
Abstract
Oxidative stress is a major contributory factor to cellular damage during semen cryopreservation and results in a decreased fertilizing capacity of cryopreserved bull sperm. The inclusion of exogenous antioxidants sometimes exerts deleterious effects on sperm quality. Thus, enhancing the endogenous production of antioxidants is a requirement. This study aimed to investigate the effect of milk type heated at different temperatures on the antioxidant potential of extenders, and the subsequent post-thaw quality parameters and in vivo fertility of buffalo bull semen. Cow (C) and buffalo whole milk (B) were used separately for semen extender preparation, heated at five different temperatures (T1 = 90°C, T2 = 100°C, T3 = 110°C, T4 = 120°C, T5 = 130°C) for 10 minutes. Reactive sulfhydryl groups were measured in each subgroup by Ellman's reagents as CT1 = 143.2 μM, CT2 = 147.4 μM, CT3 = 151.5 μM, CT4 = 157.2 μM, CT5 = 161.8 μM, BT1 = 168.3 μM, BT2 = 172.5 μM, BT3 = 176.7 μM, BT4 = 196.3 μM, and BT5 = 205.7 μM. All semen samples were cryopreserved in milk-based extenders by using standard procedures. Post-thaw quality parameters including total and progressive motility, mitochondrial membrane potential, plasma membrane integrity, and acrosome integrity were found to be higher (p < 0.05) in the group (BT3) containing buffalo milk heated at 110°C, whereas in the same group, lipid peroxidation was found to be lower (p < 0.05) as compared with other treatment groups and control group. In vivo fertility of cryopreserved buffalo sperm was compared among BT3, CT1 (conventionally used milk extender), and a Tris egg yolk extender group. The fertility rates [47% (54/114), 30% (33/108), and 36% (37/103)] were higher (p < 0.05) in BT3 as compared with other groups. This study suggests that buffalo milk heated at 110°C has high antioxidant potential and improves post-thaw quality and in vivo fertility of cryopreserved buffalo bull semen.
Introduction
In recent years, the buffalo population is showing an increasing trend in Southeast Asian and Mediterranean regions, indicating that buffaloes play a pivotal role in strengthening the agricultural economy and increasing food security in these countries. Owing to its prime quality milk, buffalo breeding has also been recorded as a growing area of interest during the last decade. 1 Human health-related beneficial fatty acids, that is, 2-omega-3 FA (C18:3 cis-9, 12, 15 and C20:5n3) and oleic acid, cis-9 trans-11 are high in buffalo milk fat,2,3 which has led to increasing reproductive biotechnologies utilization for the improvement of the genetic pool.
However, harmful effects occur in sperm because of freezing and thawing procedures that result in a decreased motility percentage and low sperm viability, 4 which compromise their survival after insemination within the female reproductive tract. 4 Sperm cells encounter these detrimental effects owing to disruption of cell membrane integrity, 6 osmotic pressure disturbance across the plasma membrane, 7 reactive oxygen species (ROS) production, 8 and cell death pathways. 4 Among the major reasons for low fertility in buffalo under field conditions, decreased sperm survival and poor freezability are considered most important. 4
As compared with cattle, buffalo sperm are more vulnerable to cryo-injuries.9–12 The higher concentration of polyunsaturated fatty acids (PUFAs) in the cell membrane makes it more prone to oxidative damage,13,14 resulting in decreased sperm motility, 15 suppression of oxidative metabolism, 16 DNA damage, 17 sperm plasma membrane damage, 18 reduced oocyte penetration ability,19,20 and impairment of mitochondria. 21 Hence, post-thaw semen quality and the fertility rate are decreased. 22
Antioxidant supplementation of semen extender has valuable effects on cryo-survivability of bovine sperm.23–27 However, these antioxidants have to be incorporated in semen extender exogenously, and sometimes also exert detrimental effects at higher concentrations. There is a lack of scientific reports investigating the process of indigenous production of the antioxidant species in semen extender during its preparation. Hence, it is imperative to explore a favorable and efficient cryopreservation regime with high antioxidant ability for buffalo bull spermatozoa.
Whole milk (WM)-based extender can be used efficiently for the cryopreservation of sperm in the presence of glycerol. Casein micelles, a major protein of milk, seems to be one of the potent protective components of milk.28–30 It also possesses antioxidant activity, which is provided by reactive sulfhydryl (SH) groups. These reactive SH groups are believed to be associated with the whey protein present in milk, 31 and produced owing to the unfolding of milk proteins, and which act as free-radical scavengers in biological systems. Owing to this behavior, SH groups are considered to be effective antioxidants. 32 Moreover, milk antioxidant activity can be potentially increased through heat treatment. 33
Furthermore, the addition of lactose to an extender containing caseins improved the extender efficiency during bull sperm freezing. 28 This is the first study, to the best of our knowledge, designed to determine the effects of antioxidant species production (SH groups) after different thermal treatments of milk on post-thaw quality parameters and in vivo fertility of buffalo bull semen cryopreserved in milk-based extender.
Materials and Methods
Ethical statement
The present study was conducted after the approval of Advance Studies and Research Board, University of Veterinary and Animal Sciences Lahore, Pakistan.
Chemicals
All the chemicals were procured from Sigma-Aldrich Co. (Missouri) unless otherwise mentioned.
Experimental site and animals
The present research was conducted at the Al-Haiwan semen production unit (SPU) Sahiwal. Nili Ravi buffalo bulls selected (n = 4; age = 5–6 years) for the research trial were regular semen donors. The housing, feeding, and management conditions were ensured to be uniform and optimum for each bull.
Milk extender preparation
Cow (C) and buffalo (B) milk were used separately to prepare WM semen extenders. Five different thermal treatments (termed T1 = 90°C, T2 = 100°C, T3 = 110°C, T4 = 120°C, and T5 = 130°C) were used in both cow and buffalo milk for 10 minutes. Using an autoclave, WM was cooled at room temperature and antibiotics were added after filtration by Whatman filter paper (WHA7404004). 34 Two fractions of WM were prepared; Fraction A contained antibiotic-supplemented milk without glycerol and Fraction B comprised antibiotic supplemented with 14% glycerol (v/v).
SH assay
After the heat treatment of milk, Ellman's reagent was used to measure the reactive SH groups in aqueous solution35,36 with a slight modification. A 2 mL milk sample was mixed with 7.9 mL buffer. The pH of the sample was adjusted to 8.2 (by adding tromethane hydrochloride) and 0.1 mL Ellman's reagent solution was mixed. The solutions were centrifuged (Beckman Coulter Floor Model Ultracentrifuge) at 116,000 g and filtered using 0.45 μm filter paper. The absorbance was recorded at 412 and 540 nm using a spectrophotometer (SPECORD 200 plus®). The absorbance of samples without DTNB was recorded against water to correct for turbidity and a standard curve was constructed.
Semen collection, processing, and cryopreservation
Semen was collected twice a week from each bull by using an artificial vagina. All samples (bulls = 4, replicates/bull = 6, total samples 6 × 4 = 24), after collection, were shifted to a water bath at 37°C for initial evaluation. Only those semen samples having a minimum of 65% motility and 500 million sperm count were selected for the study.37,38
Every ejaculate from each bull was divided and diluted in milk-based extenders (groups mentioned previously). First, individual ejaculates were added to fraction A of various heat-treated milk samples at 37°C and gradually cooled down to 4°C. Fraction B, having 7% (v/v) glycerol, was added in equal volume to make a final concentration of sperm of 20 × 106 cells/mL. 23 About 0.5 mL straws were used for the collection of semen. After the glycerol addition, semen straws were sealed and frozen within 4 hours. After keeping in liquid nitrogen (LN2) vapor for 10 minutes, these straws were plunged directly into LN2. 39
Post-thaw sperm structural and functional evaluation
Post-thaw sperm quality analysis was conducted after 24 hours of cryopreservation40,41 at the Theriogenology Laboratory, University of Veterinary and Animal Sciences, Lahore, Pakistan. Three straws/replicate of each group were evaluated for quality parameters. 38
Computer-assisted semen analysis
Sperm total and progressive motilities, velocity distribution, and kinematics were evaluated using computer-assisted semen analysis (CASA) (number of frames: 30, frame rate: 60 Hz/sec, minimum cell contrast; 15, minimum cell size [pixel] 8, cell intensity, threshold straightness 80, medium VAP 25 μm/s, low-VAP cutoff 5.0, VSL 0.05 μm/s). 42 A drop of 7 μL thawed semen sample was placed onto on a prewarmed slide. The observation was made at 200 × . Post-thaw parameters including total motility (TM %), progressive motility (PM %), average path velocity (VAPμm/s), curvilinear velocity (VCLμm/s), straight-line velocity (VSLμm/s), straightness (STR%), and linearity (LIN %) were recorded. For each motility parameter, 200 sperm were assessed during CASA.
Sperm viability percentage
A propidium iodide assay was performed for the assessment of sperm viability. 43 In brief, 50 μL of thawed semen sample was diluted with 50 μL of Tris critic acid fructose buffer (TCF) (Tris 2.42 g, citric acid 1.34 g, fructose 1 g, 100 mL distilled water) and centrifuged at 300 g for 5 minutes to make a sperm pellet. To make a final volume of 47.5 μL, the sperm pellet was resuspended in Tris critic acid fructose buffer. About 2.5 μL propidium iodide (10 μg/mL in distilled water) was added in the solution.
The sperm–PI mixture was incubated at 37°C for 5 minutes in the dark. A drop of this mixture was placed on a glass slide and observed under the microscope (Olympus BX53) at 100 × and a minimum of 200 sperm were counted. Sperm with red fluorescence at the head region were described as nonviable, whereas those without red fluorescence were counted as viable.
Plasma membrane integrity
For plasma membrane integrity assessment, the Hypo Osmotic Swelling test (HOST) was performed. 44 Five hundred microliters of HOST solution (sodium citrate 0.735 g, fructose 1.35 g, and 100 mL of distilled water) were mixed with 50 μL of semen and incubated at 30°C in an incubator for 30 minutes. A drop of 5 μL of semen sample was placed on a glass slide and observed at 400 × magnification under a phase-contrast microscope (BH-2; Olympus, Japan). A minimum of 100 sperm were observed. Sperm having intact plasma membrane showed coiling of the tail 45 were considered to be biologically active.
Sperm acrosome integrity
The normal apical ridge (NAR) test was performed for the assessment of post-thaw acrosomal integrity. Fifty microliters of 1% formal citrate solution was mixed with 500 μL of semen sample. One hundred spermatozoa were counted at 100 × using a phase contrast microscope (BH-2; Olympus). Sperm with a crescent shape appearance on the apical ridge were considered as having a normal acrosomal membrane. 45
DNA integrity
The acridine orange (AO) staining method was used for the evaluation of sperm DNA integrity. In brief, 50 mg AO (Cat. No. A6014-10 g; Sigma-Aldrich) was dissolved in 10 mL of distilled water to prepare a stock solution and stored in the refrigerator. One milliliter of AO stock solution and 0.5 mL of glacial acetic acid were added to 50 mL of distilled water to prepare a working solution. The semen extender was removed by adding distilled water to a 100 μL semen sample and then centrifuged at 500 g for 7 minutes. A thin smear was prepared using a 10 μL droplet from the sample on a glass slide. The air-dried slide was then fixed by placing in Carnoy's solution (glacial acetic acid 25% v/v and ethanol 75% v/v) at 4°C for 2 hours.
After fixation, the slide was dried in air and placed in AO stain for 3 minutes. The glass slide was then rinsed with water and observed under a fluorescence microscope (Olympus BX41). Sperm having normal DNA fluoresced green at the head region, whereas sperm with yellow to red fluorescence were considered to be the sperm with damaged DNA. 46
Mitochondrial membrane potential
Mitochondrial membrane potential (MMP) was assessed as described 47 with slight modification. In brief, 50 μL of a semen sample was diluted by adding 50 μL Tris critic acid fructose buffer and centrifuged at 300 g for 5 minutes. The pellet containing sperm was suspended again with Tris critic acid fructose buffer to make a final volume of 245 μL. The suspension was mixed with 5 μL (10 μg/mL) Rhodamine (Cat. No. R6626-25G; Sigma-Aldrich) and incubated in the dark for 20 minutes at room temperature. A 10 μL drop of a sample mixture was placed onto a glass slide and observed under a fluorescence microscope (Olympus BX41). A total of 200 sperm were counted. Those having green fluorescence at the mid piece were counted as sperm having intact mitochondria.
Survival kinetics of progressive sperm motility
Semen was incubated at 37.8°C after thawing and analyzed for the motility parameter at 0, 1, 2, 3, 4, 5, 6, 7, and 8 hours of incubation. The threshold limit for progressive sperm motility was set at 30% following the recommendation of the Theriogenology Society. 6
Biochemical evaluation
Catalase activity
The reported procedure 48 was followed for cell lysis with slight modification. After thawing at 37°C for 30 seconds, the semen straws were placed in a glass beaker and sonication was performed using a BMS Ultrasonic Liquid processor (ULP-750). Eight cycles with 30-second cooling periods were performed for complete sonication. After sonication, the semen samples were placed in 3 mL Eppendorf tubes and centrifuged at 3000 rpm for 5 minutes. The supernatant (50 μL) was collected in a cuvette containing 500 μL of H2O2 (test sample) and into another cuvette holding 500 μL H2O (control test). In addition, a standard (500 μL H2O2, 50 μL H2O) and a blank cuvette (550 μL H2O) were also prepared.
All cuvettes were incubated at 37°C for 3 minutes and ammonium molybdate (2 mL) was supplemented to stop further chemical reaction. A 10 μL sample was taken from each cuvette and observed for absorbance at 374 nm with a spectrophotometer (SPECORD 200 plus). The following equation was used for the calculation of catalase activity:
where t = total time, S° = absorbance of standard tubes, S = absorbance of test tubes, M = absorbance of control test, VT = total volume of reagents in test tubes, and VS = volume of semen. 49
Lipid peroxidation
The thiobarbituric acid reactive substances assay was performed for the evaluation of lipid peroxidation (LPO). Semen (200 μL) was mixed with 200 μL of 8% sodium dodecyl sulfate, 750 μL of 20% acetic acid, and 1.5 mL thiobarbituric acid. Distilled water was added to make the final volume of 4 mL. The mixture was heated at 95°C for 1 hour and then cooled to room temperature. After the addition of 1.5 mL N-butanol, the mixture was centrifuged at 4000 rpm for 15 minutes. The supernatant was used to measure the absorbance at 532 nm against the blank. A standard curve prepared using the known concentration of 1,1,3,3-tetraethoxypropane was used to calculate the malondialdehyde (MDA; nmol/mL) concentration of the sample and described as nmol/mL. 50
In vivo fertility rate
Based on better post-thaw quality parameters, BT3-treated semen samples were selected and compared with semen samples extended with conventionally used semen extender (TRIS-Egg Yolk) and previously used milk semen extender for in vivo fertility. Adult cyclic Nili-Ravi buffaloes (n = 325) of mixed parity (second to third lactation) having no palpable reproductive issues were selected for artificial insemination. The buffaloes (minimum 100 animals/group) were inseminated after 24 hours of standing heat without any hormonal intervention. All the animals after inseminations were kept under observation for returning to heat. Pregnancy diagnosis was performed after 60 days of A.I. through rectal palpation by an experienced technician. Buffaloes were declared pregnant based on the presence of a fetus.
Statistical analysis
The Shapiro–Wilk test was used to check the normality of data. The data for post-thaw semen quality parameters were analyzed through a 2 × 5 factorial analysis of variance [two types of milk (cow and buffalo milk) × 5 thermal treatments of milk (90°C, 100°C, 110°C, 120°C, and 130°C)]. The results of this study are presented as mean ± standard error of the mean. Bulls and replicates were designated as random variables. Multiple group comparison was performed by applying Tukey's test and p < 0.05 was considered significant. Chi-square analysis was performed to compare in vivo fertility data. The Statistical Analysis System, University Edition (SAS Institute, Inc., Cary, NC) was used to perform all statistical analyses.
Results
The data on the effect of heat treatments on reactive SH group's formation in cow and buffalo milk are given in Figure 1. An increasing trend of concentration of the SH groups was recorded with the rise in temperature in each type of milk. Figure 1 shows that the buffalo milk has a significantly higher (p < 0.05) SH concentration in each group (BT1, BT2, BT3, BT4, and BT5) where milk was treated at different temperatures (90°C, 100°C, 110°C, 120°C, and 130°C), respectively, than cow milk heated at the same temperatures.
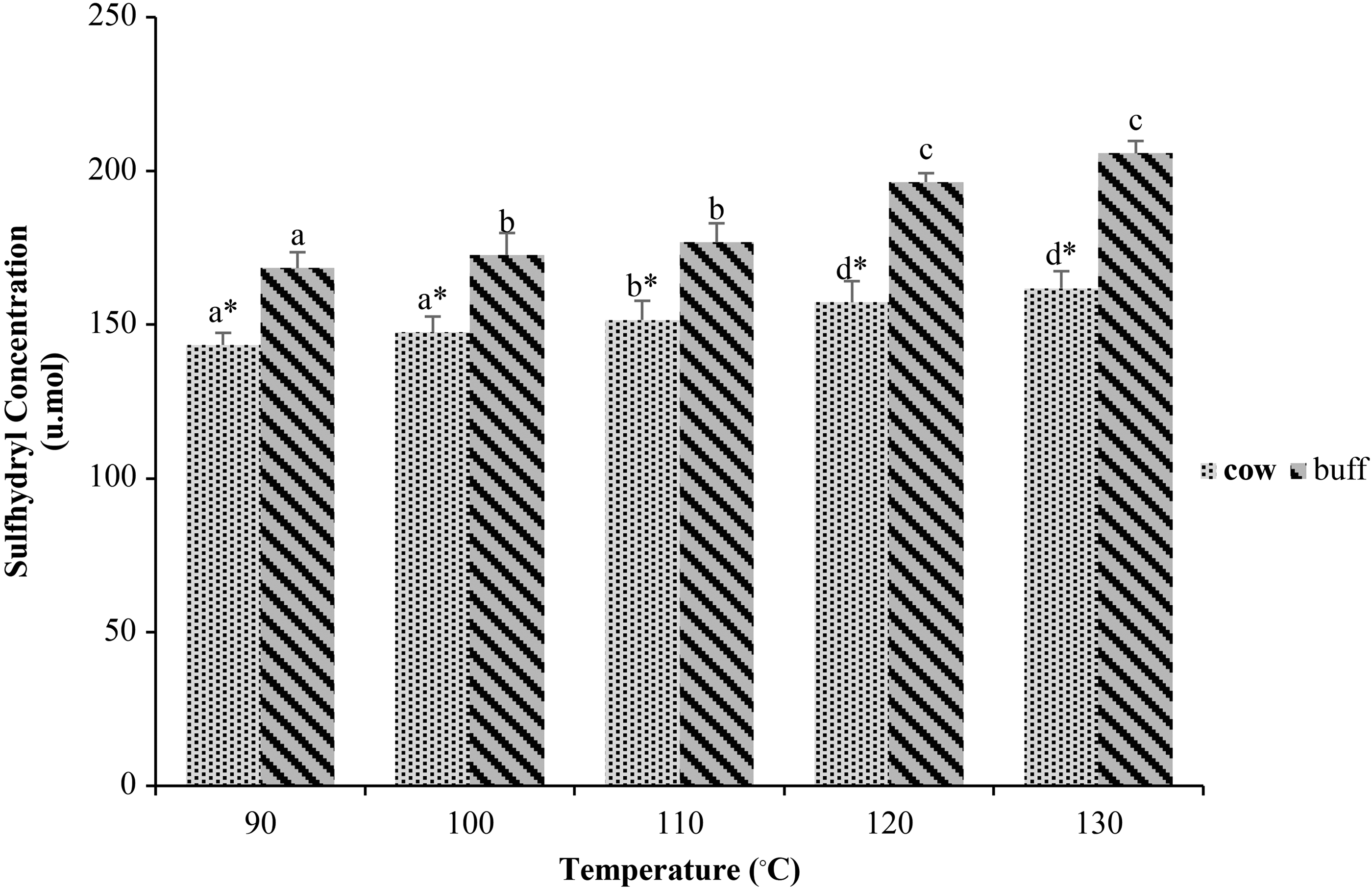
Reactive sulfhydryl concentration in cow and buffalo heat-treated milk. Data were analyzed through factorial ANOVA for two types of milk [cow and buffalo] × 5 thermal treatment (°C) [90, 100, 110, 120, and 130] and presented as mean ± SEM of six independent replicates. Bars with lowercase letters (a–d) represent a significant difference (p < 0.05) among thermal treatments in the same milk type, whereas bars with “*” represent a significant difference (p < 0.05) between milk types at the same thermal treatment for each parameter. ANOVA, analysis of variance; SEM, standard error of the mean.
The data for post-thaw total and progressive motility (%) after cryopreservation in heat-treated milk-based extenders are given in Figure 2. These parameters differed (p < 0.05) owing to the interaction of milk type and level of heat treatments. An increasing trend was observed in both parameters up to the T3 group, where both milks (cow and buffalo) were heated at 110°C. However, there was a decrease in total and progressive motility (%) as the temperature increased.
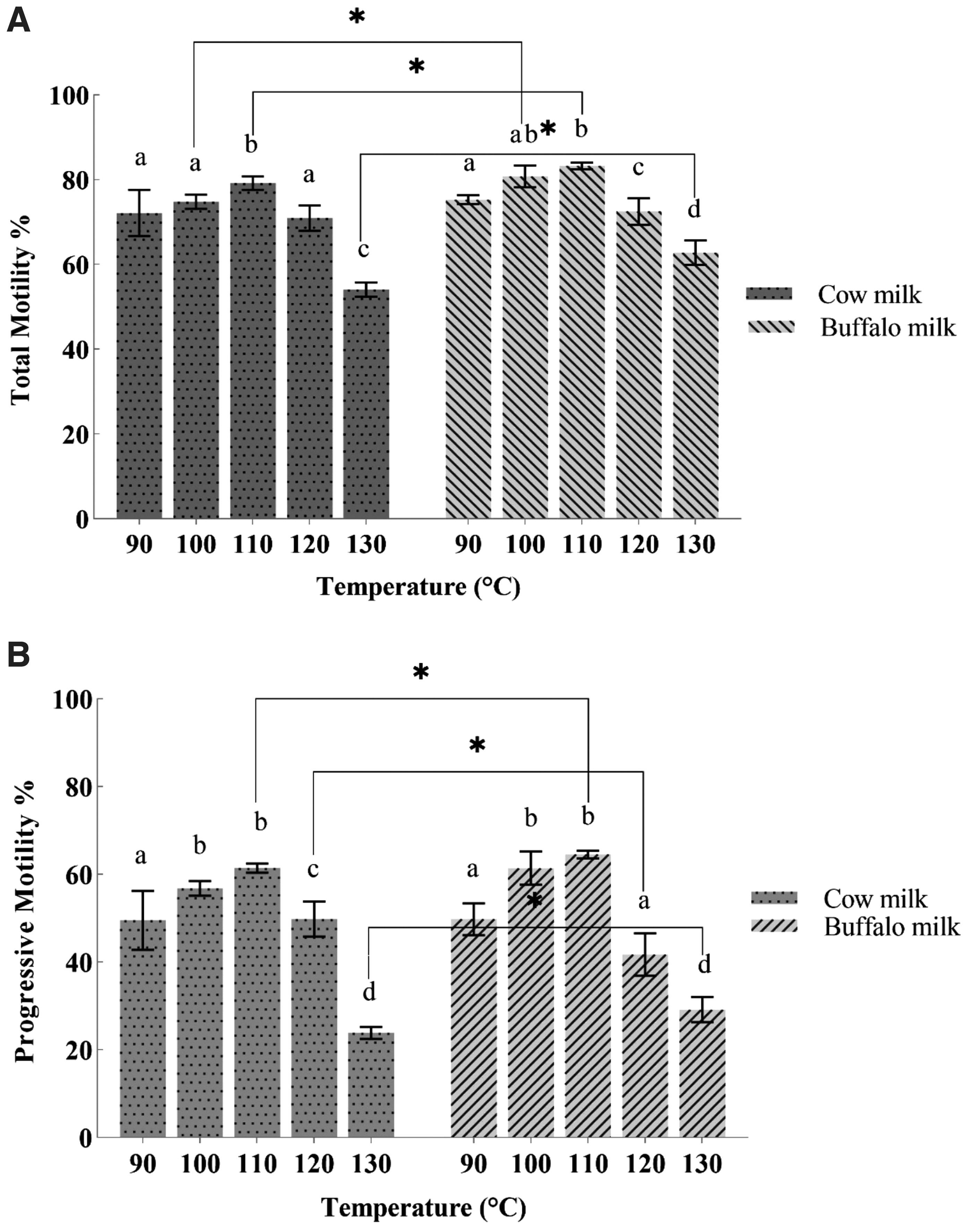
Post-thaw total motility
Figure 2 illustrates that total motility (%) was significantly higher (p < 0.05) in BT2 and BT3 where buffalo milk was heated at 100°C and 110°C, respectively, than cow milk heated at the same temperatures and the control group (BT1), whereas progressive motility was significantly higher (p < 0.05) in BT3 (where buffalo milk heated at 110°C) than the CT3 group, where cow milk was heated at the same temperature and all other temperature groups. Both parameters were found to be lowest (p < 0.05) in the treatment groups CT5 and BT5 (where temperature was highest, 130°C).
Furthermore, data for velocity distribution and kinematics are given in Table 1. Both the average path velocity and straight-line velocity (μm/s) were found to be higher (p < 0.05) in the BT3 (buffalo milk; 110°C) group than the control (buffalo milk; 90°C) and other treatment groups. Linearity (%) and straightness (%) were also found to be higher (p < 0.05) in the same group BT3 (buffalo milk; 110°C) than the other treatment groups and control.
Effect of Different Heat Treatments on Two Types of Milk Used for Extender Preparation on Velocity Distribution and Kinematics
Factorial ANOVA for two types of milk [cow and buffalo milk] × 5 heat treatments (°C) [90, 100, 110, 120 and 130] and presented as mean ± SEM of six independent replicates.
Lowercase letter superscripts (a–e) represent difference (p < 0.05) between heat treatments groups within same milk type, whereas symbol (*) represents difference (p < 0.05) between milk types within same heat treatment groups for each post-thaw quality parameter respectively.
Velocity distribution and kinematics: VCL = curvilinear velocity, VSL = straight line velocity, VAP = average path velocity, DCL = distance curved line, DSL = distance straight line, LIN = linearity, STR = straightness.
ANOVA, analysis of variance; SEM, standard error of the mean.
Results for post-thaw cell viability (A), acrosome integrity (B), plasma membrane integrity (C), and MMP (D) are given in Figure 3. Sperm viability (%), was not significantly different in the BT2 and BT3 groups (where buffalo milk was heated at 100°C and 110°C, respectively) from the cow milk groups heated at the same temperatures. However, MMP (%) was significantly higher in BT3 (buffalo milk; 110°C) group than in the CT3 group (cow milk; 110°C), and plasma membrane integrity (%) was significantly higher in BT2 (buffalo milk; 100°C) than in the CT2 group. Acrosome integrity (%) was significantly higher (p < 0.05) in BT3 (buffalo milk; 110°C) group than in the CT3 (cow milk; 110°C) group and other treatment groups. It was significantly lower in BT5 and CT5 groups where both milk samples were heated at 130°. Figure 4 illustrates the data for post-thaw DNA integrity (%). It was observed that there was a difference (p < 0.05) owing to interaction between milk type and thermal treatment.
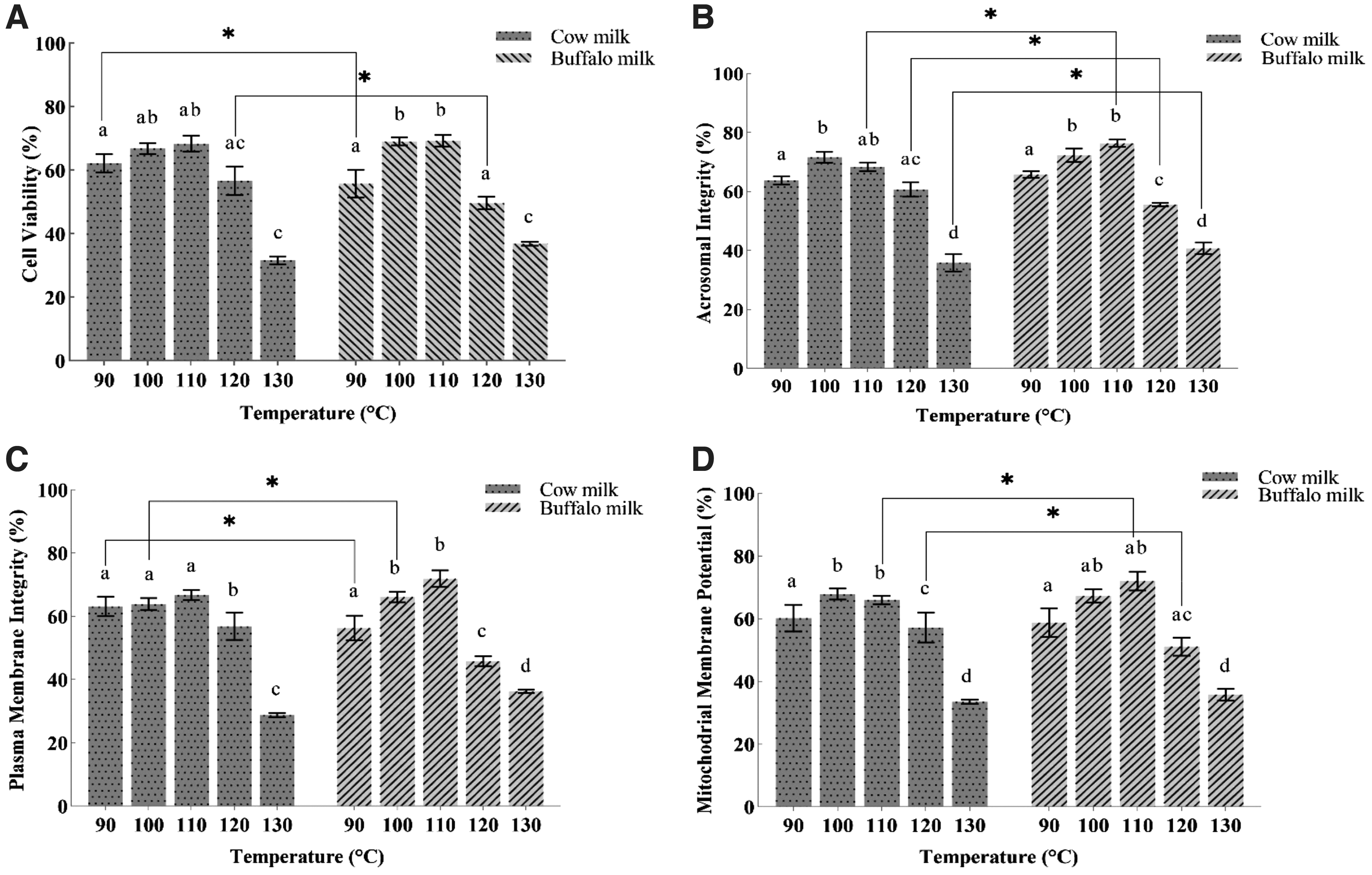
Cell viability
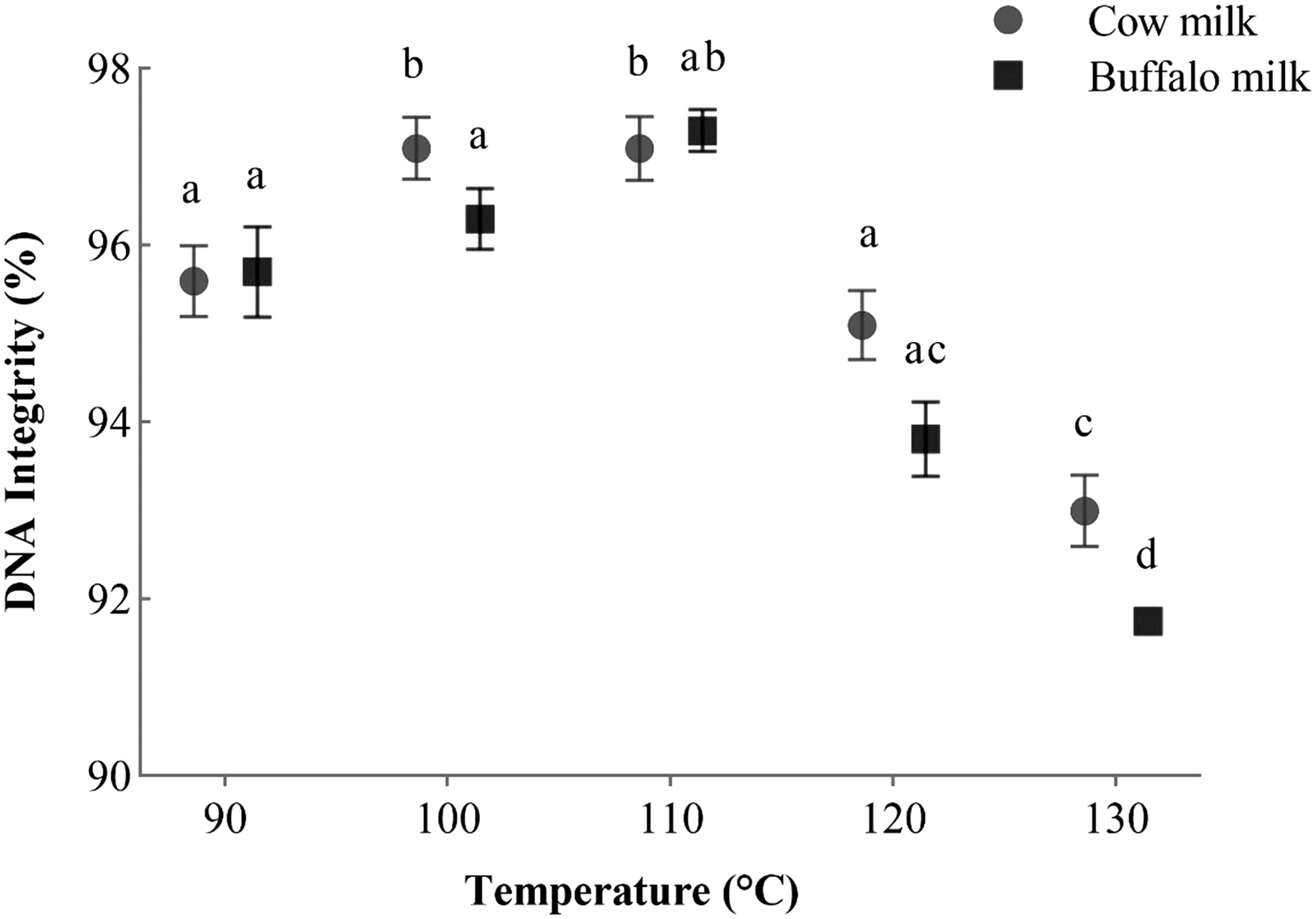
DNA integrity of frozen–thawed buffalo semen. Data were analyzed through factorial ANOVA for two types of milk [cow and buffalo] × 5 thermal treatment (°C) [90, 100, 110, 120, and 130] and presented as mean ± SEM of six independent replicates. Bars with lowercase letters (a–d) represent a significant difference (p < 0.05) among thermal treatments in the same milk type, whereas bars with “*” represent a significant difference (p < 0.05) between milk types at the same thermal treatment for each parameter.
DNA integrity (%) was found to be higher (p < 0.05) in CT2, CT3, and BT3. However, BT5 showed a lower (p < 0.05) DNA integrity percentage where buffalo milk was heated at 130°C than all other treatment groups and the control group.
The data for post-thaw biochemical profile (A) LPO and (B) catalase activity are given in Figure 5. An increasing trend of catalase activity was observed with the increase in temperature up to the 110°C group and then there was decrease in the activity as the temperature was increased further up to 130°C. Moreover, the MDA level was found to be steady up to 110°C temperature group. But it increased significantly as the temperature increased above 110°C. Figure 5 illustrates that MDA level (LPO) was lower (p < 0.05) in the BT3 group, whereas catalase activity was higher (p < 0.05) in BT3 and CT3 groups than all the other heat-treated groups and control group.

Biochemical profile
It has been revealed by Kaplan–Meier log-rank survival analysis that the survival time to reach the threshold limit of sperm progressive motility differed owing to the interaction of milk types and heat treatments. It was higher (p < 0.05) in CT3 (cow milk; 110°C) and BT3 (buffalo milk; 110°C) groups (Fig. 6). The survival function of sperm progressive motility in BT3 was reduced to 80% at 6 hours, 40% at 7 hours, and 0% at 8 hours, whereas the survival function of sperm progressive motility in CT3 was reduced to 40% and 0% at 7 and 8 hours, respectively.

Survival function of progressive sperm motility of Nili Ravi semen cryopreserved in buffalo milk semen extenders
The results for in vivo fertility rates of buffalo spermatozoa cryopreserved in BT3 (having best post-thaw quality parameters), CT1 (cow milk; 90°C) (control group; milk-based extender), and Tris Egg Yolk Extender TEYE (commonly used extender) are given in Table 2. The fertility rate was significantly higher (p < 0.05) in buffaloes inseminated with semen cryopreserved in BT3 (buffalo milk; 110°C) (47%) than CT1 (cow milk; 110°C) (30%) and TEYE (36%).
Comparison of In Vivo Fertility of Buffalo Semen Cryopreserved in TEYE, CT1, and BT3 Extenders
Difference (p < 0.05) between groups.
Different types of extenders for semen cryopreservation: TEYE = Tris egg yolk extender, CT1 = cow milk heated at 90°C, BT3 = buffalo milk heated at 110°C.
Discussion
A growing body of evidence suggests that heat treatment of milk increases its antioxidant potential provided by reactive SH groups.35,51–55 We hypothesized that these reactive SH groups are supposed to be more efficient and less cytotoxic antioxidants when used in a cryopreservation medium, thus, enhancing the post-thaw quality and in vivo fertility of buffalo bull spermatozoa.
The most salient outcome of this study is the significant reduction in post-thaw MDA production that is considered a biomarker for LPO, when buffalo milk was used for the preparation of semen extender after heating at 110°C (BT3). This reduction in the MDA level is owing to decreased LPO of PUFAs present in the plasma membrane of spermatozoa. 56 Few studies have reported that the antioxidant property of milk increases with heat treatment because of protein unfolding and exposure of the reactive SH groups. These groups act as hydrogen donors.36,57 The reactive SH groups directly react with ROS and protect the cell membrane from damage through masking the ROS, as also reported by Genet et al. 55
Furthermore, during LPO, membrane integrity goes through irreversible changes and cell permeability is increased, which result in a decreased level of antioxidant enzyme activity.58,59 Hence, the high LPO of cell membranes during cryopreservation induces increased membrane leakiness and consequently, a loss of intracellular antioxidant enzymes occurs. However, in the present experiment, the antioxidant activity (catalase) was recorded to be higher when buffalo semen was cryopreserved in the BT3 group. This higher catalase value is attributed to higher membrane stability owing to less damage by ROS, and a low level of subsequent LPO. The lower levels of LPO have been reported in cryopreserved sperm in the presence of a potent antioxidant system.25,60
The sperm cell membrane has been shown to be affected during cool storage.61–64 In this study, a significant positive effect on post-thaw cell membrane integrity was obtained when heated milk of buffalo was used for extender preparation, which may be because of PUFAs, which are required to give the plasma membrane consistency that is essential for structural integrity. This structural integrity keeps the sperm viable. 65 Our data proved that heating buffalo milk at 110°C (BT3) before extender preparation produces an optimum level of reactive SH groups and increases post-thaw sperm viability in Nili-Ravi buffalo. The reason may be the protective effect of casein molecules along with glycerol augmented in the presence of high contents of reactive SH groups, resulting in less oxidative stress to the lipids of the sperm cell membrane.
Moreover, plasma membrane integrity, sperm viability, and motility have shown a negative correlation with MDA concentration.66,67 The increased reactive SH groups in heated milk directly react with the oxygen-free radicals 55 and efficiently mask their damaging effects. Subsequently, it may strengthen the intercellular antioxidant system, including catalase. 67
Furthermore, our results revealed that the NAR, acrosomal and MMP were also improved. It has previously been reported that electron transport couples with oxidative phosphorylation to maintain a high MMP 68 and intracellular ROS are mostly formed in mitochondria, which results in a disruption of electron transport. 69 Disrupted by excessive ROS production, sperm fail to maintain a high MMP, 70 and a remarkable decrease in MMP occurs. In this study, SH groups played a pivotal role in protecting the mitochondrial membrane by engaging the free radicals, through transfer of their hydrogen atoms, hence, neutralizing the ROS 71 and maintaining a high MMP. It is worth mentioning that intact plasma membrane acts like a barrier that protects the cellular structures, thus enhancing the sperm viability. 72
Sperm motility is considered as one of the major parameters for the prediction of fertility. It is positively associated with other post-thaw quality parameters including sperm viability, plasma membrane integrity, and MMP. 73 In this study, cryopreservation of Nili-Ravi buffalo sperm in buffalo milk semen extender (BT3) resulted in improved post-thaw total and progressive motility. There are several proposed mechanisms that might be responsible for the decreased sperm motility associated with oxidative stress, which include peroxidation of PUFAs, decreased ATP owing to a decrease in MMP, and so on. In our study, as already highlighted, the reactive SH groups are scavengers of free radicals, reacting directly or by transfer of hydrogen atoms to ROS, thus counteracting their deleterious effects and improving the post-thaw total and progressive motility of the cryopreserved buffalo bull sperm.
It is a well-known fact that bulls having <3% of DNA damage results in high fertility rates. 74 However, freezing and thawing processes induce injuries to the chromatin structure of mammalian sperm.74,75 During cryopreservation, the production of excessive ROS is associated with DNA damage. 66 The results of this study showed that semen extender prepared from milk after heating at 110°C resulted in <3% damage to DNA.
In our study, the in vivo fertility rate was markedly improved when buffalo semen was cryopreserved in buffalo milk (BT3). Reduced oxidative stress may be taken as the key factor for this improvement because heat treatment improved the antioxidant potential of the milk.35,51 Another possible reason for this better fertility may be the high plasma membrane integrity. It is also worth mentioning that some exogenous antioxidants have deleterious effects on sperm genetic material. 76 Hence, these damages lead to lower fertility. Heat treatment of milk helped in the endogenous production of antioxidants species in milk extender and is considered to lessen the genetic damage on spermatozoa during cryopreservation and thawing, resulting in a high fertility rate.
It is evident from this study that post-thaw semen quality is better in medium that was prepared by using buffalo milk. The reason may be the higher protein concentration in buffalo milk (45.7 ± 1.2; g/kg) than cattle milk (33.6 ± 1.7; g/kg).77,78 So, the heat treatment of buffalo milk produced a higher concentration of reactive SH contents than cow milk. That is why buffalo milk is found to be a more efficient freezing medium than cow milk.
However, it was observed that the post-thaw quality of semen was decreased when it was diluted and preserved using milk heated at higher temperatures (120°C and 130°C), despite having higher reactive SH contents. This might be due to a decrease in the availability of casein, which plays a protective role for the sperm against freezing damage, along with glycerol. 79 Because of the heat treatment of milk, casein is considered to be trapped physically with other milk proteins. Moreover, high temperatures cause the attachment of some portions of whey protein to the casein micelles by calcium linkages and disulfide bridging. Consequently, casein remains in the micelles 80 and a lower amount becomes available to protect bull sperms.
However, there are some limitations in this experiment that could be addressed in future studies. First is the sample size of the bulls and second is the lack of experiments to observe the molecular changes in sperm structure during cryopreservation in heat-treated milk. Although we performed the viability assays and in vivo fertility trials to assess the survival of buffalo sperm, molecular level studies in the future can give further insights about the successful freezing of sperm in a milk-based extender.
In conclusion, this trial indicates that the cryopreservation of bull semen in buffalo milk extender prepared after heating at 110°C resulted in enhanced antioxidant capacity and less damage to the cellular structures and functional parameters of buffalo bull sperm, mainly by mitigating the oxidative stress. Subsequently, the in vivo fertility rate of Nili Ravi buffalo inseminated with semen cryopreserved in BT3 milk-based extender also improved.
Footnotes
Acknowledgments
The authors acknowledge the staff of Al-Haiwan SPU Sahiwal for the use of their bulls and facilities.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
No funding was received for this study.