
Abstract
Cryopreservation of male germline stem cells (GSCs) is an essential technique for their long-term preservation and utilization in various fields. However, the specific apoptosis pathways involved in cryoinjury during freezing remain unclear. Therefore, our study sought to identify the pathways involved in cryoinjury-induced apoptosis and thereby to improve freezing efficiency during GSC cryopreservation through the creation of a specific molecular-based cryoprotectant. The activities of caspase-8, caspase-9, caspase-3, and caspase-7 were assessed by Western blot analyses to determine the role of specific apoptosis pathways in GSC cryoinjury. Specifically, the role of a specific caspase was identified by recovery rate, relative proliferation rate, Annexin V/propidium iodide co-staining, and caspase activity using its inhibitor and activator. Moreover, the safety of the cryoprotectant was assessed by immunofluorescence and quantitative real-time polymerase chain reaction (qRT-PCR). Furthermore, the efficacy of the molecular-based cryoprotectant was assessed using frozen cells in the presence of dimethyl sulfoxide (DMSO) (control), trehalose, a caspase-8 inhibitor Z-IETD-FMK [ZIF], or a mixture of the aforementioned compounds, after which the changes in Src signaling were measured. Our results demonstrated that caspase-8 plays a major role in cryoinjury-induced apoptosis and therefore its inhibition improves freezing efficiency. Specifically, a significantly higher relative proliferation rate was observed in the Z-IETD-FMK 0.01 μM-treated cells than in the DMSO control (100% ± 6.2% vs. 189.8% ± 9.5%), with decreases in both early apoptosis (16.6% ± 2.2% vs. 7.5% ± 1.0%) and caspase-8 activity (1.0-fold vs. 0.4-fold). The relative proliferation rate was significantly higher in the cryoprotectant mixture (246.0% ± 12.2%) than other individual treatment groups (trehalose 200 mM, 189.8% ± 9.5%; Z-IETD-FMK 0.01 μM, 189.7% ± 2.2%) with no significant differences in Src signaling. Therefore, our findings provide novel insights into the development of freezing protocols to enhance GSC freezing efficiency, thereby facilitating the wider adoption of GSCs in the livestock industry and/or clinical trials.
Introduction
Male germline stem cells (GSCs) have been considered a potential source for male infertility treatment and male germline livestock preservation.1,2 The cryopreservation of these GSCs is essential for their long-term preservation and utilization in various fields. However, frozen cells are inevitably submitted to severe stress conditions during cryopreservation, including cold shock, cellular membrane instability, osmotic stress, caspase activation, and oxidative stress.3,4 Importantly, cryoinjury triggers apoptotic cell death, which reduces freezing efficiency after cryo-thawing.
A cryoprotectant is routinely added to the freezing media to protect the frozen cells from cryoinjury and enhance freezing efficiency.5–7 However, despite the development of various cryoprotectants such as glycerol, dimethyl sulfoxide (DMSO), or their combination with sugar molecules, these existing cryoprotectants mechanically do not protect frozen cells from cryoinjury-induced cell death due to apoptotic stress during freezing.8–10 Therefore, these limitations highlight the need for molecular-based cryoprotectants to alleviate apoptosis during cell freezing and enhance freezing efficiency. 3
Z-VAD-FMK, an apoptosis inhibitor, has been verified to serve as an effective cryoprotectant additive to suppress cryoinjury-induced apoptosis.11,12 In our previous study, Z-VAD-FMK was also found to enhance the proliferation capacity of spermatogonial stem cells after cryo-thawing. 13 Nevertheless, the specific apoptosis pathway (e.g., extrinsic, intrinsic, and both apoptosis) responsible for cryoinjury during freezing has not been identified, which hinders the development of a specific molecular-based cryoprotectant.10,14,15 Therefore, a better understanding of the pathways involved in cryoinjury-induced apoptosis would facilitate the development of more effective and specific molecular-based cryoprotectants.
Our study sought to identify the main cryoinjury-induced apoptosis pathway in GSCs, which would be targeted by a specific molecular-based cryoprotectant, thereby improving GSC freezing efficiency by the addition of a specific inhibitor to the freezing media. Therefore, our study contributes to the development of freezing protocols to enhance GSC freezing efficiency, which may facilitate the wider adoption of GSC in the livestock industry and/or clinical trials.
Materials and Methods
Experimental animals
All animal experiments were approved by the Animal Care and Use Committee of Chung-Ang University (Permit No.: 201900048) and were conducted according to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health. The animal room was maintained at a constant temperature of 23°C ± 1°C and humidity of 55% ± 10% under a 12-hour light/12-hour dark cycle. The mice used in this study were transgenic C57BL/6-TG-EGFP (C57-GFP; Jackson Laboratory, Bar Harbor, ME) in which chicken beta-actin promoter drives enhanced GFP (EGFP) expression in all tissues, except for the erythrocytes and hair.
Isolation and culture of GSCs
All reagents were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise stated. GSCs were isolated from 6- to 8-day-old C57-GFP mouse pup testes following a previously described method with some modifications. 16 The testicular tunica albuginea was first decapsulated, after which the seminiferous tubules were washed in Dulbecco's phosphate-buffered saline (DPBS; Life Technologies, Grand Island, NY). To obtain single cells, the seminiferous tubules were treated with a 2:1 solution of 0.25% trypsin-EDTA (Invitrogen, Carlsbad, CA) and 7 mg/mL DNase I (Roche, Basel, Switzerland) in DPBS (v/v) at 37°C for 5 minutes. The active enzyme was inhibited by adding 10% fetal bovine serum (FBS; Biowest, Nuaillé, France). After filtration through a 40-μm pore nylon mesh (BD Biosciences, San Jose, CA), the cell suspension was centrifuged at 600 g and 4°C for 7 minutes. Erythrocytes and cell suspension debris were eliminated using a 30% Percoll gradient. Single cells resuspended in 1% FBS in DPBS (v/v) were incubated with anti-Thy-1 antibody microbeads (1:10; Miltenyi Biotech, Auburn, CA) for 15 minutes at 4°C and then enriched through magnetic-activated cell sorting, as previously described. 17 Thy-1+ GSCs were plated onto mitotically inactivated SIM mouse embryo-derived thioguanine- and ouabain-resistant (STO) feeder cells and cultured in a mouse serum-free medium (mSFM) consisting of 10 ng/mL glial cell line-derived neurotrophic factor (GDNF; R&D Systems, Minneapolis, MN), 75 ng/mL GDNF family receptor alpha 1 (GFRα1; R&D Systems), and 1 ng/mL basic fibroblast growth factor 2 (bFGF2; BD Biosciences). 18 In this study, GSCs were passaged once weekly, and 8–18 passages were used.
Cryopreservation
All cryoprotectants assessed herein were prepared before freezing. The optimal concentration of each activator and inhibitor was chosen by comparing the negative control (10% DMSO and 10% FBS in DPBS, a basal cryoprotectant) to the positive control (trehalose 200 mM, 10% DMSO, and 10% FBS in DPBS, a standard cryoprotectant), as described in a previous study. 19 Z-IETD-FMK (ZIF; R&D Systems) and gambogic acid (GBA; Tocris Bioscience, Minneapolis, MN) were used as a caspase-8 activator and inhibitor, respectively. All reagents were dissolved in DMSO to prepare stock solutions, which were then diluted 1:1000 with DPBS before use. GSCs were first suspended in each reagent and immediately diluted with an equal volume of cryoprotectant consisting of DMSO and FBS in a dropwise manner. Cell suspensions were transferred to 1.8-mL cryovials (Corning, Midland, MI), placed in a Nalgene® freezing container (Nalgene, Rochester, NY) containing isopropyl alcohol, and stored overnight in a −80°C freezer. The vials were then transferred and stored in liquid nitrogen for at least 1 month before thawing.
Cryoprotectant cytotoxicity assay using the Quanti-MAX™ WST-8 cell viability assay kit
To assess the cytotoxicity of each cryoprotectant, the WST-8 assay was performed using the Quanti-MAX WST-8 assay kit (Biomax, Seoul, Korea) following the manufacturer's instructions with some modifications. GSCs (0.5 × 105) were plated into a 96-well plate and incubated in mSFM containing growth factors for 12 hours in a 5% CO2 incubator at 37°C. The medium was then discarded and the GSCs were further incubated in cryoprotectant for 20 minutes at room temperature (RT; 20°C–25°C) and washed with DPBS. After incubation with the Quanti-Max reagent in a 5% CO2 incubator at 37°C for 2 hours, optical density (OD) values were quantified using a microplate reader (SpectraMax 190; Molecular Devices, Sunnyvale, CA) at a 450 nm wavelength. All data were reported as the mean of triplicate OD values collected using the SoftMax® Pro 5 software (Molecular Devices).
Recovery rate and relative proliferation rate
One month after cryopreservation, the cryovials containing the frozen GSCs were thawed in a 37°C water bath for 2.5 minutes. Then, each cell suspension was diluted 1:10 in 10% FBS (v/v) in minimum essential medium alpha (MEMα) in a dropwise manner. After centrifugation at 600 g and 4°C for 7 minutes, each cell pellet was resuspended in mSFM to calculate the recovery rate (%). The recovery rate (%) was calculated using trypan blue exclusion as follows:
Recovery rate (%) = number of viable cells after freezing, thawing, and washing × 100/number of initial frozen GSCs
After a 1-week culture period, the GSCs were dissociated with 0.25% trypsin and manually pipetted. Following centrifugation at 600 g for 7 minutes at 4°C, the pellet was resuspended in mSFM, and the relative proliferation rate (%) was calculated as follows:
Proliferation rate (%) = number of cells recovered after freezing, thawing, and culture × 100/number of initial cells plated after freezing and thawing
Relative proliferation rate (%) = proliferation rate of treatment groups × 100/proliferation rate of control groups
The control group consisted of frozen GSCs with 10% DMSO (v/v) and 10% FBS (v/v) in DPBS without small molecules.
Western blotting
After thawing, GSCs were cultured for 12 hours in a 5% CO2 incubator at 37°C, as previous studies have reported that the highest rates of cryoinjury occur ∼12 hours after thawing. 20 Proteins were extracted using RIPA buffer (Thermo Fisher Scientific, Rockford, IL) containing protease and phosphatase cocktails (Thermo Fisher Scientific) at 10 × 105 cells/mL and incubated for 30 minutes at 4°C. The supernatants were then collected after centrifugation of the lysate at 13,000 rpm for 20 minutes at 4°C. Protein was quantified using a BCA protein assay (Thermo Fisher Scientific), and 5 μg of each protein was loaded onto a 12% SDS-polyacrylamide gel. The separated protein was then blotted onto a polyvinylidene difluoride (PVDF) membrane (Millipore, Billerica, MA). The membrane was blocked with 0.3% electrochemiluminescence (ECL) in DPBS containing 0.2% Tween 20 (PBS-T) at RT (20°C −25°C) for 1 hour. After washing in PBS-T, the membrane was incubated with primary antibodies diluted 1:1000 at 4°C overnight (rabbit anti-caspase-8 [4790S; Cell Signaling Technology (CST), Danvers, MA, USA], rabbit anti-caspase-9 [9504S; CST], rabbit anti-caspase-3 [14220S; CST], rabbit anti-caspase-7 [9492S; CST], and rabbit anti-Src [2108; CST]). Mouse anti-α-tubulin (ab7291; Abcam, Cambridge, United Kingdom) was used as a loading control at a 1:5000 dilution. After washing with PBS-T, the membrane was incubated with HRP-conjugated secondary antibody diluted 1:2000 at RT (20°C–25°C) for 2 hours; anti-rabbit IgG (7074S; CST) and anti-mouse IgG (7076S; CST) were also used. Protein expression was determined using the ECL method, and the band intensity was evaluated using the ImageJ software (version 1.8.0; National Institutes of Health). In our study, caspase activity was measured by the ratio of active to inactive caspase intensity.
Apoptosis assay
GSCs were harvested at 12 hours after thawing and culturing in a 5% CO2 incubator at 37°C. After washing in cold DPBS, the pellet was resuspended in 1 × binding buffer (BD Biosciences), and then the cell suspension (1 × 105 cells) was transferred to a new tube. The cell suspension was incubated in annexin V-APC (BD Biosciences) and propidium iodide (PI) for 15 minutes at RT (20°C −25°C) in the dark. Apoptotic cells were assessed using flow cytometry with a FACSAria II cell sorter (BD Biosciences) equipped with BD CellQuest™ Pro software (Becton Dickinson, Oxford, United Kingdom).
Immunofluorescence
After a week of in vitro culture following cryopreservation and thawing, the GSCs were harvested and fixed with 4% paraformaldehyde (Biosesang, Seongnam, Korea) at RT (20°C–25°C) for 30 minutes and permeabilized with 0.1% Triton X-100 (v/v) in DPBS at RT (20°C–25°C) for 10 minutes. The cells were then washed with DPBS and blocked with 5% (w/v) bovine serum albumin for 1 hour, followed by overnight incubation with primary antibodies diluted 1:200 at 4°C overnight (rabbit anti-promyelocytic leukemia zinc finger [PLZF, NBP1–80894; Novus Biologicals, Centennial, CO], rabbit anti-glial-derived neurotrophic factor family receptor alpha 1 [GFRα1, ab8026; Abcam], rabbit anti-DEAD-box polypeptide 4 [DDX4, also known as VASA, ab13840; Abcam], and mouse anti-KIT proto-oncogene receptor tyrosine kinase [c-Kit, sc-365504; Santa Cruz Biotechnology]). The cells were washed three times with DPBS and incubated with Alexa Fluor 568-conjugated anti-rabbit IgG (A11011; Invitrogen) or Alexa Fluor 568-conjugated anti-mouse IgG (A11004; Invitrogen) at RT (20°C–25°C) for 1 hour. VectaShield® mounting medium containing 4,6-diamidino-2-phenylindole (DAPI; LSBio, Seattle, WA) was used to mount the cells and stain their nuclei. Marker expression (%) was analyzed using a TS-1000 microscope interfaced with the NIS Elements imaging software (Nikon, Tokyo, Japan). Five microscopic fields were then randomly selected to calculate the number of labeled cells among the GFP+ GSCs.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using the TRIzol reagent (Invitrogen). The PureLink™ RNA Mini Kit (Invitrogen) was then used for RNA purification according to the manufacturer's recommendations. RNA purity and quantity were measured using a NanoDrop spectrophotometer, after which 1500 ng of total RNA was used for complementary DNA (cDNA) synthesis using the SuperScript IV First-Strand Synthesis System (Invitrogen) and oligo-(dT) primers. The cDNA was diluted to 300 ng/μL using RNA-free water, and 1 μL of the dilution was used in quantitative real-time polymerase chain reaction (qRT-PCR) comprising 5 μL of 2 × SYBR Green PCR Master Mix and 1 μL (10 pmol/μL) of each primer. [LIM homeobox 1 (Lhx1), DAZ-like (Dazl), and synaptonemal complex protein 1 (Sycp1); glyceraldehyde-3-phosphate dehydrogenase (Gapdh); Table 1]. Gapdh was used as the internal control. The qRT-PCR assays were conducted on a 7500 Real-Time PCR System (Applied Biosystems, Carlsbad, CA) with the following conditions: initial denaturation at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute in a two-step thermal cycle, and a final melting curve step at 95°C for 15 seconds, 60°C for 1 minutes, 95°C for 30 seconds, and 60°C for 15 seconds. All quantification cycle (CT) values were normalized to the Gapdh levels and quantification was performed using the 2−ΔΔCT method.
Quantitative Real-Time Polymerase Chain Reaction Primers
Statistical analyses
All statistical analyses were conducted with SPSS version 20 (IBM, Armonk, NY). The Shapiro–Wilk and Levene's tests were used to assess normal distribution and homogeneity of variance, respectively, and multiple comparisons between samples were conducted using one-way analysis of variance (ANOVA) coupled with the post hoc Tukey's honestly significant difference test. Dunnett's test was used to determine whether each of the treatments has a significant difference than the control for screening of dose-dependent effect. All experiments were conducted at least in triplicate unless otherwise stated, and all data were reported as the mean ± SEM. A significance level of p < 0.05 was deemed statistically significant.
Results
Cryoprotective efficiency decreased due to extrinsic apoptosis after freezing
Proliferation rate was used as an indicator to assess the freezing efficiency on GSCs after cryo-thawing. The proliferation rate of frozen GSCs was significantly lower compared with unfrozen GSCs, 4 days (160.7% ± 22.3% vs. 113.3% ± 12.8%; fresh vs. cryopreserved) and 7 days (285.2% ± 53.8% vs. 174.1% ± 35.9%) after thawing (Fig. 1A). To identify a major cryoinjury-induced apoptosis pathway, the activity of different caspases was assessed using representative markers for extrinsic apoptosis (caspase-8), intrinsic apoptosis (caspase-9), and executioner caspase (caspase-3 and caspase-7). In our study, caspase activity was measured by the ratio of cleaved- to pro-form (Supplementary Table S1). The activity of caspase-8 (1.0 ± 0.3-fold vs. 28.0 ± 11.8-fold), caspase-3 (1.0 ± 0.1-fold vs. 4.1 ± 0.3-fold), and caspase-7 (1.0 ± 0.1-fold vs. 8.5 ± 2.3-fold) was significantly higher in frozen GSCs compared to that of unfrozen GSCs, except for caspase-9 activity (1.0 ± 0.1-fold vs. 0.9 ± 0.0-fold; Fig. 1B). These findings indicate that cryoinjury is mainly associated with extrinsic apoptosis rather than intrinsic apoptosis in germ cells.
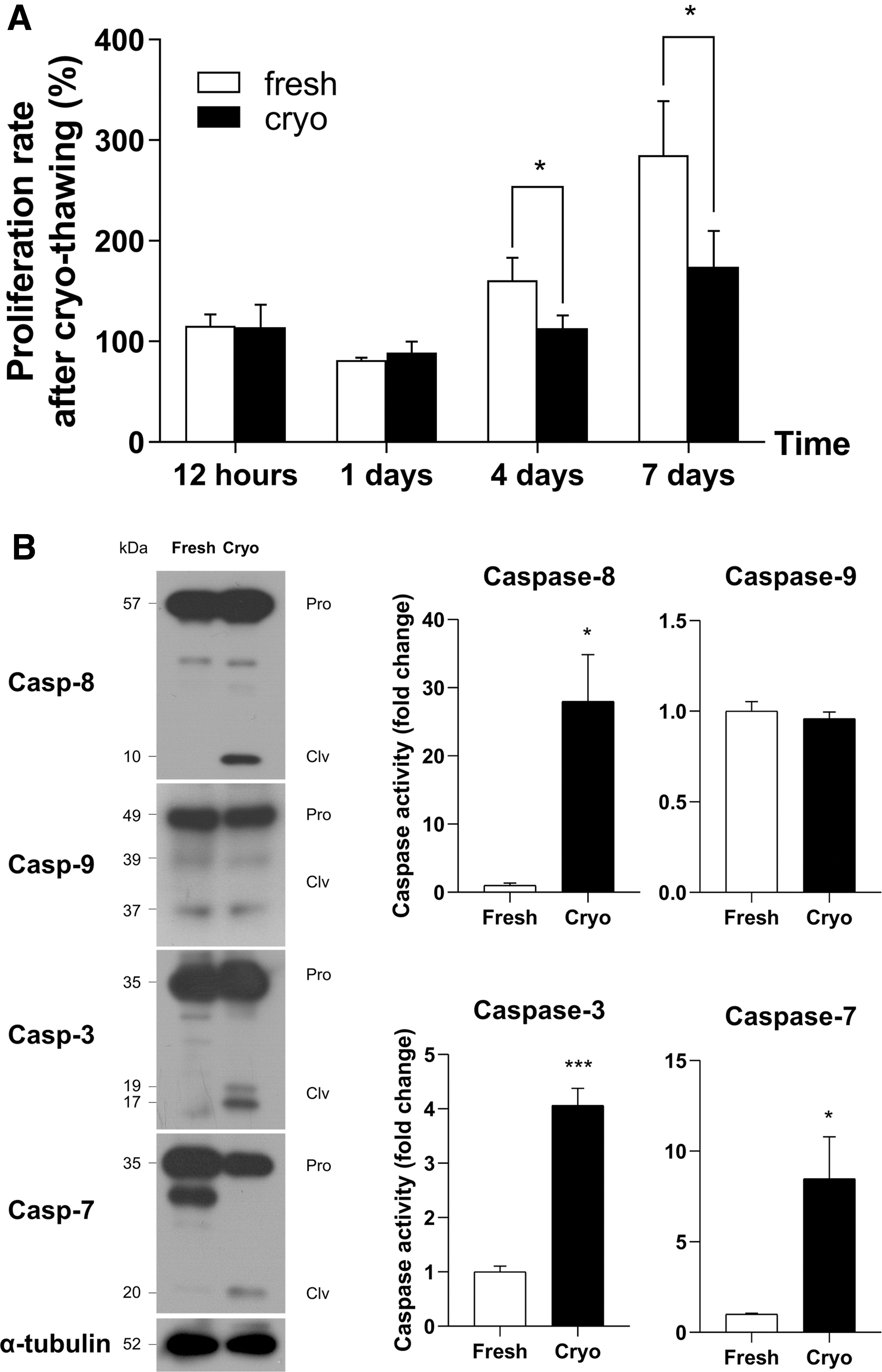
Cryoinjury-induced apoptosis in frozen GSCs.
Inhibition of caspase-8 activity improves GSC cryoprotective efficiency
Although cryoprotectants are necessary to protect frozen cells from ice crystallization during freezing, these benefits are often inevitably coupled with several negative side effects (e.g., cytotoxicity). Therefore, a WST-8-based cytotoxicity assay was conducted to confirm that cryoinjury was not caused by the cryoprotectant itself. There were no significant differences in cytotoxicity between any of the treatment groups and the negative controls (Fig. 2A). Therefore, the potential negative impacts of the cryoprotectant itself could be ruled out in all subsequent experiments. Based on the data discussed above (Fig. 1), we assessed freezing efficiency using different concentrations of a caspase-8 inhibitor (ZIF) and activator (GBA) to determine the role of extrinsic apoptosis in cryoinjury. No significant difference in recovery rate after cryo-thawing was observed between the control and treatment groups (Fig. 2B). However, 0.01 and 0.1 μM ZIF treatment rendered a significantly higher relative proliferation rate (189.8% ± 9.5% and 159.8% ± 14.5%, respectively) after cryo-thawing and culture compared to the DMSO control (100% ± 6.2%), whereas a significantly lower relative proliferation rate was observed with ZIF 100 μM and GBA 1 μM (52.7% ± 7.7% and 61.9% ± 5.7%, respectively; Fig. 2C). Furthermore, normal-shaped GSC clumps were observed after freezing with ZIF 0.01 μM (Fig. 2D). Therefore, the ZIF 0.01 μM and GBA 1 μM treatments were used in all downstream experiments to investigate whether extrinsic apoptosis modulation has beneficial effects on germ cell freezing efficiency.
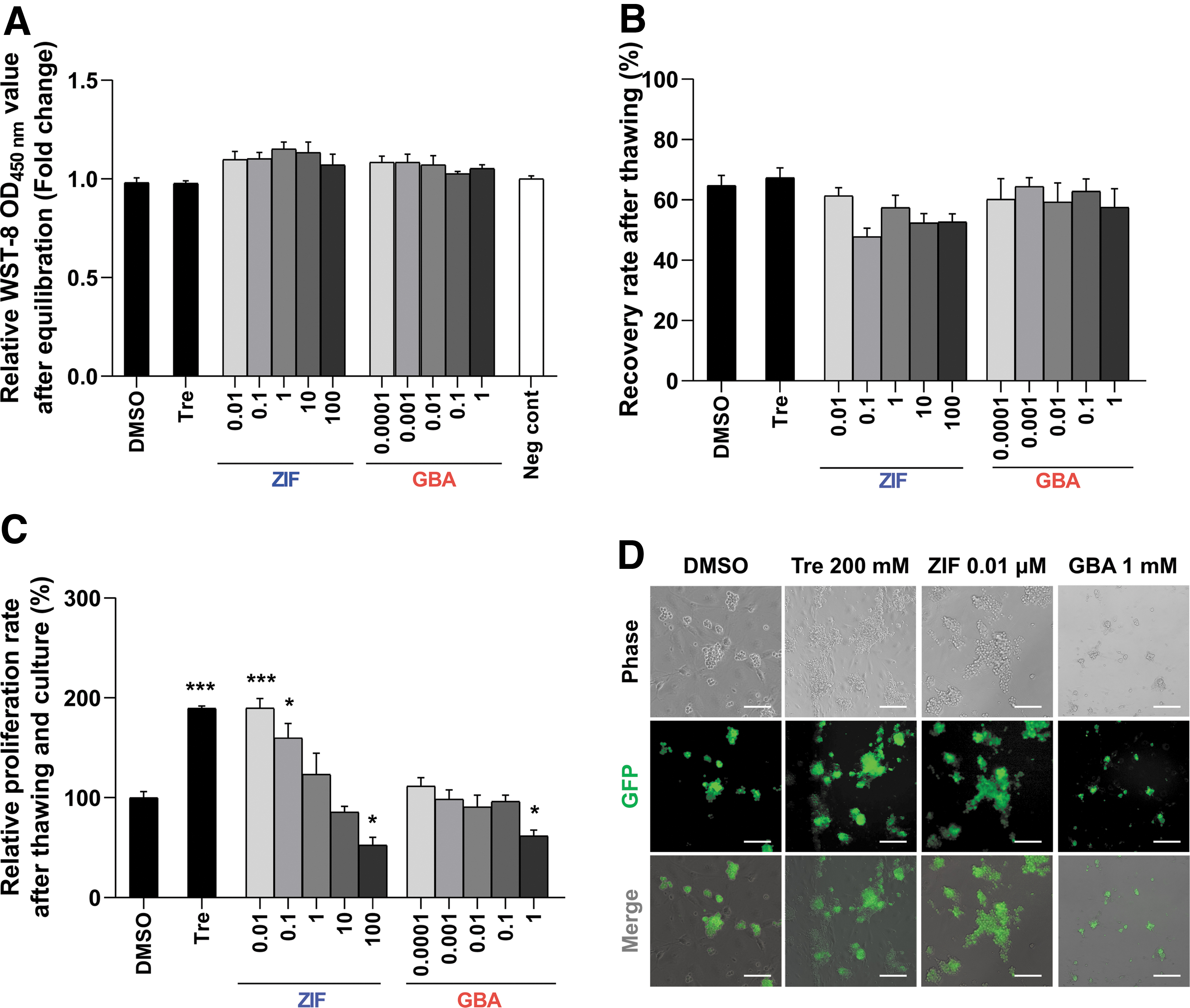
Effect of a dose-dependent caspase-8 inhibitor (ZIF) or activator (GBA) on frozen GSCs.
Z-IETD-FMK reduces apoptosis in frozen GSCs
Apoptosis level (annexin V-APC/PI co-staining) and caspase activity were assessed to verify the effects of extrinsic apoptosis modulation in frozen GSCs. Early apoptosis was significantly alleviated in the trehalose 200 mM and ZIF 0.01 μM treatment groups compared to the DMSO control (16.6% ± 2.2%, 7.2% ± 0.9%, and 7.5% ± 1.0%; DMSO control, trehalose 200 mM, and ZIF 0.01 μM, respectively; Fig. 3A). In addition, caspase activity of each control and treatment group was measured by the ratio of cleaved- to pro-form (Supplementary Table S2). Our Western blot analyses indicated that caspase-8 activity was significantly downregulated in the frozen GSCs with ZIF 0.01 μM compared to the DMSO control, whereas no significant differences were observed in the trehalose 200 mM group (1.0 ± 0.0-fold, 0.2 ± 0.0-fold, and 0.4 ± 0.0-fold; Fig. 3B). Likewise, caspase-3 and caspase-7 activities were also significantly decreased by ZIF 0.01 μM than with DMSO control, which was similar to the effect of trehalose 200 mM (caspase-3, 1.0 ± 0.1-fold, 0.5 ± 0.1-fold, and 0.5 ± 0.1-fold and caspase-7, 1.0 ± 0.1-fold, 0.6 ± 0.1-fold, and 0.3 ± 0.1-fold). In contrast, caspase-8 activity was higher in the GBA 1 μM treatment group compared to the DMSO control (1.0 ± 0.1-fold and 2.6 ± 0.1-fold; DMSO control and GBA 1 μM, respectively), whereas no significant difference was observed between caspase-3 and caspase-7 activity compared to the DMSO control. Therefore, our results indicated that the ZIF 0.01 μM treatment effectively suppressed cryoinjury in GSCs.
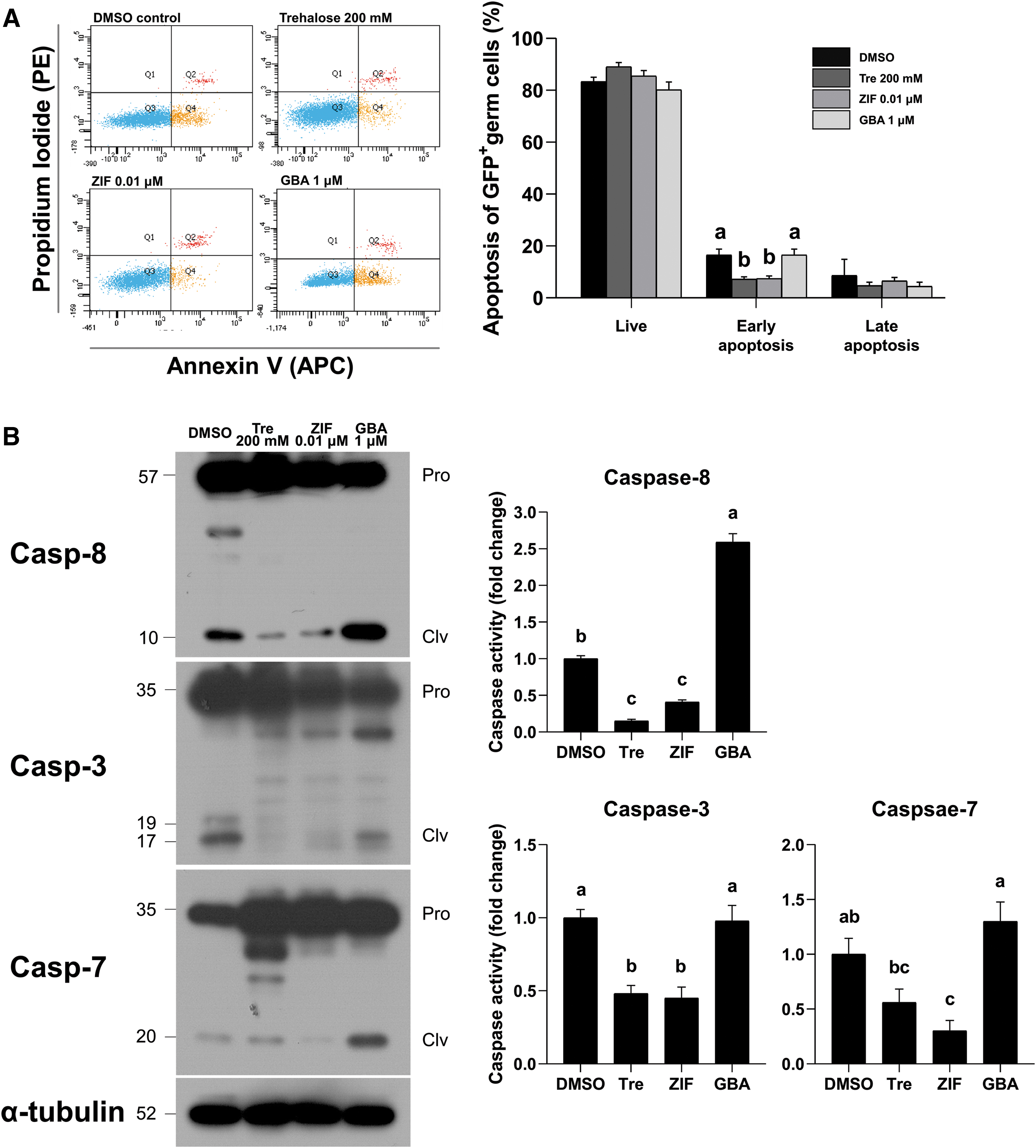
Cryoprotective efficiency of 0.01 μM caspase-8 inhibitor (ZIF) in GSCs.
GSCs remained stable after freezing with Z-IETD-FMK
Unlike other cells, GSCs pass down their genetic information to subsequent generations. Therefore, assessing the safety of cryoprotectant additives is especially crucial for germ cell freezing purposes. Therefore, the stability of germ cells that had been previously cryopreserved with ZIF 0.01 μM was evaluated by immunofluorescence and qRT-PCR. According to our immunofluorescence assays, the expression of GFRα1 and PLZF (undifferentiated spermatogonia markers), as well as VASA (a germ cell lineage marker), fell within a normal range, with no significant difference between any of the tested groups (Fig. 4A). Similarly, no difference in the expression of c-Kit (a differentiating spermatogonia marker) was observed between any of the tested groups. Also, no difference in the expression of Lhx1 (undifferentiated spermatogonia markers), as well as Dazl and Sycp1 (differentiated spermatogonia markers), was observed between groups, as demonstrated by qRT-PCR analyses (Fig. 4B). Our findings indicate that the normal properties of GSCs were retained after cryo-thawing regardless of the type of cryoprotectants used, suggesting that the proposed cryoprotectant treatment not only enhances cell yields after cryo-thawing but also preserves the genetic integrity of the GSCs.
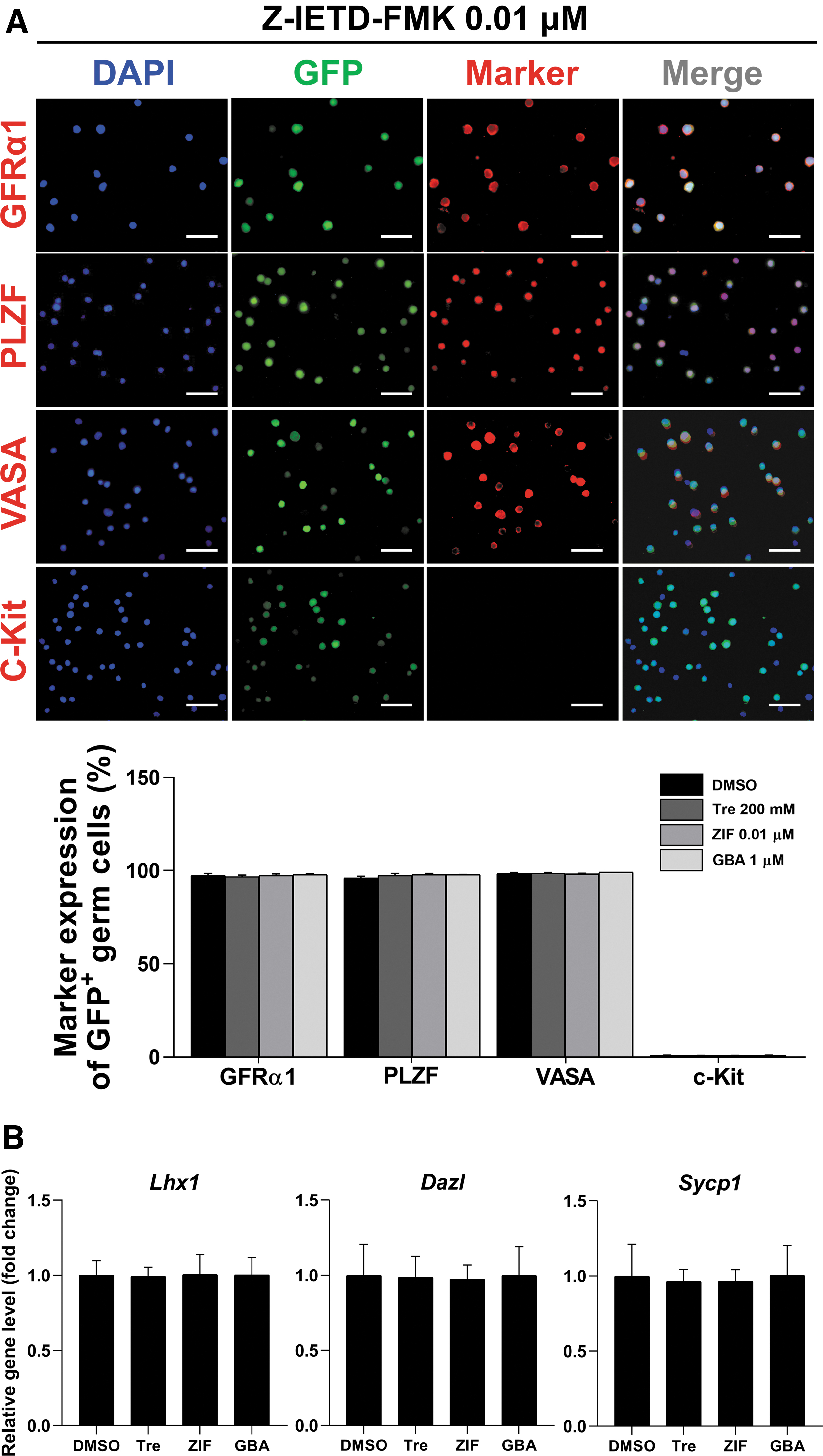
Characterization of GSCs after freezing with 0.01 μM caspase-8 inhibitor (ZIF).
Trehalose and Z-TETD-FMK synergistically enhance cryo-thawed GSC proliferation
To assess whether trehalose and ZIF could provide a synergistic cryoprotective effect, the effect of a trehalose 200 mM and ZIF 0.01 μM mixture on recovery rate, relative proliferation rate, and GSC self-renewal regulation-related signaling was evaluated. Although recovery rate was not significantly affected by this mixture, the relative proliferation rate was significantly higher in the combined treatment group compared to each individual treatment group (189.8% ± 9.5%, 189.7% ± 2.2%, and 246.0% ± 12.2%; trehalose 200 mM, ZIF 0.01 μM, and trehalose 200 mM + ZIF 0.01 μM, respectively; Fig. 5A). Moreover, normal-shaped GSCs were also exhibited in the combined treatment group (Fig. 5B), and no significant effect on Src expression was observed in any of the tested groups (Fig. 5C), thus confirming that the cryoprotectants did not affect the properties of the GSCs during freezing. These results highlight the capacity of the proposed mixed trehalose and ZIF treatment to render high cell yields after cryo-thawing and culturing.
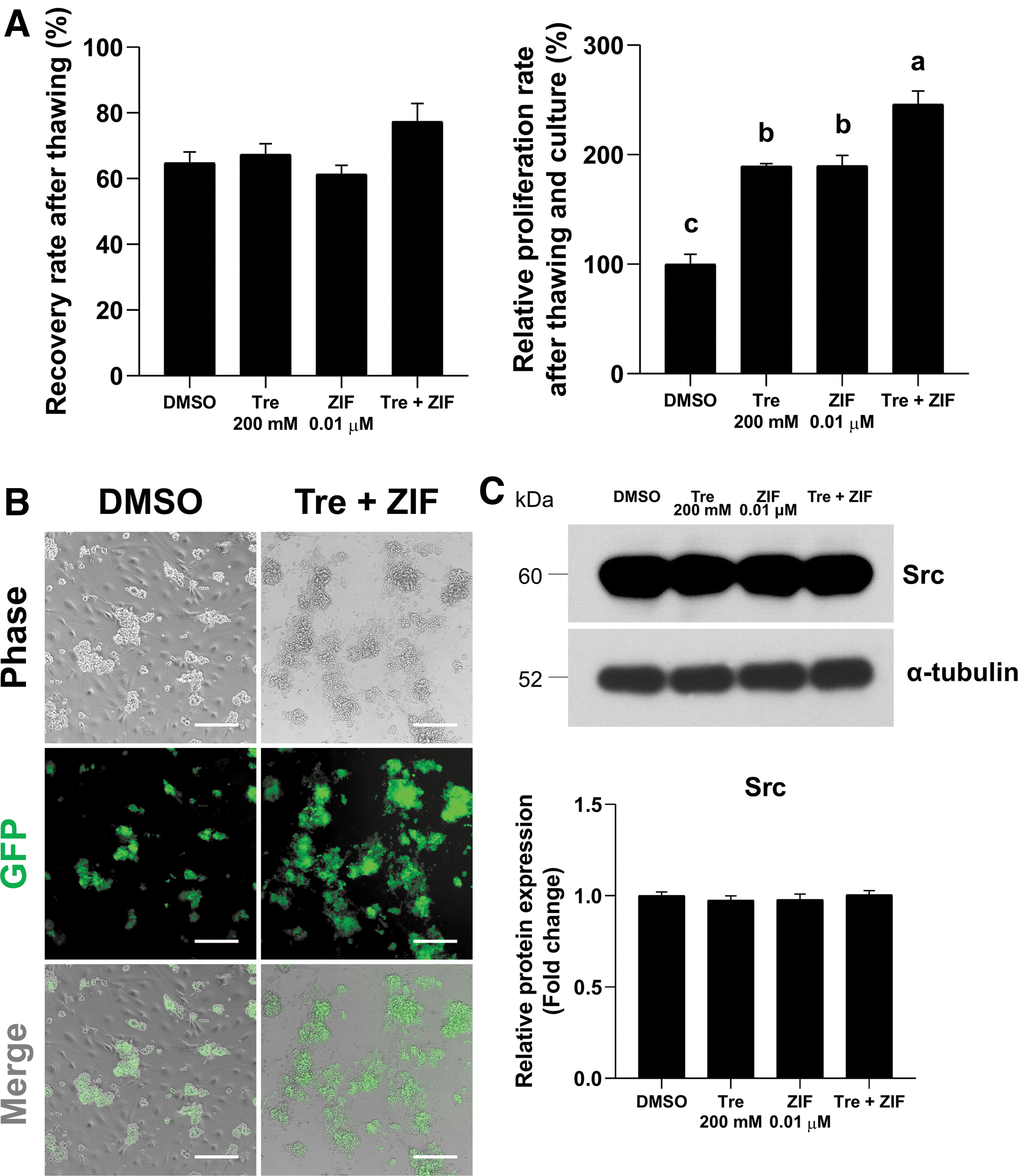
Combined effects of 0.01 μM caspase-8 inhibitor (ZIF) and 200 mM trehalose on GSC freezing efficiency.
Discussion
Cryopreservation is an essential technique to preserve male GSCs in both clinical settings and/or the livestock industry. However, despite recent efforts to improve freezing protocols based on the inhibition of apoptosis, previous strategies only targeted general apoptotic mechanisms of cryoinjury, thus neglecting the role of extrinsic and intrinsic apoptosis. Therefore, a better understanding of cryoinjury-related apoptosis pathways would establish a molecular basis for the development of new strategies to improve GSC cryopreservation efficiency. This study sought to investigate the involvement of caspase-mediated pathways in GSC cryoinjury and the potential of a caspase modulator to serve as a cryoprotectant additive to improve freezing efficiency.
Our results determined that GSCs exhibited a significant decrease in proliferation rate 4 and 7 days after cryo-thawing. This may be attributed to cryoinjury early after cryo-thawing, which is consistent with previous studies that reported cryoinjury occurs within the first 24 hours after cryo-thawing. 21 In particular, our results demonstrated a significant increase in caspase-8 activity in GSCs after cryo-thawing, which is consistent with a previous study that freezing triggers the production of ROS, resulting in the activation of caspase-8 and caspase-3 in frozen granulosa cells. 22 In contrast with our results, some studies have demonstrated that cytochrome c release resulting from freezing increases caspase-9 activity in frozen human colon carcinoma. 23 These inconsistencies may be associated with different cryoprotectant concentrations, which is in accordance with a previous study reporting that apoptotic signaling pathways are dependent on the concentration of the cryoprotectant used during freezing. 24 Therefore, our results suggest that cryoinjury was likely involved in a caspase-8-dependent pathway in frozen GSCs.
To verify the correlation between cryoinjury and the caspase-8-dependent apoptotic pathway, small molecules that inhibit or activate caspase-8 were added to the cryoprotectant at different concentrations. As expected, the modulation of caspase-8 activity affected freezing efficiency. Moreover, the inhibition of caspase-8 activity improved the relative proliferation rate of GSCs after cryo-thawing and vice versa. In addition, a decrease in early apoptosis as well as caspase-8, caspase-3, and caspase-7 activity was observed in frozen GSCs treated with ZIF 0.01 μM according to flow cytometry and Western blot analyses, respectively. These observations are further supported by reports that freezing triggers caspase-8-dependent cryoinjury, which is a factor that decreases freezing efficiency in GSCs.
Interestingly, our data also indicated that excessively high concentrations of the caspase-8 inhibitor considerably reduced its efficiency, thus highlighting the importance of optimization. Previous studies have also reported that the best cryopreservation performance is achieved with an optimal range of a general apoptosis inhibitor in frozen hematopoietic progenitor cells. 25 Furthermore, given that the potential toxicity of the caspase-8 modulator was ruled out by a cytotoxicity assay before freezing in this study, the fact that higher concentrations of inhibitor were less protective might be explained by a positive or negative potential role of apoptosis in cellular homeostasis. 26 Therefore, our results suggest that optimal modulation of caspase-8 activity can render maximal cryoprotective effects.
Quality assurance of frozen GSCs is also necessary to ensure that these cells can be safely applied after cryo-thawing. 27 In this study, GSCs remained normal after freezing with ZIF 0.01 μM, which indicated that the GSCs maintained their ability to self-renew without spontaneous differentiation after freezing. In addition, our results demonstrated that the cells remained stable regardless of the type of cryoprotectant used. This result is consistent with another study that reported stem cells remained stable after cryo-thawing, 28 which suggests that an effective freezing protocol for the preservation of GSCs is conducive to high cell yields after cryo-thawing and culturing.
Taken together, our data indicated that ZIF 0.01 μM can be used as an effective cryoprotectant additive for GSC freezing. Nevertheless, we recommend the development of a protocol for multilateral protection from not only apoptosis inhibition but also other types of cryoinjury. Frozen cells endure a series of cryoinjuries such as cold shock, cellular membrane instability, osmotic stress, caspase activation, and oxidative stress during freezing.3,4 Therefore, to develop more effective GSC cryoprotectants, we examined the combined effects of ZIF and trehalose (200 mM), the latter of which has been previously reported as an effective GSC cryoprotectant. 19 In addition to evaluating freezing efficiency, quality assurance after cryo-thawing was also demonstrated by the normal expression of Src, a crucial self-renewal signal, 16 in frozen GSCs treated with the ZIF and trehalose mixture. Unlike ZIF capable to inhibit apoptosis after cryo-thawing, trehalose physically protects the cellular membrane from ice crystals through their disaccharide-bilayer system. 29 Thus, our study emphasizes that multilateral protection with a cryoprotectant mixture can synergistically enhance the outcomes of GSC freezing without spontaneous differentiation after cryo-thawing.
Conclusion
Our study demonstrated the involvement of an extrinsic apoptosis pathway in the occurrence of major GSC cryoinjuries. Therefore, ZIF (i.e., a caspase-8 inhibitor) can be used as an effective cryoprotectant additive to improve GSC freezing efficiency. However, our study also determined that a mixture of trehalose 200 mM and ZIF 0.01 μM provided additional multilateral protection from cryoinjury. Thus, our findings establish the basis for the creation of molecular mechanism-based freezing protocols, and therefore the use of GSCs could become more widespread in the livestock industry and/or male infertility clinics.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MEST) (NRF-2020R1A2C1009974) and the Chung-Ang University Research Scholarship Grant in 2019.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.