
Abstract
Aim:
This work examined the influence of induced changes in prolactin (PRL) secretion on sperm cryoresistance of ibex and the mouflon.
Materials and Methods:
PRL secretion was modified in a first experiment by the use of bromocriptine (BCR, dopamine agonist) during the non-breeding season, and in a second experiment by the use of sulpiride (SLP, dopamine D2-receptor antagonist) during the rutting season. Slow and ultra-rapid freezing protocols were used to cryopreserve sperm samples.
Results:
BCR decreased blood plasma PRL concentrations, whereas SLP increased them. Cryoresistance ratios (CRs) for curvilinear velocity (VCL), straight-line velocity (VSL), and average path velocity (VAP) in BCR-treated mouflons were lower than in controls using slow-freezing (p < 0.05), while CRs of motility and morphologically normal sperm of BCR-treated mouflons were greater than controls with ultra-rapid freezing (p < 0.05). BCR increased the head sperm dimensions in ibexes (p < 0.001); conversely, BCR decreased the head dimensions in mouflons (p < 0.001). CR-motility, CR-amplitude of lateral head displacement (ALH), CR-viability, and CR-acrosome integrity in SLP-treated mouflons were lower than in controls with slow-freezing (p < 0.01); CR-viability and CR-acrosome were lower than controls with ultra-rapid freezing (p < 0.05). In ibexes, CR-ALH was lower for SLP-treated (p < 0.05). SLP treatment increased head dimensions in ibexes (p < 0.001) but did not affect the sperm head of mouflons.
Conclusion:
Our findings show that high levels of blood plasma PRL negatively affect the cryoresistance of ibex and mouflon sperm.
Introduction
The use of artificial insemination with cryopreserved sperm allows the genetic exchange between isolated populations of wild animals that are more likely to have inbreeding risks. Nevertheless, the number of studies to improve sperm cryopreservation in wild species is scarce compared to domestic species, 1 and the fertility results are highly variable.2,3 Optimum cryoresistance is dependent not only on semen extenders 4 but also on the cooling rates used. 5 In this sense, ultra-rapid freezing is a simple and inexpensive alternative that can be used in field conditions to preserve the sperm of certain wild species. 6 Seasonal endocrine-related changes in the testicular germinal epithelium, and subsequently in sperm morphometric characteristics, can occur in a relatively short period. 7 These changes can modify the sperm, thus influencing the sperm cryoresistance since water and electrolyte membrane transport during the cooling and freezing process is affected by cell size. 8
Prolactin (PRL) acts synergistically with testosterone to maintain specific aspects of the functions and secretory activity of male accessory sex glands. 9 In wild ruminants, it is secreted with a strong seasonal pattern, following a trend roughly parallel to a photoperiod.9–11 Thus, those changes are probably related to seasonal variations of sperm cryoresistance. Indeed, in vitro supplementation with PRL affects the acrosome integrity of sperm from rams and bucks. 12 In addition, the PRL receptor (PRL-R) is expressed in pachytene spermatocytes, spermatids, and Leydig cells,13,14 suggesting a putative role of PRL during spermatogenesis.
The aim of the present work was to manipulate PRL secretion at different times of the year to determine its influence on sperm head dimensions and cryopreservation in ibex and mouflon.
Materials and Methods
Experimental design
The secretion of PRL was modified experimentally by administrating dopaminergic agonists and antagonists. Considering sensitivity to PRL concentration-associated sperm damage could be different depending on the cooling rate used, sperm samples were cryopreserved with two procedures, a conventional slow freezing and an ultra-rapid freezing protocol. All treatments and diluents were prepared in the laboratory using reagent-grade chemicals purchased from Sigma-Aldrich Inc.
In a first experiment with bromocriptine (BCR) treatment (a dopamine agonist that inhibits the release of the PRL from the pituitary gland), seven ibexes and nine mouflons were allocated to two groups: (1) Control group: 1 mL of a 0.9% NaCl solution containing 60 mg of Dextran 70 was administered i.m. twice weekly to three animals of each species, from May 15th to June 15th 2016; (2) BCR group: 10 mg of BCR in the same Dextran 70-containing vehicle were administered i.m. twice weekly to four ibexes and six mouflons from May 15th to June 15th 2016.
In a second experiment with sulpiride (SLP) treatment (a specific dopamine D2-receptor antagonist that elicits an increase in PRL secretions), 8 ibexes and 12 mouflons were allocated to two groups: (1) Control group: 2 mL of 0.9% NaCl solution containing 30 mg of tartaric acid (vehicle) were daily administered to three ibexes and six mouflons s.c from December 15th 2016 to January 15th 2017; (2) SLP group: 100 mg of s.c SLP were daily administered to five ibexes and six mouflons in the same vehicle from December 15th 2016 to January 15th 2017.
For both experiments, samples collection (7 per animal) was taken with an interval between successive collections of 15 days; one before treatment administration, two during the treatment month, and four during post-treatment period.
Animals, ultrasound analysis, and collection of samples
All experimental procedures were approved by the INIA Ethics Committee (reference number PROEX 271/14). Animals were housed in captivity at the INIA Department of Animal Reproduction. The animals were anesthetized, and the testes and sexual accessory glands were examined using real-time transrectal ultrasonography, and semen (7 samples per animal, one before treatment administration, two during treatment month, and four post-treatment) was collected using the TUMASG method as previously described. 15 Blood samples were collected by jugular venipuncture.
Sperm quality assessment
Sperm concentration was calculated before freezing using a Neubauer chamber. Sperm motility was determined using a computer-aided sperm analysis system, 15 employing Sperm Class Analyzer v.4.0. software (Microptic S.L., Barcelona, Spain). Membrane integrity and acrosomal status were analyzed with fluorescence microscopy using a fluorochrome combination of propidium iodide and fluorescein isothiocyanate-conjugated peanut agglutinin. 16 The incidence of sperm with morphological abnormalities was determined by phase-contrast microscopic examination of glutaraldehyde-fixed samples according to Esteso et al. 17 Sperm head dimensions were analyzed in fresh samples before the treatments and 50 days after treatments.
Cryopreservation procedures
The semen samples were separated into two aliquots and diluted 1:1 (vol/vol) and washed twice by centrifugation at 900 g for 20 min to retrieve the seminal plasma. The sperm was resuspended at room temperature with one of two experimental extenders until reaching a final concentration of 100 × 106 sperm/mL. The extender used for the conventional slow freezing samples was TCG for ibex and TEST for mouflon. 3 The extenders for the ultra-rapid freezing samples were the same, with the glycerol substituted by 100 mM sucrose. Sperm samples were frozen (conventional slow freezing and ultra-rapid freezing) and thawed as previously described. 5 The freezability was determined by calculating the cryoresistance ratio (CR). 6
PRL analysis
PRL plasma concentrations were determined in both seminal and blood plasma by radioimmunoassay. 18 The technique sensitivity was 1.6 ng/mL and the intra-assay CV was 5% (n = 10) and the inter-assay CV was 8% (n = 10).
Statistical analysis
Data are expressed as means ± SEM. Normal distribution of the data was determined with the Shapiro–Wilk test, and non-normally distributed data were log-transformed, arcsine-transformed (percentages), or arcsine-transformed (in case of variables including zero values) before analysis. The effects of treatments and their interaction with the data period on sperm variables and the CR values returned by each freezing procedure were compared with GLM repeated measures ANOVA. All statistical analyses were carried out using Statistica for Windows v.13.3 software (Tibco, Inc., Tulsa, OK).
Results
The influence of BCR treatment on sperm head dimensions and sperm cryoresistance was analyzed in the first experiment. The treatment with this dopamine agonist affected PRL secretion in both species. Specifically, BCR decreased blood plasma PRL concentrations in both ibexes (p < 0.001; 39.4 ± 14.5 ng/mL vs. 119.0 ± 26.0 ng/mL and 22.0 ± 4.5 ng/mL vs.134.0 ± 10.9 ng/mL, in May and June, respectively) and mouflons (54.2 ± 14.8 ng/mL vs. 205.0 ± 52.1 ng/mL and 11.0 ± 2.1 ng/mL vs. 220.0 ± 51.0 ng/mL, in May and June, respectively). However, treatment with BCR did not affect seminal plasma PRL concentrations in any species. Regarding the ultrasound analysis made 2 months after treatment, data showed that the testicular diameter of BCR-treated mouflons was lower than untreated males (p < 0.05; 3.4 ± 0.1 cm vs. 3.8 ± 0.2 cm and p < 0.01; 3.8 ± 0.1 cm vs. 4.5 ± 0.1 cm, in July and August, respectively).
The size of seminal vesicles and bulbourethral glands were not affected by treatment with BCR in either ibexes or mouflons. Sperm analysis of fresh semen revealed that BCR-treated mouflons and ibexes had a greater percentage of sperm with morphological abnormalities than controls (p < 0.05; 52.4% ± 6.6% vs. 24.9 ± 7.3% in mouflons; p < 0.01, 92.0% ± 2.8% vs. 64.8% ± 7.0% in ibexes). During the post-treatment period, the percentage of sperm with morphological abnormalities was greater in ejaculates from mouflons treated with BCR than control mouflons (p < 0.01; 61.8% ± 6.4% vs. 29.6% ± 6.9%). During the post-treatment period, the semen collected from BCR-treated mouflons had lower CRs for curvilinear velocity (CR-VCL), straight-line velocity (CR-VSL), and average path velocity (CR-VAP) than controls (p < 0.05; Fig. 1) after slow freezing.
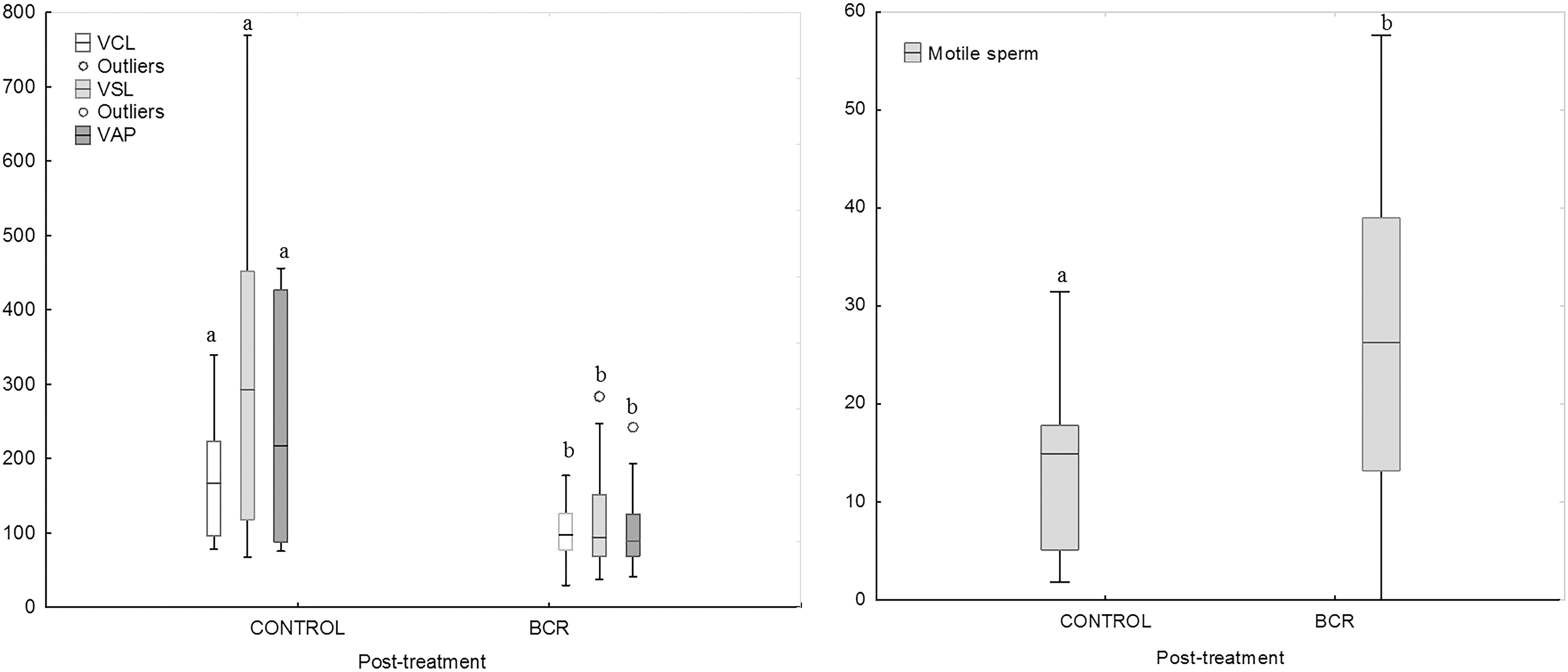
Cryoresistance ratios in control and BCR-treated mouflons, after slow-freezing (left) and ultra-rapid freezing (right). Different letters indicate differences (p < 0.05) between the control and treatment group. BCR, bromocriptine; VCL, curvilinear velocity; VSL, straight-line velocity; VAP, average path velocity.
On the other hand, the CR-motility and CR for morphological normal sperm were greater in semen collected from BCR-treated than from untreated mouflons after ultra-rapid freezing (p < 0.05) (Fig. 1). The fresh sperm cells collected from the BCR-treated ibexes had greater head width, area, and perimeter than controls (p < 0.001 for all the variables). Conversely, the sperm cells collected from BCR-treated mouflons had a lower head width, area, and perimeter than untreated during the post-treatment period (p < 0.001 for all the variables) (Supplementary Table S1).
The influence of SLP treatment on sperm head dimensions and sperm cryoresistance was analyzed in the second experiment. The treatment with this dopamine D2-receptor antagonist increased blood plasma PRL levels, but basal levels were reached by 24 hours post-administration in both ibexes and mouflons. However, there were no differences in seminal plasma PRL concentration between SLP-treated and controls in both species. The ultrasound evaluation made 2 weeks after the treatment revealed that testicular diameter was lower in SLP-treated than control ibexes (p < 0.05; 2.3 ± 0.1 cm vs. 2.8 ± 0.1 cm respectively), while seminal vesicles were greater in SLP-treated than control mouflons (p < 0.001; 0.4 ± 0.0 cm2 vs. 0.3 ± 0.0 cm2). Sperm analysis of fresh semen showed that VCL and amplitude of lateral head displacement (ALH) were greater in samples collected from SLP-treated than from control mouflons during the treatment period (p < 0.01; 77.8 ± 4.0 vs. 56.7 ± 5.5 for VCL and 2.9 ± 0.2 vs. 2.1 ± 0.2 for ALH, respectively).
There were no differences for motility and kinetic variables in fresh semen collected in ibexes. During the treatment period, CR for normal sperm morphology was lower in samples collected from SLP-treated than from untreated ibexes with both procedures, when semen was slow and ultra-rapidly frozen (p < 0.05) (Fig. 2). CR-motility, CR-ALH, CR-viability, and CR-acrosome integrity in semen collected from SLP-treated mouflons were lower than in semen collected from controls when semen was slow-frozen (p < 0.01 for all) (Fig. 3); CR-viability and CR-acrosome were lower than in controls when semen was ultra-rapidly frozen (p < 0.05 for all) (Fig. 3).

Cryoresistance ratios in control and SLP-treated ibexes, after slow-freezing (left) and ultra-rapid freezing (right). Different letters indicate differences (p < 0.05) between the control and treatment group. ALH, amplitude of lateral head displacement.
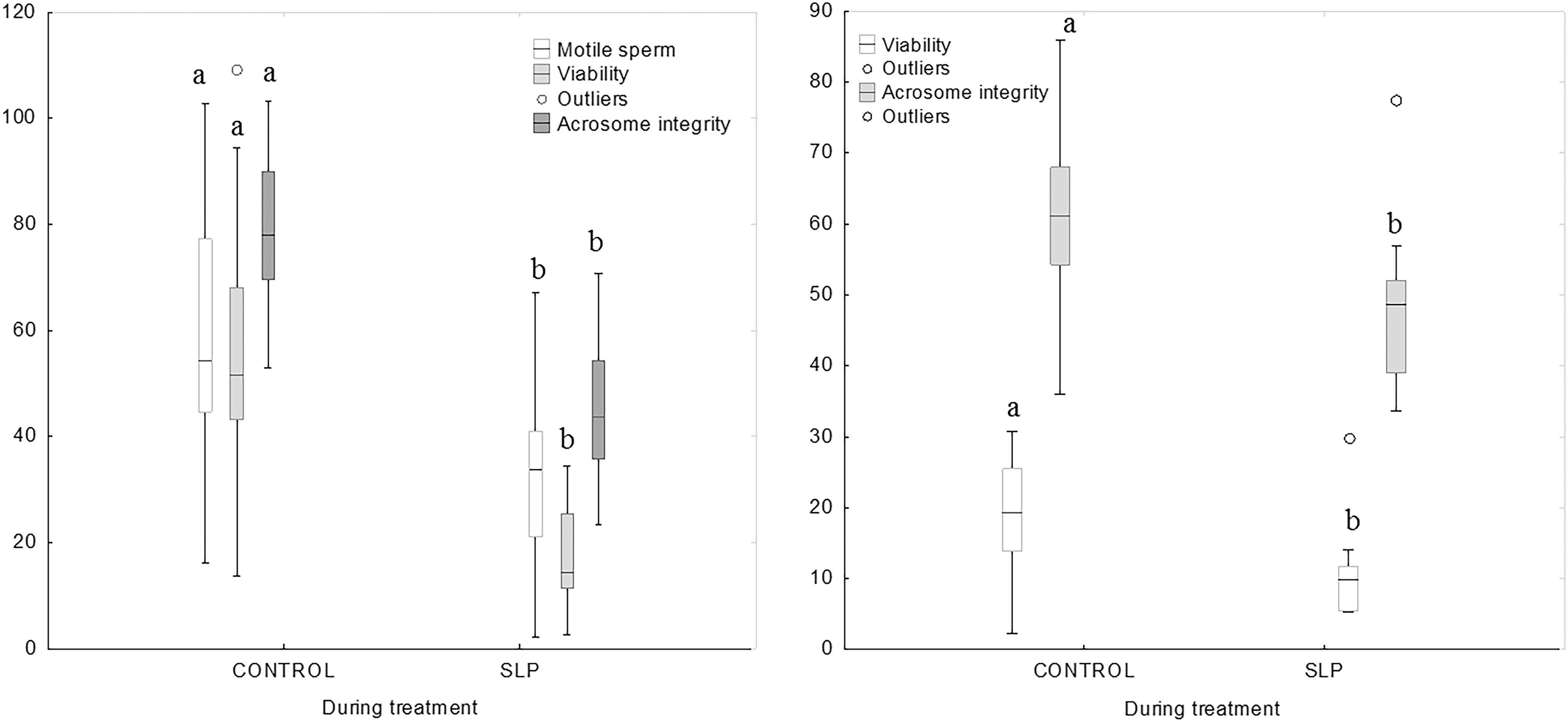
Cryoresistance ratios in control and SLP-treated mouflons, after slow-freezing (left) and ultra-rapid freezing (right). Different letters indicate differences (p < 0.05) between the control and treatment group.
During the post-treatment period, CR-ALH and CR-acrosome integrity were lower in semen collected from SLP-treated than from untreated mouflons when sperm was slow-frozen (p < 0.05 for both) (Fig. 4). CR-ALH was lower in semen collected from SLP-treated than untreated ibexes (p < 0.05) (Fig. 2). SLP-treated ibexes showed greater head dimensions than controls (p < 0.001). The treatment with SLP did not affect sperm head dimensions in sperm collected from mouflons (Supplementary Table S2).
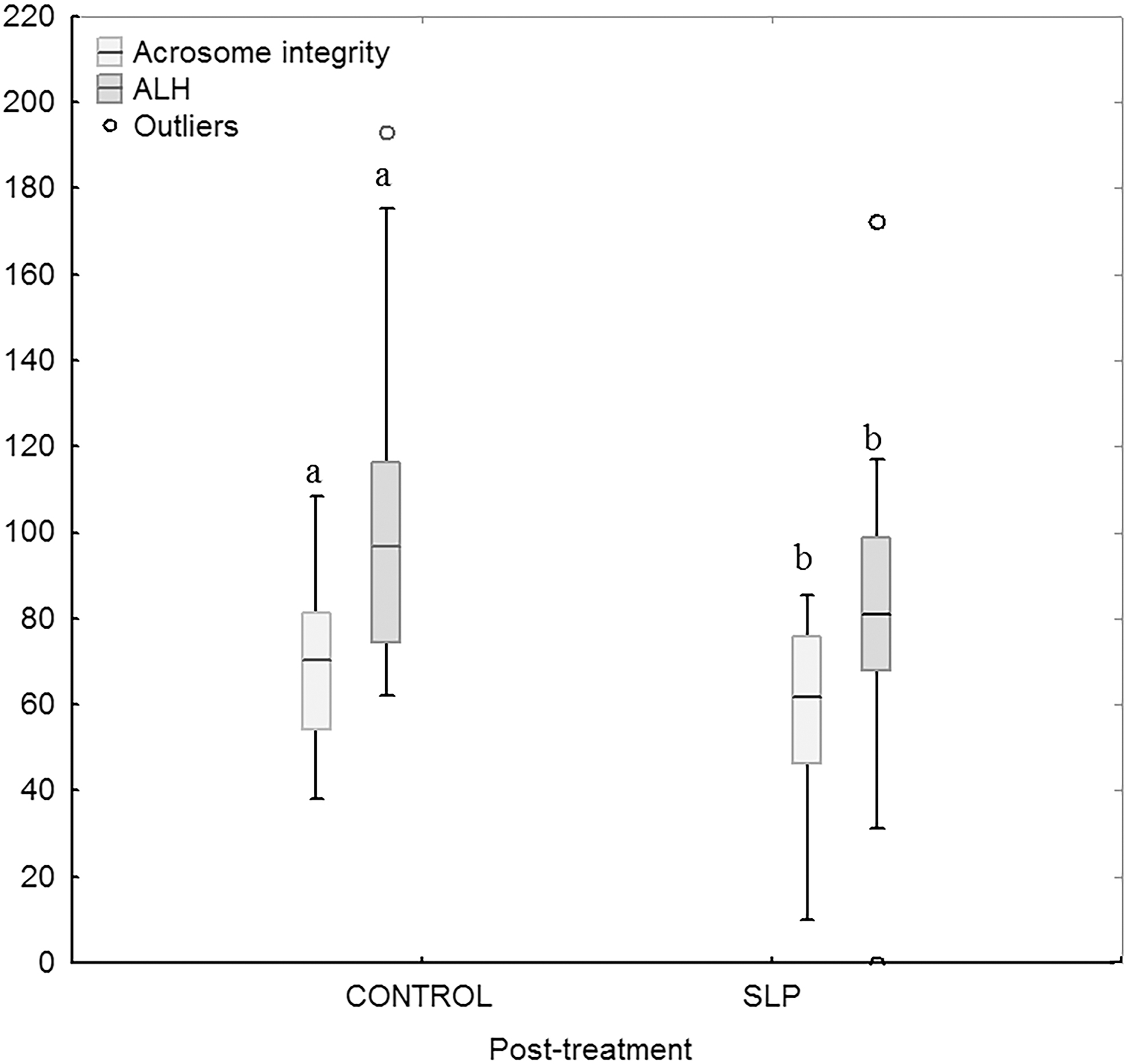
Cryoresistance ratios in control and SLP-treated mouflons after slow-freezing. Different letters indicate differences (p < 0.05) between the control and treatment group.
Discussion
This is the first study in wild ruminants demonstrating that PRL plasma concentrations are directly related to sperm cryopreservation. The sperm cryoresistance was associated with decreases or increases in blood plasma PRL concentrations provoked by treatments with BCR and SLP, as described by Toledano et al. 11 and Lincoln et al. 19 The effect of endocrine changes (e.g., testosterone variations) on sperm freezability by an indirect influence during the spermatogenic cycle has been previously reported in mouflons, 7 but these results expand the information to the influence of PRL blood concentration. In general, BCR treatment produced a variable effect on sperm cryoresistance (decreasing or increasing sperm motility variables), but SLP treatment–which increased blood plasma PRL concentrations–induced a negative influence on sperm freezability.
To the best of our knowledge this is the first study demonstrating that blood PRL concentrations are directly related to sperm freezability in wild ruminants. However, apparently this effect is not related to PRL concentrations in seminal plasma, as these values did not differ even when differences in blood PRL concentrations were higher.
In fresh samples, there were more morphological abnormalities when blood plasma PRL secretion decreased as a consequence of the treatment with BCR. In agreement, the treatment with SLP, and thus, increased blood levels of PRL enhanced certain kinetics variables, such as VCL and ALH, and the percentage of sperm with normal morphology. PRL appears to act directly and also synergistically with androgens, to maintain the activity of male accessory sex glands.20,21 In this sense, the endocrine balance between hormones produced in the pituitary and the testis affects spermatogenesis and spermiogenesis processes. 22 PRL increases the sensitivity of the Leydig cells to LH to restore spermatogenesis in rats, 23 and has a direct effect on sperm cell metabolism, motility, and fertilizing capacity. 24 Therefore, the present results expand this relationship to sperm cryoresistance in wild ruminants.
The influence of SLP, and thus, higher blood plasma PRL concentrations, on cryoresistance during the period of treatment in mouflons and ibex suggests an effect at the epididymal level, where PRL is involved in sperm maturation. 25 Although ultra-rapid cooling rates usually produce stronger ultrastructural damages on sperm cells than conventional slow cooling rates, 5 our finding revealed that decreasing the CR after SLP treatment was variable for different sperm parameters when slow or ultra-rapid freezing was used. However, effects of elevated PRL levels are not only at the epididymal level, as during the post-treatment period CR for ALH and acrosome were also affected, suggesting that there were negative effects of high PRL concentrations through the spermatogenic cycle in both species.
Our data confirm that despite high levels of PRL in blood plasma possibly being associated with an appropriate sperm function, it negatively affects the sperm CR. 26 The decreasing of CR-acrosome integrity after SLP treatment is in agreement with previous findings in rams and bucks, in which in vitro incubation with PRL decreased the post-thaw acrosome integrity. 12
Unlike SLP treatment, the BCR treatment returned a variable effect on sperm freezability, for instance decreasing the CR for kinetic variables (VCL, VSL, and VAP) and improving the CR for total motile sperm. The treatment with BCR affected blood PRL secretion, but it might also affect the dopaminergic signaling in capacitation and sperm motility. Indeed, dopamine type 2 receptors are present in a wide range of mammalian sperm.27,28 Previous reports have shown that BCR and low-concentration dopamine increased total and progressive motility of boar sperm; conversely, BCR and high dopamine levels increased tyrosine phosphorylation during the capacitation period. 28 During sperm capacitation, the sperm cell suffers some modification on permeability and fluidity of the membrane, and ions transport, and thus cryo-capacitation might explain the decrease of CR for certain sperm variables, according to the cooling rate applied. On the other hand, BCR has direct negative effects on the motility and some kinetic variables of human sperm. 29
Our findings showed that both treatments affected the sperm head dimensions in fresh samples ∼50 days post-treatment, demonstrating that the treatments had effects for a long time after they ended. BCR treatment increased the head sperm size in ibexes; conversely, BCR reduced the head sperm size in mouflons, as has been reported in domestic rams. 26 Variations in sperm head size might affect the sperm water volume, and the membrane permeability to water and cryoprotectants, and the sperm response to the freeze-thawing process. 30 The decrease in sperm head sizes could explain the better CR-motility after ultra-rapid freezing. In turn, SLP treatment increased the sperm head size in both species, which may be also associated to a decreased cryotolerance.
In some species, PRL may play a physiological role in the growth of the testes and sex accessories.15,31,32 This effect is consistent with the increase of seminal vesicles size after SLP treatment of mouflons and the decrease of testicular size after BCR treatment. Variations in the dimensions of these glands are accompanied by changes in their functional activity that in turn might affect sperm and their tolerance to cryopreservation process.
In conclusion, our findings showed that PRL has an important role in sperm cryoresistance in wild ruminants, and thus seasonal variations in sperm freezability appear to be associated with seasonal changes in PRL secretion. High levels of blood plasma PRL negatively affect the cryoresistance for many sperm variables, but in a species-specific way. This study contains relevant information that could be applied in futures works related with the use of PRL like biomarkers of cryopreservation efficacy.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The study reported here was funded by MINECO/AEI/FEDER and EU grant AGL2017-85753-R. P. Bóveda was the recipient of a grant for pre-doctoral researchers from MINECO (AEI/FSE, UE). Octavio Mejía was the recipient of a research fellowship from the PASPA-DGAPA-UNAM (México).
This work is based in part on research presented in Dr. Bóveda's doctoral thesis (available at https://repositorio.uam.es/bitstream/handle/10486/699401/boveda_gomez_paula.pdf?isAllowed=y&sequence=1).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.