
Abstract
In this study, the effects of trehalose and 1, 3-cyclohexanediol (1, 3-CHD) on the ultrastructure of frozen-thawed ram sperm were assessed and compared. In the control group, sperm were frozen without trehalose and 1, 3-CHD. In the trehalose group, 100 mM trehalose was used for sperm cryopreservation. In the cyclohexanediol group, the freezing extender contained 100 mM 1, 3-CHD. The transmission electron microscope (TEM) was used to observe the ultrastructural alterations of sperm. For verification of the TEM results, the plasma membrane and acrosome integrity of ram frozen sperm was assessed. Three fertility-proven rams were used in this study. Semen collection was repeated 6 times. The collected semen was pooled to preclude the individual difference each time. The sperm collected from a representative ram were used for ultrastructural observation. The TEM results indicated extensive and severe cryoinjuries on the main organelles of ram frozen sperm. Some alterations in plasma membrane, including detachment, rupture, dilation, or loss, appeared in post-thaw sperm. The bending shape and leakage of genetic materials were also observed in the nucleus. In addition, the outer acrosome membrane in some frozen sperm was broken or partly lost. Further, leakage of the inner contents of acrosomes also occurred. Sperm mitochondria was negatively influenced by cryopreservation. With 1, 3-CHD or trehalose, the percentage of sperm with normal acrosomes was 62% or 64%, and it was significantly higher than that of the control (41.51%, p < 0.05). However, different from trehalose, 1, 3-CHD cannot efficiently protect the post-thaw integrity of the plasma membrane (48.09% vs. 26.92%, p < 0.05). The TEM results were consistent with the quality assessment of frozen-thawed sperm. Collectively, trehalose and 1, 3-CHD can mitigate cryoinjuries on sperm ultrastructure. The cryoprotective effects of trehalose on sperm plasma membrane are superior to 1, 3-CHD. Sperm plasma membrane is the most sensitive to cryoinjuries, followed by acrosomes, mitochondria, and nuclei.
Introduction
Investigators have attempted to reduce cryoinjury and improve cryosurvival of mammalian sperm since the cryoprotective effects of glycerol on fowl sperm were confirmed in 1949. However, in spite of progress obtained during the past 70 years, 50% of mammalian sperm lose their viability after a freezing and thawing process.1,2 Many factors, such as ice formation, chemical toxicity, and oxidative stress, can compromise the fertility of frozen sperm.3–5 In addition, the sperm plasma membrane has been proved to be particularly sensitive to external stresses, consequently leading to poor viability and fertility.4–8
Sperm cryopreservation is an important technology for the establishment of animal genetic resource biobanks and their use for artificial insemination (AI). However, owing to a specific lipid composition in their plasma membranes, ram sperm are sensitive to cryoinjuries caused by the cryopreservation process.3,9,10 In accordance with some previous investigations, the traditional cervical AI using post-thaw ram semen resulted in a poor pregnancy rate.11,12 In the dairy sheep industry, fresh or chilled semen is generally used for AI to guarantee a relatively higher pregnancy efficiency. 13
To reduce negative effects of the freezing and thawing process on ram sperm, oligosaccharides, such as trehalose and sucrose, have been used for cryopreservation of ram sperm.6,7,14–20 The cryoprotective effects of trehalose have become a hot topic in the field of cryobiology. Further, mainstream opinions support the cryoprotective roles of trehalose in ram sperm during the freezing and thawing process.6,14,15,17,19,20 In addition, in previous studies, some synthetic ice blockers (SIBs), such as 1, 3-cyclohexanediol (1, 3-CHD) and 1, 4-cyclohexanediol (1, 4-CHD), have been used for the cryopreservation of ram sperm, indicating their cryoprotective roles.8,21 However, the freezing and thawing process still seriously damages the structures of ram sperm, especially the plasma membrane.
Morphological observation is also an important approach to evaluate the post-thaw quality of mammalian sperm. In current investigations, sperm morphology is generally observed by using an optical microscope after the smearing of semen on a glass slide. 22 However, this method can only assess the approximate motility of sperm and cannot precisely observe sperm structural alterations. In addition, aided by fluorescent microscopy or flow cytometry, some specific fluorescent probes, such as Annexin-V, propidium iodide (PI), peanut agglutinin (PNA), pisum sativum agglutinin (PSA), and tetrechloro-tetraethylbenzimidazol carbocyanine iodide (JC-1), have been used for the detection of subcellular and functional alterations in sperm organelles, such as plasma membrane, acrosomes, and mitochondria.8,10,13,23,24
However, some limitations still exist. First, there is not a standardized procedure for the fluorescent probes that are used, due to different requirements of the excitation wavelength and the emission wavelength among these probes. Therefore, the status assessment of an individual sperm cell cannot be performed in a same visual field. 25 Second, before staining with fluorescent probes, sperm cells generally need a complex treatment, such as chemical fixation, repeated washing, and dilution. These treatments can also influence the analytical results of sperm. For instance, the dilution procedure aggravated the structural injury on the sperm plasma membrane owing to dilution shock. 21
In contrast, the electronic microscopy technology, especially transmission electron microscopy, can precisely observe the ultrastructural alterations of main sperm organelles.5,26,27 Currently, transmission electron microscope (TEM) has been applied to assess the effects of cryopreservation on mammalian sperm in some sporadic studies.6,22,28,29 According to the study carried out by Aisen et al., trehalose efficiently protected the plasma membrane of frozen-thawed ram sperm via observation by TEM. 6 In another study, with the aid of the TEM technology, López Armengol et al. developed an approach that was used to describe 29 morphological categories of sperm heads based on different regions in fresh, chilled, or post-thaw ram sperm.
The authors confirmed that cryoinjuries were mainly present in the plasma membrane and acrosome. 22 In goats, after freezing and thawing, the sperm nucleus was generally characterized by ruptures, bending, and deformities. Moreover, sperm plasma membranes were ruptured or fully lost. 28 In bovines, the ultrastructural alterations of freeze-dried sperm were also observed in a study carried out by Martins et al. Their results indicated that the negative effects of the freeze-drying process on sperm plasma membrane were most severe. However, the acrosomes and mitochondria of freeze-dried sperm were well conserved. Moreover, trehalose can efficiently protect the acrosomal integrity of freeze-dried bovine sperm. 25
In our previous studies, the freezing and thawing procedure of ram sperm was optimized through the addition of trehalose or SIBs in freezing extenders.8,19 In this study, the TEM technology was applied to observe the post-thaw ultrastructural alterations of ram sperm. According to our knowledge, there are a few reports using the TEM technology to evaluate ram sperm frozen with these cryoprotectants. This study can help us precisely evaluate the impacts of the cryopreservation process on the ultrastructure of ram sperm, thus providing an important direction for further optimization of the freezing procedure of ram sperm.
Materials and Methods
Ethics statement
In the present study, all treatments including ram application were permitted by the ethical committee of Yunnan Animal Science and Veterinary Institute (YASVI; 201506008). Further, during the study, all authors strictly complied with the Regulations on the Administration of Laboratory Animals (Order-No. 2 of the State Science and Technology Commission of the People's Republic of China, 1988) and the Regulations on the Administration of Experimental Animals of Yunnan Province (issued by the Standing Committee of Yunnan Provincial People's Congress 2007.10).
Semen extenders
Unless otherwise stated, all chemicals and kits used in this study were purchased from Sigma Chemical Co. (St. Louis, MO).
The basic extender used in this study was composed of Tris (224.0 mM), citric acid (66.6 mM), and glucose (55.5 mM). Three types of freezing extenders were used and named as CON, TRE, and SIB, respectively. The CON freezing extenders consisted of 224.0 mM Tris, 66.6 mM citric acid, 55.5 mM glucose, 10% (V/V) egg yolk, 5% (V/V) glycerol, 100,000 IU penicillin, and 100,000 IU streptomycin. The TRE or SIB freezing extender was the CON extender supplemented with 100 mM trehalose or 100 mM 1, 3-CHD. The freezing extenders were adjusted to pH 7.0 by using Tris.
The crystal osmolarity of the basic extender or the freezing extenders was measured by using the FISKE ONE-TEN osmometer (FISKE Associates, Norwood, MA). Since this osmometer cannot measure colloid osmolarity, the effects of egg yolk and glycerol on measurement of crystal osmotic pressure should be precluded in advance. The crystal osmolarity of the freezing extenders (CON, SIB, and TRE) in which egg yolk and glycerol were replaced by distilled water was 326, 425, and 421 mOsm, respectively. After centrifugation at 10,000 × g for 40 minutes at 4°C, the supernatants were collected and further filtered through a 0.45 μm membrane filter (Millipore Corporation, Bedford, MA).
Semen collection, evaluation, and dilution
The study was conducted at a farm belonging to YASVI, located in the Kunming City, Yunnan Province, China (103°40′E; 26°22′N). The Yunnan semi-fine wool rams were raised in an environment with uniform feeding, housing, and light conditions. The rams had free access to water and salt lick.
Semen samples were collected from three fertility-proven rams (2–4 years of age) by using electric stimulation. After collection, the ejaculates were directly placed in a 37°C water bath and transported to our laboratory within 15 minutes. The semen quality was assessed immediately. The ejaculates used in this study should meet the following criteria: volume of 0.75–2 mL, minimum sperm concentration of 2 × 109 sperm/mL, and motility higher than 75%. The semen was diluted in the freezing extenders. The concentration of sperm in the diluted semen was adjusted to 2 × 108 sperm/mL. The sperm concentration was measured with NucleoCounter® SP-100™ (Chemo-Metic AS, Allerød, Denmark).
Cooling, freezing, and thawing
The diluted semen was loaded into the 0.25 mL plastic straws (IMV) and then cooled to 5°C at 0.40°C/min. After equilibration at 5°C for 2 hours, the straws were placed on an aluminum plate (10 cm × 2 mm × 40 cm) and frozen in liquid nitrogen vapor for 10 minutes. The freezing rate was ∼100°C/min. The distance between the aluminum plate and the surface of liquid nitrogen was 4 cm. After freezing in liquid nitrogen vapor, the straws were plunged into liquid nitrogen and preserved for 1 month.
In this study, the cooling and freezing rates were measured by using a thermometer (MCT-200; Yushi Technology Development Co., Beijing, China). The thermocouple probe was inserted into the middle of a plastic straw to measure the temperature fluctuation. Meanwhile, the cooling time (from 37°C to 5°C) and the freezing time (from 5°C to −50°C) were also recorded. The cooling rate was calculated by using the following formula: (5°C–37°C)/the cooling time (minutes). The freezing rate was calculated by using the following formula: (−50°C to 5°C)/the freezing time (minutes). The frozen ram semen was thawed at 37°C for 30 seconds.
Quality assessment of frozen-thawed sperm
The acrosome status of ram sperm was assessed by using fluorescein isothiocyanate-labeled PSA (FITC-PSA) together with flow cytometry. The operating procedure has been previously described.8,13,23,30 The effects of the freezing and thawing procedure on the membrane integrity of ram sperm were determined by using an Apoptosis Detection Kit (KeyGEN Biology, Nanjing City, China). The operating procedure was described in a previous study carried out by Wu et al. 31 After flow cytometry, sperm stained with PI were considered to be dead sperm.
The hypo-osmotic swelling test (HOST) was used for the quality assessment of sperm. The procedure was based on the investigations of Matsuoko et al., 17 with some minor modifications. Briefly, 10 μL of post-thaw semen was mixed equally with 100 μL of a hypotonic medium containing 5 mM fructose and 2.55 mM sodium citrate. After incubation at 37°C for 30 minutes, 20 μL of the sperm suspension was smeared on a slide and then covered with a cover slip. Four hundred sperm were counted by using an inverted microscope equipped with the computer-assisted sperm analysis system (MaiLang, Songjingtianlun Co., Nanning City, China). The percentage of sperm with a looped tail was calculated by the following formula: the number of sperm with looped tails/400 × 100%.
Transmission electron microscopy
The ultrastructural observation of ram sperm was conducted by the TEM technology. The thawed sperm were washed three times by centrifugation for 10 minutes at 600 × g in the phosphate buffered saline (GE Healthcare Life Sciences, Hyclone Lab, South Logan, UT) at 4°C. After removal of the supernatant, the cold 3.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.2–7.4) was slowly supplemented to the cell pellets. After fixation for 48 hours at 4°C, the samples were then washed three times in 0.1 M cacodylate buffer. The cell pellet was further fixed in 1% osmium tetroxide for 1 hour and then washed with 0.1 M cacodylate buffer for 15 minutes.
After fixation in osmium tetroxide, the specimens were dehydrated through a gradated series of ethanol and acetone according to the standard procedure, and they were further infiltrated with Epon-618 for 24 hours. The thin sections were cut with the Leica R ultramicrotome (Leica Microsystems Ltd., Heidelberg, Germany) at a thickness of about 60–70 nm. The sections were sequentially stained with 4% uranyl acetate and 4% lead citrate for 10 minutes in the darkness. Finally, the sections were observed under a Jeol JEM-1011 electron microscope (Jeol Ltd., Akishima, Japan).
The percentage of abnormal membranes or acrosomes in frozen-thawed sperm was statistically analyzed. The feature of sperm with a damaged membrane included detachment from underlying sperm structure, dilation, rupture, and partial or full loss. The feature of sperm with damaged acrosome included abnormal outer acrosomal membrane and leakage of inner content. In this study, ∼50 cells were observed by using TEM. The percentage of sperm with normal membrane or acrosome was calculated by the following formula: the number of sperm with normal membrane or acrosome/the counted cell number × 100%.
Statistical analysis
All statistical analyses were performed by using the SPSS Software (version 11.0.1; SPSS, Inc.). The data related to the quality evaluation of the post-thaw sperm were collected from three rams in this study. Each time, the collected semen was pooled to preclude the individual differences. The experiment was repeated six times. The data were expressed as means ± standard error of the mean, and they were analyzed by one-way analysis of variance with calculation of the least significant difference. All results with a value of p < 0.05 were considered statistically significant.
The analysis of ultrastructural alteration was performed by using sperm derived from a representative ram. Approximately 50 cells were observed under the TEM. The normal cells and the damaged cells were counted separately. The chi-square test was used to statistically analyze these data. All results with a value of p < 0.05 were considered statistically significant.
Results
Quality assessment of frozen-thawed ram sperm
The results related to the post-thaw quality of ram sperm are presented in Table 1. The percentages of sperm with intact acrosomes after freezing with trehalose or 1, 3-CHD showed a significant increase as compared with the control group (p < 0.05). Further, no significant difference was found between the TRE group and the SIB group, regarding acrosomal status (p > 0.05). However, in the presence of trehalose, the percentage of frozen-thawed sperm with intact membrane or looped tail was significantly higher than the other two groups (p < 0.05). However, there is no significant difference between the SIB group and the control group (p > 0.05).
Effects of Trehalose and 1, 3-Cyclohexanediol on Plasma Membrane and Acrosome Integrity of Frozen-Thawed Ram Sperm
The data in the same column with no or same superscripts indicate no significant difference existing among the treating groups (p > 0.05).
Significant difference (ANOVA-LSD test, p < 0.05).
“CON” represents the control group in the absence of trehalose or 1, 3-CHD; “SIB” represents the group frozen with 1, 3-CHD; “TRE” represents the group frozen with trehalose.
1, 3-CHD, 1, 3-cyclohexanediol; ANOVA, analysis of variance; LSD, least significant difference.
Ultrastructural alteration of frozen-thawed ram sperm
The effects of the freezing and thawing procedure on the main organelles of ram sperm are shown in Figure 1. Here, we used the CON group to observe the post-thaw ultrastructural changes. The TEM results indicated extensive abnormal alterations on plasma membranes of post-thaw ram sperm. After freezing and thawing, the plasma membrane was dilated or detached from underlying structures in the head or tail region of some ram sperm (Fig. 1A, C, E). Most importantly, the rupture and loss of plasma membranes were also observed in post-thaw sperm (Fig. 1C, E).
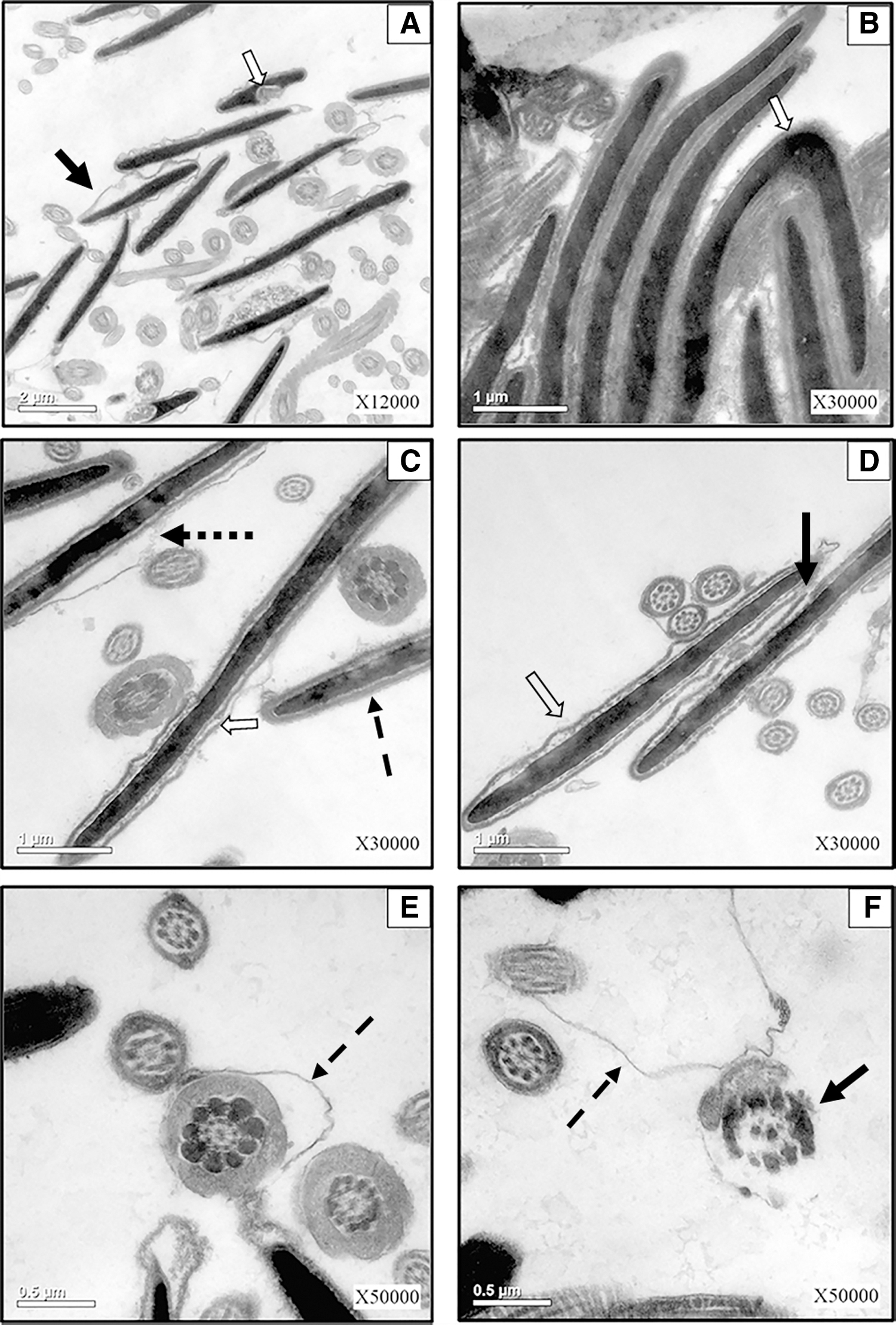
The effects of the freezing and thawing process on the ultrastructure of ram sperm.
In addition to plasma membrane, the freezing and thawing process also influenced the nucleus shape of ram sperm. As shown in Figure 1A, cryopreservation led to partial leakage of nuclear content. Further, the nucleus shape of some frozen-thawed sperm showed bending, as indicated in Figure 1B. However, the reason still needs further elucidation.
This study also indicated that the sperm acrosome is susceptible to the freezing and thawing procedure. As shown in Figure 1C, the abnormal alterations in acrosomes, such as rupture or loss of the outer acrosome membrane, occurred in some frozen-thawed sperm. Further, partial or complete loss of acrosomal inner contents was also observed in post-thaw sperm (Fig. 1C, D).
The negative effects of the cryopreservation process on mitochondria of ram sperm are shown in Figure 1E and F. In the middle piece region, the plasma membrane of some sperm was detached from the underlying mitochondrial helix (Fig. 1E). Most severely, the mitochondrial structures in some sperm were completely destroyed after freezing and thawing, as shown in Figure 1F.
Effects of trehalose or 1, 3-CHD on ultrastructure of frozen-thawed ram sperm
The effects of trehalose or 1, 3-CHD on the plasma membrane of frozen-thawed ram sperm are presented in Figure 2. After freezing and thawing, the plasma membrane of most sperm showed a detachment from their heads. In some frozen-thawed sperm, the plasma membranes have been ruptured or lost. The presence of 1, 3-CHD cannot mitigate the negative effects of the freezing and thawing process on plasma membrane.
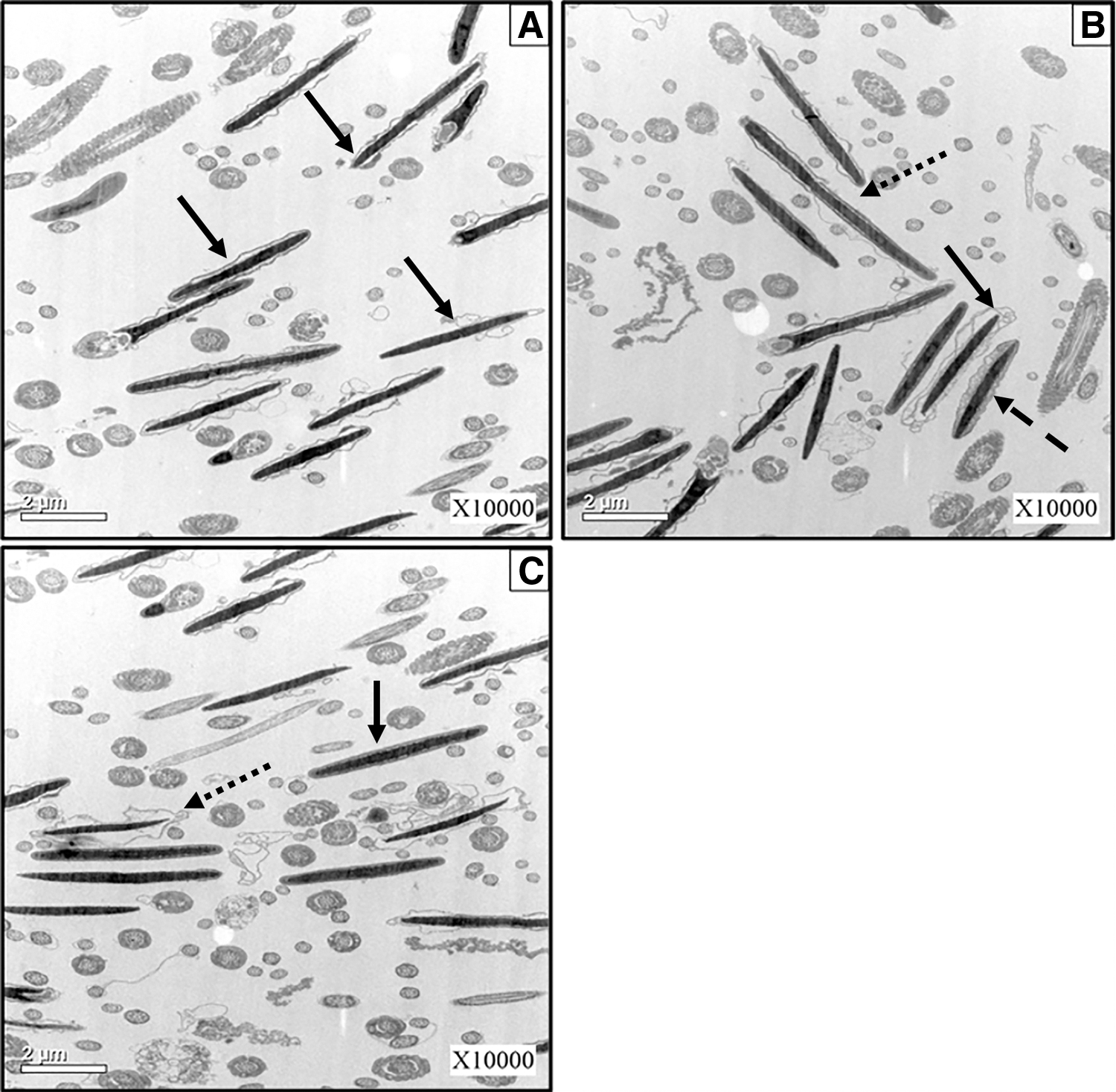
The effects of trehalose or 1, 3-CHD on plasma membrane of frozen-thawed ram spermatozoa (10,000 × ).
As shown in Figure 2B, although the plasma membranes in most of the frozen-thawed sperm remained intact, they have been detached from the sperm head. However, the ability of trehalose to protect the post-thaw membrane integrity of sperm was superior to that of 1, 3-CHD. As indicated in Figure 2C, the plasma membranes in most of the frozen-thawed sperm were intact and tightly attached to the sperm head. However, in a few post-thaw sperm, the separating phenomenon of plasma membrane from the sperm head still existed, showing severe damage of the cryopreservation process to the sperm plasma membrane.
With respect to the effects of the freezing and thawing procedure on the ultrastructure of sperm acrosomes, the comparison between trehalose and 1, 3-CHD is presented in Figure 3. In the absence of these two cryoprotectants, the inner contents of the acrosome in some post-thaw sperm partly or completely leaked, as shown in Figure 3A. However, when trehalose or 1, 3-CHD was added, the acrosomal ultrastructure of frozen-thawed sperm remained intact, although the plasma membrane in some sperm had been detached or dilated.

The effects of trehalose or 1, 3-CHD on acrosomal ultrastructure of frozen-thawed ram spermatozoa.
The TEM results were further statistically analyzed to quantitatively analyze the difference between these groups. The data related to sperm acrosome and plasma membrane are presented in Table 2. The percentage of sperm with normal acrosomes in the CON group was significantly less than that of sperm frozen with trehalose or 1, 3-CHD (p < 0.05). In addition, there is no difference between the TRE group and the SIB group regarding the acrosomal changes (p > 0.05). Regarding the plasma membrane, the post-thaw percentage of sperm with intact plasma membrane in the TRE group was significantly higher than that in the CON or SIB group (p < 0.05). However, no difference was found between the CON group and the SIB group (p > 0.05).
Effects of Trehalose or 1, 3-Cyclohexanediol on Plasma Membrane and Acrosome of Frozen-Thawed Ram Sperm Analyzed by Transmission Electron Microscopy
The data in the same column with no or same superscripts indicate no significant difference existing among the treating groups (p > 0.05).
Significant difference (chi-square test, p < 0.05).
Discussion
Ram sperm are particularly sensitive to the freezing and thawing procedure.2,7,10 Unlike bull semen, a standardized freezing approach has not been established for ram sperm.4,12 At present, most investigators tend to agree with the cryoprotective effects of trehalose on frozen ram sperm.6,7,17–20,32 In some previous studies, the cryoprotective effects of trehalose may have been superior to those of the other disaccharides, such as sucrose.19,23 In addition, as a potential replacement of antifreeze proteins, SIBs, such as 1, 3-CHD or 1, 4-CHD, have also been used for cryopreservation of ram sperm.8,21 Cyclohexanediol cannot interact with egg yolk and glycerol.
Similar to natural antifreeze proteins, cyclohexanediol may show its cryoprotective effects through modifying ice crystal shape and further reduce cryoinjuries during cryopreservation.8,21 In this study, the effects of trehalose and 1, 3-CHD on the ultrastructure of frozen-thawed ram sperm were further assessed. The TEM technology was used to observe the ultrastructural alteration of post-thaw sperm. Further, the data related to the effects of trehalose or 1, 3-CHD on ram sperm plasma membrane, acrosome, and osmotic tolerance were also provided to verify the results of TEM.
As indicated in Figure 1, the freezing and thawing process can lead to extensive and severe cryoinjuries on the main sperm organelles, including plasma membrane, acrosome, nucleus, and mitochondria. Among these abnormal alterations, the morphological change of sperm plasma membrane was especially obvious. Some abnormal alterations including detachment, rupture, and swelling of plasma membrane appearing in sperm after the cryopreservation process, confirming that the plasma membrane of ram sperm is extremely sensitive to cryoinjuries. Similar conclusions were also drawn in the investigations carried out by Aisen et al. 6 and Shi et al. 28
In kangaroo, an investigation carried out by McClean et al. indicated that cryoprotectants, such as glycerol or dimethyl sulfoxide (DMSO), particularly influenced the integrity of sperm plasma membrane, 33 indicating that the sperm plasma membrane is sensitive to the toxicity of permeable cryoprotectants. In addition, in accordance with the report of Plummer and Watson, cold shock can severely influence the structure of boar sperm. 5 Their results demonstrated that the number of abnormal sperm and the degree of damage gradually increased when sperm were exposed to cold shock. Moreover, the earliest signs of injury appeared in the plasma membrane, which became broken or lost. 5 These investigations further proved that the sperm plasma membrane is extremely sensitive to cryoinjury.
In addition, the freezing and thawing process greatly injured the ultrastructure of sperm acrosomes. It is well known that the structural integrity of the acrosome is critical for sperm normal fertilization with oocytes.10,23 As indicated in Figure 1C and D, in some post-thaw sperm, the outer acrosome membrane was ruptured or lost, suggesting its weak tolerance to cryoinjuries. Most severely, the inner contents in acrosomes were also partly or fully leaked (Fig. 3A). These structural alterations may directly lead to lower fertility after AI or in vitro fertilization.
Further, the main functional role of sperm is to carry male genetic materials to fertilize oocytes. Therefore, the integrity of the nucleus is especially essential to maintain the normal fertility of a sperm cell. In this study, some abnormal phenomena including deformation and partial leakage of nucleus in a few of frozen-thawed sperm were observed. The leakage from nuclei may imply loss of genetic materials. However, most frozen-thawed sperm still possess an intact nucleus, suggesting that the negative effects of the freezing and thawing process on sperm nuclei are less than that on the plasma membrane or acrosome.
Based on some previous investigations, the freezing and thawing process induced a significant increase in sperm DNA fragmentation,24,34,35 which may directly influence fertilization and subsequently embryonic development. However, the potential interrelation between the ultrastructural alterations of the nucleus and increase in post-thaw DNA fragmentation still needs further elucidation.
According to the investigation carried out by Ozkavukcu et al., the plasma membrane of the sperm head may be more sensitive to cryoinjury than the tail membrane. 26 The reason remains largely unknown. Based on this study, the plasma membranes in a few sperm were also detached from the midpiece and principal piece of the tail after the freezing and thawing process. As a primary metabolic organelle, mitochondria play critical roles in the maintenance of sperm motility and fertile capability. 36
According to the report of Aisen et al., the freezing and thawing process cannot negatively influence the mitochondrial structure of ram sperm. 6 On the contrary, in this study, although the mitochondrial sheath, outer dense fibers, and axonemes remained intact in most frozen-thawed sperm, the plasma membrane of some sperm became detached from the underlying mitochondrial helix. More seriously, the mitochondria structures in some sperm were completely disintegrated after cryopreservation. However, it cannot be determined whether the cryoinjury of mitochondrial ultrastructure is caused by the freezing and thawing procedure or the chemical toxicity of cryoprotectants.
In accordance with the report of McClean et al., the presence of glycerol or DMSO can severely damage mitochondrial ultrastructure and reduce the percentage with normal axonemes in kangaroo sperm. Moreover, DMSO can mitigate toxic injury caused by glycerol on kangaroo sperm. 33 In addition, the freezing and thawing process can severely injure the mitochondrial function.21,33 Combined with these studies, we can speculate that the post-thaw compromised function of sperm mitochondria may tightly correlate with cryoinjuries on their ultrastructure.
In the present study, the TEM results further supported the viewpoint that trehalose or 1, 3-CHD can protect the acrosome integrity of frozen-thawed ram sperm. Our results were similar to the conclusion drawn by Aisen et al. 6 In their study, trehalose was found to provide better cryoprotection for the sperm plasma membrane after observation by TEM. In addition, trehalose significantly reduced the percentage of sperm with the damaged plasma membrane. 6 However, until now, there are no reports associated with the effects of SIBs on the ultrastructure of frozen-thawed sperm.
This study first verified the cryoprotective effects of SIBs on sperm acrosome. However, different from trehalose, 1, 3-CHD cannot efficiently protect the plasma membrane integrity of ram sperm. The TEM results indicated that 1, 3-CHD cannot alleviate the negative effects of the freezing and thawing process on plasma membrane of ram sperm. However, as their natural counterparts, antifreeze proteins may provide certain cryoprotections for the sperm plasma membrane through direct interaction with the plasma membrane during the freezing and thawing process, according to some previous investigations.37,38
Different from antifreeze proteins, SIBs may only modify the shape of extracellular ice crystals and may not interact with the sperm plasma membrane, which may explain the difference of post-thaw membrane integrity between antifreeze protein and SIBs. In comparison with 1, 3-CHD, trehalose can efficiently protect the plasma membrane of ram sperm during the freezing and thawing process. As indicated in Figure 2, when trehalose was presented in the freezing extender, in most frozen-thawed sperm, their plasma membranes remained intact and tightly attached to their heads.
To quantitatively analyze the effects of trehalose or 1, 3-CHD on the post-thaw ultrastructure of ram sperm, the frequency of plasma membrane or acrosome ultrastructural alterations after cryopreservation was also assessed in this study. Our data further proved that trehalose or 1, 3-CHD can provide cryoprotection for sperm acrosomes. However, as compared with trehalose, 1, 3-CHD cannot efficiently maintain the membrane integrity of frozen-thawed sperm. This study was similar to the investigation carried out by Aisen et al. 6 In their study, when ram sperm were frozen in the presence of trehalose, only subtle detachments and swelling of the plasma membranes were observed. The heads of most of the post-thaw sperm remained intact, particularly at the apical ridge. 6
Conclusion
Based on the ultrastructural observation by TEM, this study systematically evaluated and compared the effects of trehalose or cyclohexanediol on frozen-thawed ram sperm. The TEM results indicated extensive and severe injuries on the main organelles of ram sperm during freezing and thawing. Based on the present study, the plasma membrane is most sensitive to the freezing and thawing process, followed by acrosomes and mitochondria.
The tolerance of nuclei to cryoinjuries is superior to those of the other three organelles. To a certain degree, trehalose or SIB can reduce the negative effects of the freezing and thawing process on the ultrastructure of ram sperm. The inclusion of trehalose or 1, 3-CHD in the freezing extender can provide certain cryoprotection for sperm acrosomes. However, the capability of trehalose to protect the plasma membrane integrity of frozen-thawed sperm is better than that of 1, 3-CHD.
Footnotes
Author Disclosure Statement
The authors declared that they have no conflicts of interest to this work.
Funding Information
This investigation was funded by the National Natural Science Foundation of China (Grant No. 31960663), the National Wool Caprine Industrial Technology System (Grant No. CARS-39), Yunnan Applied Basic Research Projects (Grant No. 202001AS070001), Yunnan Young Academic Leaders Program (Grant No. 202005AC160004), and Major Science and Technology Project of Yunnan Province (Grant No. 202102AE090039).