pre-estrus, gonadotropins improve follicular maturation, resulting in increased secretion of estradiol. In addition, estradiol contributes to male sexual function in various ways, from spermatogenesis to libido and erectile function. 2 Through estrogen receptors and aromatase, which convert testosterone to estradiol, estrogen modulates the spermatogenesis, first through the hypothalamus-pituitary-gonadal axis, then by the Leydig, Sertoli, and germ cells, and finally with the reproductive duct and sperm maturation.2,3 In addition, neurons and astrocytes, which both have information-transmitting functions in the nervous system, contain high levels of aromatase and estrogen receptors.3,4 It has been verified that estradiol plays important roles in the development and programming of the sexual behavior in the early brain. 5 Administration of estrogen through aromatase inhibitors at early embryo development has been shown to effectively improve female-to-male sex reversal in chickens.6–8
Estradiol, in the absence of progesterone, improves the estrous behavior through the hypothalamus-pituitary-gonadaxis. 9 Estradiol has an antioxidant and a protective effect on the isolated mitochondria 10 and improves lipid deposition and antioxidant response in male zebrafish. 11 Physiological concentrations of estradiol reduce the oxidation of low-density lipoprotein and high-density lipoprotein.12–14 It has also been verified that estradiol mediates cardiovascular protection through its antioxidant activity and reducing ROS production in brain stem neurons.15,16 Estradiol also regulates the antioxidant production. 17 The beneficial effects of using antioxidants in semen extender have been reported in several studies on the freezing-thawing process (for review, see Amidi et al. 18 ). Moreover, the estradiol level of human serum has a positive association with the embryo cryopreservation. 19 Apart from the antioxidant activity, estradiol and progesterone can regulate the intracellular level of calcium in spermatozoa. 20 Beneficial effects of progesterone have been verified to reduce sperm kinematics and postpone sperm capacitation.21,22 However, the potential effects of estradiol are yet to be explored for sperm cryopreservation.
Sperm motility is a calcium-dependent process. The introduction of extracellular Ca2+ into the fresh semen has been shown to increase the sperm motility as well as the cellular free radicals. 23 It has been reported that incubation of fresh sperm in a calcium ionophore-containing medium, such as A23187, leads to increased production of ROS. 24 Premature sperm hyperactivation through the freeze-thaw process reduces sperm quality and fertility compared to fresh semen. 25 The inevitable partial or serious damage to the sperm plasma membrane throughout the freeze-thaw process can increase the concentration of intracellular calcium ions. 26 Therefore, reducing calcium ion in the freezing medium can modulate its intracellular concentration. Use of calcium chelators is one approach to prevent the surge of intracellular Ca2+ concentration and increase the sperm quality after freezing-thawing.26,27 In this study, we supplemented estradiol, as a multifunctional agent, in the goat semen extender and assessed the sperm parameters after the freeze-thawing process. To assess possible interaction between estradiol and calcium ionophore on the postfreeze sperm parameters, a combined group of estradiol and ethylenediaminetetraacetic acid (EDTA) was also evaluated.
Materials and Methods
Chemicals
All chemicals used in this study were purchased from Sigma Chemical CO. (St. Louis, MO), unless stated otherwise. The Bioxcell semen extender was obtained from Minitube, Germany. We also used the water-soluble form of 17-beta-estradiol (E4389; Sigma-Aldrich, Germany), which hereafter is designated as estradiol.
Animals
These experiments were conducted at the animal farm of Reproductive Biotechnology Research center, Royan Institute (Isfahan, Iran latitude 32390 N). All practices outlined in the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching 28 were also used for the approval of all experimental procedures as part of the local requirements. The semen samples were collected from four Saanen bucks aged 2–3 years. Bucks were offered a base diet according to small ruminant nutrition system, Cornell University (Version: 1,9,4468).
Experimental groups
First, we optimized the concentration of supplemented estradiol and EDTA in two independent experiments. The initial levels of estradiol and EDTA were chosen based on the literature.28,29 In the first experiment, semen was diluted with the extender supplemented with 0, 3, or 5 μM of estradiol. In the second experiment, the experimental groups comprised 0, 1, or 10 mM of EDTA supplementation into the semen extender. From these two experiments, we selected 3 μM estradiol and 1 mM EDTA, by which the highest rate of progressive motility with minimum adverse effects on the membrane integrity was obtained. Finally, in a completely randomized design, the combined effect of estradiol with EDTA (3 μM and 1 mM, respectively) was compared to groups supplemented individually with these molecules, using the nonsupplemented diluter as a control.
Semen collection, processing, and freezing-thawing
Semen collection and processing were carried out according to Kargar et al. 30 In brief, ejaculates were obtained by artificial vagina from the bucks twice a week in a period of 6–8 weeks, corresponding to the breeding season (April/May, 2018). The samples with 1–2 mL volume from each buck were separately transported at 35°C within 30 minutes to the laboratory for microscopic evaluations. Ejaculates were subjectively observed, and only those with at least 70% motile and 80% morphologically normal sperm were mixed and used for further steps. For freezing, a mixed semen sample was diluted with the extender (∼1:40 ratio) so that a final concentration of 400 × 106 spermatozoa/mL (100 × 106 spermatozoa per 250-μL straw) was achieved. Then the samples were cooled to 4°C over a period of 2 hours, drawn into 0.5 mL French straws (Biovet, L'Agile France), and sealed. Then the straws were kept at 4°C for 1 hour followed by an exposure to liquid nitrogen (LN2) vapor for 12 minutes, plunged into the LN2, and stored until being thawed and used for evaluation of sperm parameters. The thawing step was carried out by plunging the straws into a 37°C water bath for 30 seconds. Immediately after thawing, each sample was evaluated by a well-trained staff at room temperature (RT).
Estimation of sperm motility after freezing-thawing process
Following thawing, sperm motility was estimated according to Kargar et al. 30 Briefly, sperm cells from each of four straws belonging to the same replicate were centrifuged and then the sperm pellet was diluted with fertilization medium (Tyrode's albumin lactate pyruvate medium-Fert-TALP; pH: 7.4, and osmolarity: 280–285 mOsmol/kg) to a final concentration of 1 × 106 spermatozoa/mL. For each sample, 5 μL of sperm suspension was loaded on a prewarmed slide covered by an 18 × 18 mm2 coverslip, and a minimum of 500 sperms were analyzed in at least three different microscopic fields. The motility of spermatozoa was assessed using a computer-assisted sperm analysis (CASA) system (Video Test, Ltd., version Sperm 2.1, 1990–2004, Russia). The setup parameters of CASA comprised 100 (Hz) frame rate and minimum contrast of 50.31,32 Velocity parameters of straight-line velocity (VSL), curvilinear velocity (VCL), average path velocity, and linearity (LIN = VSL/VCL) were measured with a10 × standard objective. Motile spermatozoa were allocated to class A or fast progressive motility, which swim fast in a straight line (if VCL >40 μm/s and LIN ≥60%), and class B or slow-medium progressive motility, which move slowly forward, but in a curved line path, (if 20 < VCL <40 μm/s). 32
Assessment of sperm plasma membrane integrity
Sperm plasma membrane integrity was estimated by the hypo-osmotic swelling test (HOST) based on curled and swollen tails. 33 Immediately after the sperm thawing, a 25-μL sperm suspension was incubated with 200 μL of HOST solution (100 m Osm/L, 57.6 mM fructose, and 19.2 mM sodium citrate) for 30 minutes at RT and 5 μL of homogenized mixture was mounted on a glass slide and covered with a coverslip. The percentage of intact membrane spermatozoa was estimated by recording of at least 200 spermatozoa in more than five different microscopic fields using an inverted microscope (400 × , Olympus, CKX41, Japan). Sperm cells with a functional membrane undergo a water influx and membrane expansion in the hypo-osmotic medium and have a curl tail. 33
Assessment of sperm viability
Sperm viability was evaluated using the eosin-nigrosin staining of freeze-thawed samples. 34 In brief, after thawing of the cryopreserved sperm, 50 μL of diluted semen was mixed with 100 μL of eosin (1%) for 30 seconds and stained with 100 μL of nigrosin (10%) for 30 seconds. One drop (7 μL) of stained semen was loaded on a glass slide and semen smears were prepared, air dried, and assessed at a magnification of 1000 under oil immersion of light microscope (at least 200 sperm). Unstained sperm head was considered alive, while sperm with pink or red color in the head region was considered dead sperm.
Statistical analysis
Each experiment was carried out with 22–25 replicates. The analysis of the normal distribution of attributes was carried out in the online web tool of the R package (https://rdrr.io/snippets/). Using the SAS software, version 9.1, the analysis of variance was carried out for the sperm attributes followed by a Tukey post hoc test for mean comparisons (p < 0.05). The results are reported as mean ± standard error for each experiment.
Results
Estradiol effect on sperm postfreeze parameters
Supplementation of estradiol in the semen extender was carried out (Fig. 1). Sperm viability and membrane integrity improved using both 3 and 5 μM concentration of estradiol (p < 0.05). The highest rate of motile spermatozoa for all classes was observed in the 3 μM estradiol group. However, a higher concentration of estradiol (5 μM) reduced the class A and class A + B (p < 0.05).

Effect of estradiol on goat sperm cryopreservation. The semen extender was supplemented with 3 and 5 μm of 17-beta-estradiol. Plasma membrane integrity and sperm viability were assessed by the HOST and eosin-nigrosin assays
EDTA effect
EDTA inclusion into the semen extender affected the sperm postfreeze parameters (Fig. 2). Both sperm viability and membrane integrity improved by supplementation of 1 mM EDTA compared to the control group and 10 mM groups (p < 0.05). However, inclusion of 10 mM EDTA in the extender media reduced the viability attributes compared to the 1 mM EDTA group, while improving the viability compared to the control group (p < 0.05). Compared to the control group, the use of 1 mM EDTA did not affect the progressive motility (class A, class B, and class A+B) (p > 0.05). However, the class B and class A + B motile sperm were increased in the 10 mM EDTA compared to the control group (p < 0.05).
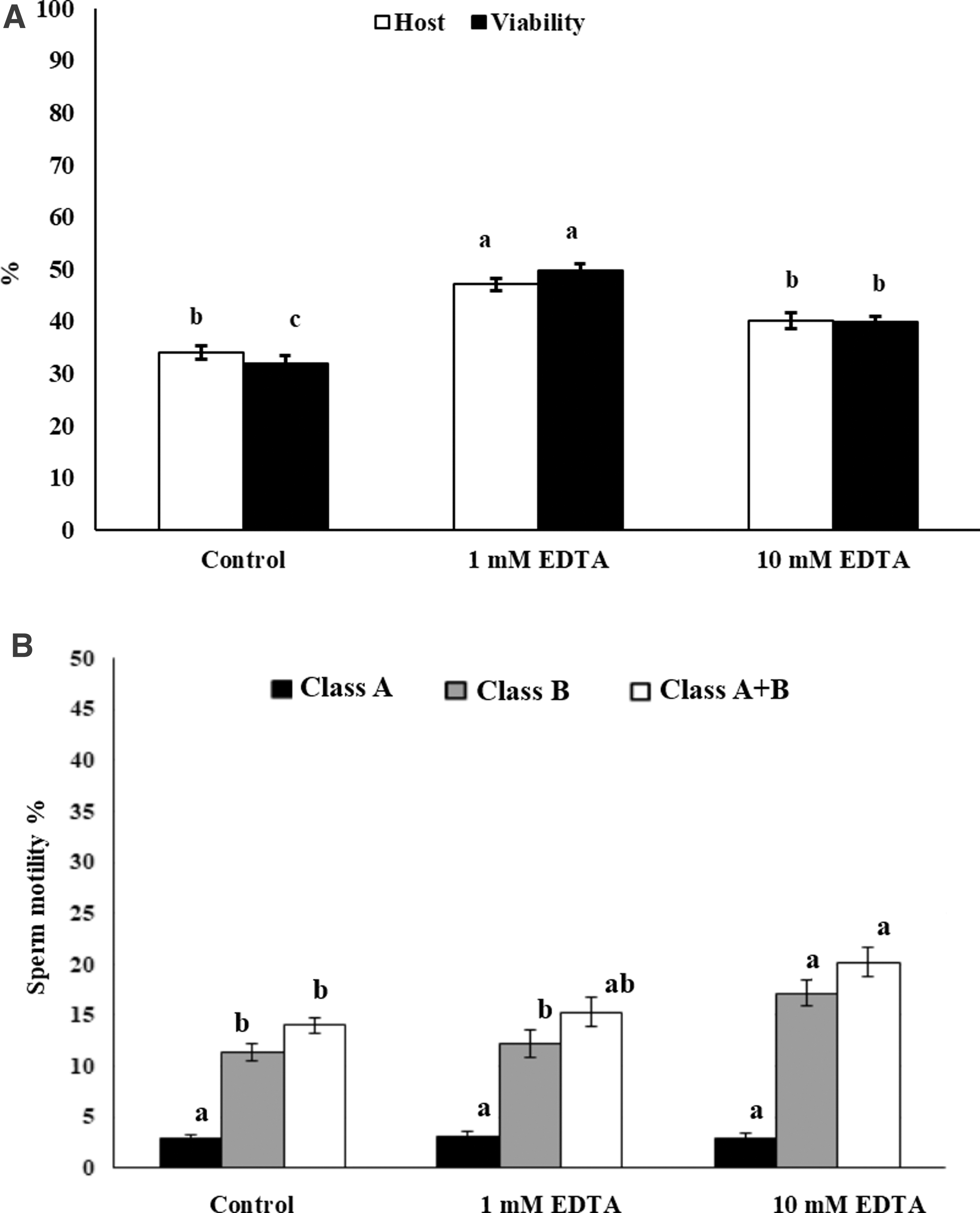
Effect of EDTA on sperm cryopreservation. The goat semen extender was supplemented with 1 and 10 mM EDTA. Plasma membrane integrity and sperm viability were assessed by the HOST and eosin-nigrosin assays
EDTA and estradiol effect on sperm parameters
We co-supplemented 3 μM estradiol, by which the highest rate of progressive motility was observed, with 1 mM EDTA, by which the highest rate of cell viability without affecting the sperm motility was obtained (Figs. 1 and 2). As depicted in Figure 3, results of co-supplementation of estradiol and EDTA showed that sperm membrane integrity and viability were the highest in the estradiol-supplemented group (with or without EDTA) (p < 0.05). The highest rate of sperm motility for class A, B, and A + B belonged to the estradiol group. Combination of EDTA with the estradiol reduced the rate of progressive motility (class A, B, and A+B) compared to the individual estradiol group (p < 0.05). The rate of progressive motility was similar in the combined estradiol + EDTA group compared to the control group (p > 0.05).

Combined effects of estradiol and EDTA on sperm cryopreservation. The goat semen extender was supplemented with 3 μm 17-beta-estradiol and 1 mM EDTA. Plasma membrane integrity and sperm viability were assessed by the HOST and eosin-nigrosin assays
Discussion
In this study, we assessed the effect of supplementing estradiol into the semen extender on the postfreeze parameters of goat spermatozoa. A positive effect of estradiol was observed on the sperm viability and motility. Nonetheless, the higher concentration of estradiol (5 μM) reduced the sperm progressive motility compared to that of 3 μM concentration. It has been shown that estradiol could protect the isolated mitochondria through cryopreservation. 10 However, inclusion of estradiol into the semen extenders has not been assessed on the postfreeze sperm parameters. The mechanism by which estradiol interacts with cells and improves cryoprotection is yet to be explored. The interference of estradiol with cell function is not supposed to be through plasma membrane receptors. 35 It has been repeatedly documented that estradiol exhibits antioxidant activity through the oxidation of lipoproteins of plasma membranes.12–14 Moreover, estradiol was shown to be considerably more effective than α-tocopherol or β-carotene in terms of inhibiting cholesterol oxidation. 13 Another property of estradiol is that it interferes with the intracellular level of calcium and sperm capacitation. 21 We also must consider that over a 2-hour period, the sperm diluents were cooled down in the presence of estradiol. This could provide an effective time for the interaction of estradiol with lipoproteins/cholesterol of the sperm plasma membrane. It has been documented that estradiol supplementation into the sperm diluent media could induce sperm capacitation and acrosome reaction. 29 Apart from the cholesterol pathway, it has been shown that steroids activate the calcium channels in sperm cells.36,37
Removing the seminal plasma might increase sperm membrane damage and permeability.38,39 Cryopreserved spermatozoa had more internal calcium than the corresponding fresh spermatozoa when calcium additives were included in the media. 24 Moreover, a calcium influx takes place in the sperm cytoplasm immediately after thawing, even in extenders with negligible level of calcium or calcium-free media. 26 This very high sensitivity of cryopreserved sperm to the extracellular calcium level resulted in an early induction of sperm capacitation and subsequently compromised the fertilization rate.25,40 Overcoming this issue, supplementation of calcium chelators into the semen extenders has been carried out. Results of this study showed that EDTA supplementation into the semen extender improved the sperm motility and membrane integrity. It has been verified that EDTA increases sperm curvilinear and VSL as well as prolongs the half-life of motile spermatozoa. 41 Reduction of extracellular calcium using EDTA and/or ethylene glycol-bis(beta-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA) improved the sperm motility in the frozen-thawed group compared to the fresh group in various species.26,27 However, in agreement with the results of this study, increasing the concentration of EDTA had an adverse effect on sperm motility and viability.26,42
Then we were interested to see if estradiol's supportive effect on sperm motility is mediated through the intracellular concentration of calcium ionophore. Achieving this aim, we evaluated the estradiol effect in the presence of EDTA, as a calcium scavenger. We detected a considerable interaction between estradiol and EDTA for the sperm motility attributes. Individual EDTA treatment improved the progressive sperm motility compared to the control group, whereas EDTA treatment of the estradiol-supplemented group reduced the progressive motility compared to the individual estradiol group. Use of EDTA into the estradiol-supplemented media reduced the supportive effects of estradiol on the sperm cryopreservation parameters. The results of this study showed that induction of higher sperm motility in response to estradiol is a calcium-dependent process, as the EDTA did completely abrogate the estradiol-mediated effect. The combination of trehalose and EDTA into the ram semen extender increased the cryopreserving activity, not only for freeze-thawing but also for post-thaw resistance, possibly by removing calcium from the medium, thereby preventing cation competition with trehalose for membrane-binding sites. 43
We need to consider that the concentration of estradiol in blood serum of goats and other mammalian species increases during the follicular phase of estrus and reaches its peak at the time of ovulation, followed by the surge of luteinizing hormone (LH) and follicular stimulating hormone (FSH) concentration.44,45 This peak time of estrogen corresponds to the estrus phase, when mating and fertilization take place. Unlike the results of a higher concentration of estradiol in this study, lower concentrations of progesterone, which have a negative correlation with the estradiol concentration during the estrus cycle, have been identified to be effective on sperm hyperactivation and acrosome reaction.20,22 Therefore, the high association of estradiol and sperm motility in this study is in good agreement with the in vivo conditions of female goats at the time of estrus. However, we have to consider that Bioxcell is a soy-based extender, 46 which includes phytoestrogen. There are contradictory reports on the interaction of soy extracts and estradiol.47,48 To address possible synergistic effects of soy extract and estradiol, further experiments with egg-yolk extenders are required.
In conclusion, results of this study showed that estradiol inclusion into the semen extender improved the cryopreservation parameters of goat spermatozoa. Since the increase of progressive sperm motility in response to estradiol was completely abrogated by using EDTA, the estradiol-mediated effects are high likely a calcium-dependent pathway.
Footnotes
Authors' Contributions
Conceived the idea: M.F. Designed the experiment: M.F. and S.E. Carried out the experiments: M.M., M.F., and S.E. Analyzed the data: S.E. Wrote the article: M.F. and S.E. All authors read and confirmed the article content.
Acknowledgments
The authors thank Royan Institute and Transgenesis Center of Excellence, Isfahan Islamic Azad University, for supporting the work.
Author Confirmation Statement
Mr. Mesbah, Dr. Fororouzanfar, and Dr. Eghbalsaied are from Islamic Azad University (Shiraz, Marvdasht, and Isfahan branches, Iran), where education and research are the primary functions.
Ethics Approval and Consent to Participate
The project did not contain clinical studies or patient data.
Availability of Data and Materials
The datasets used and/or analyzed during this study are available from the corresponding author.
Author Disclosure Statement
The authors have no conflict of interests to declare.
Funding Information
This project was not supported by any national/international funding.