
Abstract
Highlights
Using cysteine and purslane extracts in extenders improved significantly the post-thaw sperm characteristics.
Sperm viability, DNA integrity, and mitochondrial activity demonstrate an improvement in post-thaw sperm.
Malondialdehyde production was decreased based on the positive effects of treated extenders.
The obtained results demonstrate that supplementation of 50 μg/mL of purslane methanolic extract with cysteine to freezing extenders was significantly superior compared with other treatments.
This study determined the effects of purslane aqueous extract (PAE), purslane methanolic extract (PME), and purslane ethanolic extract (PEE) compared with cysteine on goat semen. The samples collected for evaluation were pooled and diluted with a basic Tris diluent 50 μg/mL of purslane aqueous, methanolic, or ethanolic extract alone and in combination with 10 mM cysteine. Results demonstrated significant improvements using treated diluents in total motility, viability, mitochondrial activity, and decreased malondialdehyde (MDA) level. Also, all of the combined treatments and PME50μg/mL resulted in higher progressive motility, live spermatozoa, and integrity of DNA and plasma membranes. The apoptotic and dead spermatozoa were decreased significantly using PME50μg/mL, cysteine10mM + PME50μg/mL, and cysteine10mM + PAE50μg/mL. DNA integrity in cysteine10mM + PME50μg/mL was demonstrated to be higher than cysteine and PEE50μg/mL. Moreover, higher viability was observed in cysteine10mM + PME50μg/mL compared with the PME50μg/mL, PAE50μg/mL, PEE50μg/mL, and cysteine. From comparison among all treatments, cysteine10mM + PME50μg/mL showed the highest percentages of total motility, mitochondrial activity, and the lowest percentages of MDA. In conclusion, inclusion of purslane extracts and cysteine into the extenders improved the quality of cryopreserved spermatozoa. However, the mixture of methanolic extract and cysteine was more beneficial among the various treatments.
Introduction
The quality of frozen semen is one of the important factors influencing successful fertilization and pregnancy rates. 1 Since about 50% of sperms are damaged during the cryopreservation processes, quality and, as a result, fertility of frozen sperms are weaker than those of fresh ones. 2 High concentrations of reactive oxygen species (ROS) play a significant role in sperm damage pathophysiology. 3 The main source of increased generation of ROS is cooling and freezing; thawing processes also reduce the antioxidant content in semen to such an extent that sperms experience oxidative stresses. 4 These factors induce apoptosis in sperms, DNA damage, and loss of sperm functions in semen samples following the thawing processes.1,5
Under normal conditions, concentrations of free radicals are controlled by enzymatic (such as CAT, SOD, and GSH-Px) and nonenzymatic (such as glutathione, urate, pyruvate, vitamin E, and vitamin C) antioxidants in semen and sperm cells. 6 However, some imbalance is created between ROS generation and antioxidative defense mechanisms due to an increase in ROS production during the cryopreservation processes. Due to the suppression of antioxidant systems, sperms undergo some damages including lipid peroxidation in cellular and mitochondrial membranes, which, in turn, results in changes in membrane permeability and destruction of membrane-cell integrity and which, eventually, mitigates mobility, membrane intactness, and fertilization ability of sperms.7–10 Such dysfunctions are caused by high amounts of unsaturated fatty acids in the cell membranes of mammalian sperms, increasing sperm sensitivity to lipid peroxidation. 2
Antioxidants are capable of preventing ROS-induced oxidation of biologically important molecules by interfering with oxidation processes in cell membranes, chelating catalytic metals, and by scavenging free radicals. 11
Cysteine is a thiol amino acid with a low molecular weight, easily permeating into cells, and is a scavenger of glutathione molecules. 12 Cryopreservation processes reduce the glutathione content in seminal plasma and sperms. Cysteine added to extenders improves functions of and increases fertility of sperms by stimulating intracellular glutathione synthesis and protecting the DNA, proteins, and lipids of cell membranes during the cryopreservation processes.13,14 Recently, research done into different species has reported that using cysteine as an antioxidant has beneficial effects on the protection of sperms against cryodamages in various species such as ram.15–22
It is known that a lot of medicinal herbs contain considerable amounts of antioxidant compounds, one of which is Portulaca oleracea L. (purslane), an annual plant from Portulacaceae family. 23 All biologically active compounds of purslane, such as phenolic acids, flavonoids, alkaloids, carotenoids, melatonin, glutathione vitamins, and minerals, have an antioxidant activity.24–29 Phenolic compounds found in dietary and medicinal plants indicate the potential for the inhibition of oxidative stresses.30,31 Purslane is rich in phenolic compounds having a good antioxidant potential. 32 Purslane exhibits its antioxidant activity by scavenging ROS and inhibiting lipid peroxidation, as shown in many studies.33–41 Moreover, the evidence indicates that administration of purslane extract to rats resulted in the reduction of MDA and nitric oxide in the testes, liver, and kidneys. 42
Cai et al. 43 and Simopoulos 44 reported that flavonoids (quercetin), omega-3, β-carotene, and glutathione found in purslane had antioxidative activities and stated that purslane might inhibit lipid peroxidation by scavenging free radicals and increasing intracellular glutathione concentrations. In addition, in a previous study we examined the antioxidant effects of different levels of aqueous, methanolic, and ethanolic extracts of purslane on frozen sperm and found that purslane extracts at levels of 50 μg/mL improved goat sperm quality after freezing. The best effect was related to the methanolic extract. 45
Based on the above research, it can be said that as antioxidants, cysteine and extracts of purslane play important roles in scavenging free radicals. Recent studies showed the effectiveness of other antioxidants such as cysteine, trehalose, and hypotaurine or rosemary plant and cysteine either in isolation or in combination, in preventing cryodamage from occurring in freezing extenders.21,46 Given that the effects of combined cysteine and purslane extracts have not been investigated yet, the aim of the present study is to evaluate the quality of goat sperms in extenders supplemented with purslane extracts, cysteine, or both to see if they have positive effects on the quality of goat sperms after performing cryopreservation processes.
Materials and Methods
According to the international guidelines, all performed experimental procedures have been approved by the Animal Care and Use Committee of the University of Kurdistan, Sanandaj/Iran, with the registration Id as following: student number 9315000102 at 21.11.2017.
Chemicals
All chemicals used in the experiment were obtained from Sigma-Aldrich (St. Louis, MO) and Merck (Darmstadt, Germany).
Extraction of purslane
Purslane was collected from a lot of land in the Kurdistan region, washed, air-dried, and, finally, powdered by a mill. To prepare an aqueous extract, powdered plant was extracted by distilled water (1 g/10 mL) at 100°C for 2 hours and centrifuged at 1000 × g for 20 minutes. The resulting supernatant was filtered by a filter paper and condensed by a rotary evaporator (Hei-Vap; Heidolph, Schwabach, Germany) at 40°C under decreased pressure. The concentrated extract was freeze-dried by a lyophilizer and the resulting powder was stored at 4°C. 47 To prepare ethanolic and methanolic extracts, 100 g of purslane powder was soaked in 800 mL each of ethanol or methanol for 72 hours. This mixture was filtered, and then, the resulting solution was condensed by a rotary evaporator vacuum system at 40°C, dried in a 30°C oven, and, finally, stored at 4°C. 48
Animals, semen collection, and preparation
This study was conducted at a testing station located in the city of Sanandaj. To perform this experiment, semen samples were collected from four adult male Markhoz goats (3–4 years old and weight 60–70 kg) by electrical stimulation of ejaculation twice a week. Samples were then transferred to the laboratory within 10–15 minutes after collecting semen of each male. In the laboratory, samples were placed in a 37°C hot water bath and then assessed for their initial quality. As a result of this assessment, samples with motility >75% and sperm concentration >1 × 109/mL were selected for the next step of the experiment. Semen samples from four males were pooled in each iteration to exclude individual differences. All the treatments were undertaken in 10 replicates.
The used freezing extender has been composed of 3.786 g Tris, 2.172 g citric acid, 1 g fructose in 100 mL distilled water, containing 5% (v/v) glycerol and 5% (v/v) egg yolk based on the method described by Evans and Maxwell. 49 The extender was set at osmolality 320 and pH 6.8. The test treatments included a basic extender with no additives (control) and a basic extender supplemented with 10 mM cysteine (cysteine10mM).
The concentration of cysteine was determined based on the optimal concentration obtained in our previous study, 16 basic extender supplemented with 50 μg/mL of purslane methanolic extract (PME50μg/mL), basic extender supplemented with 50 μg/mL of purslane aqueous extract (PAE50μg/mL), basic extender supplemented with 50 μg/mL of purslane ethanolic extract (PEE50μg/mL), basic extender supplemented with 10 mM cysteine plus 50 μg/mL of purslane methanolic extract (cysteine10mM + PME50μg/mL), basic extender supplemented with 10 mM cysteine plus 50 μg/mL of purslane aqueous extract (cysteine10mM + PAE50μg/mL), and basic extender supplemented with 10 mM cysteine plus 50 μg/mL of purslane ethanolic extract (cysteine10mM + PEE50μg/mL).
Concentrations of extracts were determined based on our preliminary study where it was observed that supplementation of goat semen cryopreservation extenders with 50 μg/mL of any of purslane methanolic, aqueous, and ethanolic extracts provided better quality of thawed semen compared with other concentrations. 45 Semen with a concentration of 240 × 106 spermatozoa/mL was diluted in the extenders. Then, diluted samples were loaded into straws (0.25 mL). Once the open ends of straws were blocked by polyvinyl chloride powder, in the cooling step, their temperatures reached 5°C for 3 hours. In the freezing step, straws held 4 cm above the liquid nitrogen level were frozen in nitrogen vapor for 15 minutes. Then, they were immersed in a container containing −196°C liquid nitrogen and stored. In the thawing step, frozen straws were thawed in a 37°C water bath for 30 seconds and used for different assessments.
Evaluation of sperm motion parameters
A computer-assisted sperm motility analysis system (CASA: IVOS version 12; Hamilton-Thorne Biosciences) was used for evaluation of sperm motion characteristics. For the assessment of the post-thaw sperm characteristics, the following parameters were evaluated: total motility (%), progressive motility (%), VAP (average path velocity, μm/s), VSL (straight-line velocity, μm/s), VCL (curvilinear velocity, μm/s), ALH (amplitude of lateral head displacement, μm), LIN (linearity [LIN% = VSL/VCL]), and STR (straightness [STR % = VSL/VAP]). CASA was set up as follows: phase contrast; frame rate—60 Hz; minimum contrast—70; low and high static size gates—0.6–4.32; low- and high-intensity gates—0.20–1.92; low- and high-elongation gates 7–91; default cell size—10 pixels; and default cell intensity—80. Straws were removed from liquid nitrogen, and 5 μL of samples was put onto a prewarmed slide (37°C) and covered with a coverslip.
Sperm motility characteristics were determined with a 10 × objective at 37°C. The CASA software captured 25 sperms from 10 fields. The images were analyzed using a phase-contrast microscope at × 1000 magnification.
Evaluations of the integrity of plasma membrane and acrosome
A hypo-osmotic test was used to assess the membrane functional integrity of sperm according to Revell and Mrode. 50 This technique involved incubation of 20 μL of diluted sperms with 200 μL of hypo-osmotic solution (9 g fructose +4.9 g sodium citrate/1 L distilled water) at 37°C for 60 minutes. After incubation, one drop of solution was placed onto a preheated slide and covered by a coverslip. Then, a total of 200 sperms in at least 5 different microscopic fields (400 × magnification) were counted and the percentage of those with swollen or coiled tails, Hypo-osmotic-Test (HOST) positive, was determined.
The morphological abnormalities in the acrosome section from frozen–thawed sperm cells were assessed according to Daramola and Adekunle, 51 by mixing sperms with citrate formalin solution (96 mL of 2.9% sodium citrate, with 4 mL of 37% formaldehyde), placing a drop of fixed sperms onto a slide and observing them by an optical microscope (1000 × magnification). A total of 200 sperms were counted and the percentage of those with intact acrosomes was determined.
Evaluation of sperm viability
Viability of sperms was assessed by using eosin and nigrosin dyes. 49 Five microliters of diluted sperms was placed onto a slide and mixed with 10 μL of eosin–nigrosin solution. Stained sperms were spread on the slide. A total of 400 sperms under an optical microscope (400 × magnification) were counted for viable (white) and non viable (red) cells.
Lipid peroxidation
As an indication of lipid peroxidation, 52 MDA was measured by the thiobarbituric acid (TBA) reactive substances test. The thawed sperm samples were centrifuged at 1500 × g for 5 minutes and 1 mL of resulting supernatant was mixed with 1 mL of EDTA (0.037 g EDTA in 10 mL distilled water), 1 mL of BHT (0.2 g BHT in 10 mL ethanol), and 2 mL of TCA (3 g TCA in 30 mL distilled water) and centrifuged at 1200 × g for 15 minutes. Subsequently, 1 mL of the resulting supernatant was incubated with 1 mL of TBA (0.134 g TBA in 20 mL distilled water) in a 90°C water bath for 20 minutes. Once the mixture was cooled at room temperature, its absorbance was read by a spectrophotometer at 532 nm. The MDA concentrations were expressed as nmol/mL.
Mitochondrial activity
Mitochondrial activities were assessed using the combination of rhodamine (R123; Invitrogen™, Eugene, OR), fluorescent, and PI dyes. Ten microliters of R123 solution (0.01 mg/mL water distillated) was incubated with 500 μL of sperms diluted by Tris buffer in the dark for 20 minutes. After centrifugation of samples at 1200 × g for 10 minutes, the sperm pellet was resuspended in 500 μL of Tris buffer and finally 10 μL of PI (1 mg/mL) was added. The percentage of viable spermatozoa with active functional mitochondria was identified by a positive signal for Rh123 and a negative signal for PI. For each sample, 10,000 events were considered according to Najafi et al. 53
Phosphatidylserine translocation assay
The apoptosis status of cryopreserved sperm has been evaluated using the method described by Najafi et al. 53 For that purpose, the phosphatidylserine translocation detection commercial kit was used according to the manufacturer's instructions (IQP, Groningen, the Netherlands). After washing spermatozoa with calcium buffer and adjusting the concentration of sperm to 1 × 106 spermatozoa/mL, 10 μL of Annexin V-FITC was added to a 100 μL sperm suspension, which was incubated for 15 minutes at room temperature. Then, 5 μL of PI was added and incubated for 15 minutes in the dark. The sperm samples were then analyzed in a FACSCalibur (Becton Dickinson, San Khosoz, CA) flow cytometer. The results were determined as follows: live spermatozoa (Annexin−/PI−), apoptotic spermatozoa (Annexin+/PI−), and dead spermatozoa (PI+). For each experimental group, a minimum of 10,000 sperms was examined.
Sperm chromatin dispersion test
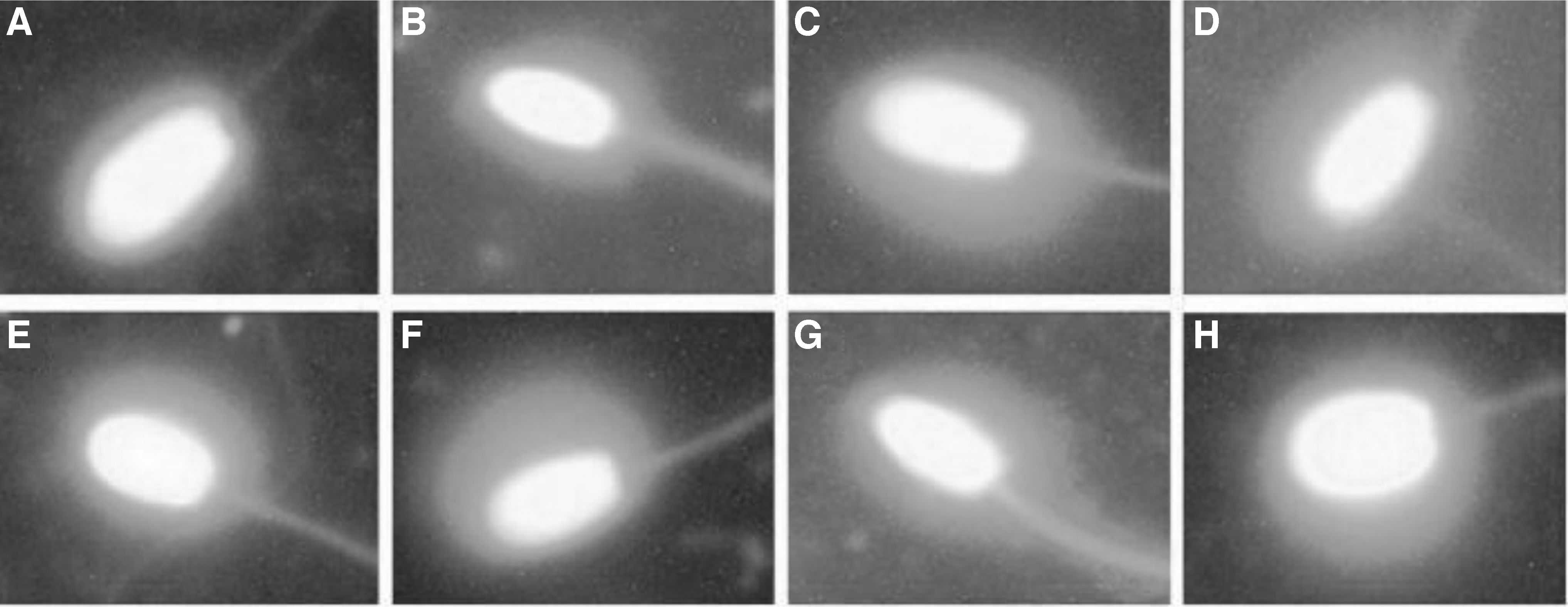
The DNA fragmentation (Fig. 1) of cryopreserved sperm were evaluated using the modified method according to Fernandez et al. 54 One hundred fifty microliters of 65% agarose was placed onto a slide, covered by a coverslip, and kept at 4°C. The coverslip was removed after 5 minutes when a solid agarose layer was formed on the slide. Next, 30 μL of sperm samples was mixed with 70 μL of 0.7% low melting point agarose, placed on the solid agarose layer and left to get drier. After removing the coverslip, the slide was put horizontally in an acid denaturing solution (0.08 N HCl) at 37°C in darkness for 7 minutes.

The DNA fragmentation using ethidium bromide, which indicates the stained sperm nucleoid containing a peripheral halo of dispersed (left arrow) and central core (right arrow) DNA loops. The halo size is being computed by dividing the HS/WNS by ImageJ (Image Processing and Analysis in Java) software. HS/WNS, halo surface/whole nucleoid surface.
Then, the slide was placed in a lysing solution (0.4 M Tris base, 0.8 M DTT, 1% SDS, 50 mM EDTA, and 2 M NaCl, pH = 7.5) at room temperature for 25 minutes. After that, the slide was removed from the solution, washed with distilled water for 5 minutes, dehydrated in 70%, 90%, and 100% ethanol, respectively, each for 2 minutes, and, finally, left to dry in the air. The slide was stained with ethidium bromide staining solution and sperms were assessed by a fluorescent microscope. The halo size of each cell was evaluated by the surface of the halo divided by the surface of the whole nucleoid.
Statistical analysis
Data were analyzed using Proc GLM of SAS (version 9.1; 2002; SAS Institute, Cary, NC), in a completely randomized design. Orthogonal contrasts were used to compare the means and statistical significance was determined as p < 0.05. The results are expressed as the mean ± standard error.
Results
The observed results from this study demonstrated that all treatments significantly enhanced (p < 0.05) the total motility, viability, and mitochondrial activity and decreased MDA levels (p < 0.05) compared with the control group. Also, the results showed that PME50μg/mL and all of the combined treatments produced the higher (p < 0.05) percentages of membrane integrity, live spermatozoa, and the halo surface/whole nucleoid surface (HS/WNS) ratio than the control (Tables 1 and 2). With regard to apoptotic and dead spermatozoa, PME50μg/mL, cysteine10mM + PME50μg/mL, and cysteine10mM + PAE50μg/mL caused significant (p < 0.05) decreases in comparison with the control group (Table 2).
Effects of PME50μg/mL, PAE50μg/mL, and PEE50μg/mL Along with Cysteine10mm on Motility Parameters, Functional Membrane Integrity, Acrosome Status, and Viability of Post-Thawed Goat Spermatozoa (Mean ± Standard Error; n = 10)
LIN % = VSL/VCL. STR % = VSL/VAP.
Different superscripts within the same row demonstrate significant differences (p > 0.05).
ALH, amplitude of lateral head displacement; cysteine10mM, 10 mM of cysteine; LIN, linearity; PAE50μg/mL, 50 μg/mL of purslane aqueous extract; PEE50μg/mL, 50 μg/mL of purslane ethanolic extract; PME50μg/mL, 50 μg/mL of purslane methanolic extract; STR, straightness; VAP, average path velocity; VCL, curvilinear velocity; VSL, straight-line velocity.
Effects of PME50μg/mL, PAE50μg/mL, and PEE50μg/mL Along with Cysteine10mM on Malondialdehyde Concentration, SCD-Stained Halo Surface/Whole Nucleoid Surface, Mitochondrial Activity, Viable, Apoptotic, and Dead Post-Thawed Goat Spermatozoa (Mean ± Standard Error; n = 10)
Different superscripts within the same row demonstrate significant differences (p > 0.05).
HS/WNS, Sperm Chromatin Dispersion (SCD), halo surface/whole nucleoid surface.
The MDA production was found to decrease significantly with the PME50μg/mL compared with cysteine, PAE50μg/mL, and PEE50μg/mL. Furthermore, PME50μg/mL produced a higher (p < 0.05) percentage of mitochondrial activity and live spermatozoa than PEE50μg/mL and cysteine (Table 2).
Total motility in cysteine10mM + PME50μg/mL and cysteine10mM + PAE50μg/mL was significantly higher (p < 0.05) compared with cysteine. Cysteine10mM + PME50μg/mL resulted in a significantly (p < 0.05) higher membrane integrity and viability rate compared with PME50μg/mL, PAE50μg/mL, PEE50μg/mL, and cysteine (Table 1). Also, the addition of cysteine10mM + PME50μg/mL provided a greater protective effect in the percentage of live spermatozoa in comparison with the PAE50μg/mL, PEE50μg/mL, and cysteine (p < 0.05).
Also, the ratio of HS/WNS was demonstrated to be higher (p < 0.05) in cysteine10mM + PME50μg/mL than that of PEE50μg/mL, and cysteine following the freeze–thaw (Table 2 and Fig. 2). Cysteine10mM + PME50μg/mL had the highest (p < 0.05) total motility and mitochondrial activity rates (Tables 1 and 2). Also, cysteine10mM + PME50μg/mL showed the lowest (p < 0.05) levels of MDA than the other treatments (Table 2).
Discussion
Data obtained in this study showed that the addition of 50 μg/mL of purslane aqueous, methanolic, and ethanolic extracts to freezing extenders improved the quality of thawed goat sperms. Our findings also showed that addition of 10 mM cysteine to diluents had antioxidative effects on the improvement of thawed goat sperm quality. These results are similar to those obtained by other researchers suggesting that cysteine had antioxidative effects in protecting sperms against cryodamage.8,16,18,20 It is not fully known what the precise mechanisms of sperm protection by amino acids are. It has been reported that the amino acids demonstrate a stabilizing effect on membranes, and amino acids limited membrane lesions, inhibited plasmolysis, and prevented membrane-based lipid peroxidation during cryopreservation processes.55,56
Based on the comparison of extracts in this study, the results indicate that the extender containing 50 μg/mL of methanolic extract exhibited higher motility and lower MDA compared with the extender containing 50 μg/mL of aqueous and ethanolic extracts, and exhibited higher mitochondrial activity and number of live sperms in comparison with the extender containing 50 μg/mL of ethanolic extract. Our previous study 45 showed that methanolic extracts contained higher amounts of phenolic compounds than aqueous and ethanolic extracts. Given the strong relationship between the antioxidative power and phenolic content of purslane, 57 it can be concluded that the higher sperm quality in the treatment containing methanolic extract is probably due to its higher amounts of phenolic compounds in comparison with aqueous and ethanolic extracts.
The antioxidative protective roles of plants can be attributed to their phenolic compounds being predominantly antioxidants with a wide range of distribution in plants and being able to scavenge free radicals directly. Purslane contains significant amounts of phenolic compounds, 37 including phenolic acids, flavonoids, and phenolic alkaloids.24,58 The scavenging capability of purslane phenolic compounds may be due to the ability of hydroxyl substitution to donate reactive hydrogen. 24
Based on results from this study, the addition of 50 μg/mL of purslane aqueous, methanolic, and ethanolic extracts to cryopreservation extenders could reduce MDA concentrations and increase sperm motility in these treatments compared with the control group by preventing lipid peroxidation. Similar to our results, the reduction of MDA levels and increased ram sperm motility after freeze–thawing processes were observed by Merati and Farshad 59 who used ginger and Echinacea extracts, and Khodaei Motlagh et al. 60 who used rosemary extracts to supplement freezing extenders.
Our results indicated that purslane extracts, cysteine, and their combinations resulted in higher percentages of living sperms with active mitochondria compared with the control treatment. Studies have shown that mitochondrial performance was positively associated with sperm quality and fertilization capability. 61 Within sperm cells, mitochondria provide the energy needed for different cell functions, including sperm motility. 62 These findings support our results regarding the observation that sperms with more active mitochondria have higher motility.
Furthermore, mitochondria play a crucial role in apoptosis development so that increased membrane permeability results in the release of cytochrome C into the cytosol, which activates caspase 9 and other effective caspases, leading to apoptosis.63,64 Our results showed an increase in the number of living sperms due to the use of aqueous and methanolic extracts and/or of each in combination with cysteine. In addition, methanolic extracts and the combination of cysteine with either aqueous or methanolic extracts resulted in the reduction of dead and apoptotic goat sperms after thawing.
These results are supported further by other investigations reporting that supplementation of semen extenders with an aqueous extract of fennel 65 and a methanolic extract of Albizia harveyi 66 increased the number of post-thaw live sperms and decreased the number of apoptotic ones. For this reason, it is assumed that purslane extracts and their combinations with cysteine could protect goat sperms against apoptosis during the cryopreservation processes by improving mitochondrial membrane integrity and, as a result, preventing ROS leakage.
When sperms are exposed to oxidative environments, it may result in base oxidation and in DNA degeneration. 67 In addition, given that sperms with damaged DNA can be motile and have normal morphology, in cases where the number of them is high during the artificial insemination stage, the likelihood of their participation in the fertilization process increases, and they affect the embryo quality. 68 Our results showed reduced DNA fragmentation resulting from using aqueous and methanolic extracts, as well as a combination of each with cysteine. Luno et al. 69 observed a decrease in the percent of post-thawing sperms with oxidized DNAs as a result of adding mate tea to the boar cryopreservation extenders. The mechanism(s) these antioxidants use to mitigate sperm damage remains to be understood. The conclusion stated that flavonoids probably prevented DNA from being damaged by transferring electrons to ROS-induced sites of free radicals on DNA.
In comparison with cysteine, methanolic extracts of purslane showed the most significant effect of extender supplementation as increasing motility and mitochondrial activity and decreasing MDA levels, indicating that the antioxidative power of methanolic purslane extracts is higher than that of cysteine.
Our results indicated that the extender containing combined cysteine and methanolic extract improved motility, viability, plasma membrane integrity, mitochondrial activities, and the number of live sperms while lowering MDA levels compared with others. In addition, the extender containing the combined cysteine and methanolic extract resulted in lowest/highest lipid peroxidation (LPO)/sperm motility rates compared with others. Also, the extender containing combined cysteine and methanolic extracts showed the highest mitochondrial activity rates in frozen–thawed samples.
Therefore, it can be concluded that cysteine combined with 50 μg/mL of aqueous, methanolic, or ethanolic extracts of purslane can be a promising cryoprotective and antioxidative compound in the processes of goat cryopreservation, which, given the best result obtained from the cysteine and PME combination, indicates the existence of some better synergetic effects between cysteine and compounds existing in PME. However, much more research is required to detect the precise antioxidative mechanisms of purslane extracts in combination with cysteine during the cryopreservation processes.
Footnotes
Acknowledgments
The researchers are grateful to all the Markhoz Goat Breeding Station performance test staff, Sanandaj, Kurdistan, Iran, particularly Ing. Kh. Jafari and Ing. Sh. Rashidi, for their sincere collaboration and permission to use their research facilities. Moreover, the authors also thank Dr. Ako Rezaei for his very friendly cooperation, particularly during semen collection.
Author Disclosure Statement
The authors declare no conflict of interest prejudging the impartiality of this scientific work.
Funding Information
This research did not receive any specific grant from any funding agency in the public, commercial, or not-for-profit sector.