
Abstract
The effect of antifreeze protein (AFP) as a cryoprotectant used in different concentrations of glycerol on post-thaw quality of epididymal sperm was investigated. Sperm were isolated from 50 testicles, obtained from 25 healthy mature goat bucks, with progressive motility >80%, and total morphological abnormalities <10% were pooled in each replication. The semen samples were diluted with Tris-citrate-fructose-soybean lecithin extender containing different concentration of AFP [0 μg/mL (A0), 5 μg/mL (A5), 10 μg/mL (A10)]. Each concentration of AFP was added in an extender containing either 7% (G7) or 5% (G5) glycerol. Post-thaw total and progressive motility were found to be higher (p < 0.05) in groups A5G5 and A5G7. Plasma membrane integrity, sperm acrosome integrity, DNA integrity, acrosome intact sperm, and mitochondrial membrane potential were found to be higher (p < 0.05) in groups A5G5 and A10G5. Sperm viability was found to be higher (p < 0.05) in group A5G5, while lipid peroxidation was recorded lower (p < 0.05) in groups A5G5 and A5G7. Regarding the apoptosis occurrence, the results demonstrate higher (p < 0.05) live post-thawed spermatozoa for groups containing 5 μg/mL AFP with 5% and 7% glycerol in addition to the lowest (p < 0.05) value for groups containing 0 μg/mL AFP with 5% and 7% glycerol. Based on these results, the present study concludes that the addition of 5 μg/mL AFP in combination with 5% glycerol in freezing extender improves the post-thaw quality, structure, and function parameters for buck spermatozoa.
Introduction
Cryopreservation is used to store many biological materials for an extended period at very low temperature.1–3 Sperm cryopreservation allows for specific opportunities for the conservation of genetic resources through existing sperm banks and the guarantee of a constant commercial supply of semen, as well as improving reproductive technologies.4–6
Despite the benefits and successes of sperm cryopreservation, the procedure currently used causes ultrastructural, biochemical, and functional damage to spermatozoa. 7 Cellular damage during cryopreservation is a complex multifactorial process caused by intracellular and extracellular ice formation, mechanical stress, oxidative stress, freeze-dehydration, and ice recrystallization.1,8,9 These lead to cell death through cell rupture, necrosis, and apoptosis, which ultimately decrease the fertilizing capacity of sperm.10,11 Intracellular ice crystallization during cryopreservation is the main cause of damage to the cells. Therefore, the composition of extender and suitable cryoprotectants are important factors for successful semen cryopreservation. 12
Glycerol (molecular weight: 92.10), being a permeable cryoprotectant, has a tremendous beneficial role in freezing extenders to preserve germ cells of multiple species.1,13 For goats, the usual concentration of glycerol is 5%–7% in freezing extenders. Apart from its usefulness, it has been reported that when glycerol is used at higher concentrations, it causes toxicity in spermatozoa by denaturation of protein, alteration of actin interactions, and induction of plasma membrane frangibility during cryopreservation.14,15 This toxicity arises from osmotic stress due to the slow permeability of glycerol to the cell membrane.16–18 Therefore, it is imperative to decrease the glycerol concentration in freezing extenders and to explore new cryopreservation regimes for efficient cryopreservation of goat buck sperm.
Antifreeze proteins (AFPs) are ice-binding proteins produced in some species of plants, insects, fungi, bacteria, and fish that live in subzero temperatures. 19 Acting as natural cryoprotectants in those species,20–23 these proteins perform several functions. They stabilize cell membranes,24,25 induce thermal hysteresis, and reversibly inhibit ice crystal growth and recrystallization.26,27 Moreover, they inhibit the leakage across the membranes,28,29 reduce the kinetics of ice formation, and influence the morphology of ice crystals. 23
AFPs can be used in cryopreservation and vitrification protocols as cryoprotectants in the cryopreservation of living cells and organs30,31 in concentrations of 3 to 500 times less than chemical cryoprotectants such as ethylene glycol.32,33 Therefore, the pressure of the osmotic solution around spermatozoa is less likely to change, 34 which is vital to keep spermatozoa inactive. 35 Furthermore, the use of AFPs reduces the use of high doses of penetrating chemical cryoprotectants that consequently reduce the toxicity of the penetrating chemicals.
To our knowledge, no report is available for the use of AFP to freeze buck sperm. Accordingly, this study was designed to evaluate the effect of AFP and to investigate whether AFP can be replaced with glycerol or a reduced level of glycerol. To this end, different concentrations of AFP and glycerol were used in the extender to assess the post-thaw quality of buck sperm.
Materials and Methods
Ethical statement
According to the international guidelines, all experimental procedures have been performed which were approved with the student number 9415000101 at 05.05.2018 by the Animal Care and Use Committee of the University of Kurdistan, Sanandaj/Iran.
Chemicals
All chemicals used were purchased from Sigma-Aldrich Company (St. Louis, MO) unless otherwise indicated. The AFPs were obtained from A/F Protein Inc. (Waltham, MA) and diluted according to the manufacturer's instructions.
Preparation of spermatozoa and cryopreservation process
A total of 50 testicles from 25 healthy mature goat bucks (4–5 years and 75–80 kg) were obtained from a local slaughterhouse. They were transported to the laboratory at room temperature. Spermatozoa were isolated from the cauda segments of the epididymis by the procedure described by Mondal et al. 36 The Cauda epididymis was cut into several pieces and suspended in modified Tyrode lactate solution to isolate the spermatozoa. The composition of the medium consisted of 100 mM NaCl, 3.1 mM KCL, 25 mM NaHCO3, 0.29 mM NaH2PO4H2O, 21.6 mM Na Lactate, 2.1 mM CaCL2 2H2O, 0.4 mM MgCL2 6 H2O, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 0.0006 g/mL Bovine serum albumin, 1 mM sodium pyruvate, 25 mg/mL Gentamycin, and 10 mg/L Phenol red. 37 After 15 minutes, the sperm suspension was centrifuged at 700 g for 10 minutes at room temperature, then prepared for the studies. Semen with progressive motility >80% and total morphological abnormalities <10% were pooled 38 in each replication and diluted with the basic extender to 250 × 106 spermatozoa/mL as the final concentration. The basic extender was composed of 27.1 g/L Tris, 10 g/L fructose, and 1.99 g/L citric acid, containing 1% (w/v) soybean lecithin. The basic extender was divided into two aliquots to dissolve glycerol in two concentrations (5% and 7% v/v) termed G5 and G7. Each glycerol extender (G5 and G7) was further subdivided into three aliquots to dissolve different concentrations of initially prepared AFP I (0, 5, and 10 μg/mL) termed as A0, A5, and A10, respectively.
The group containing 7% glycerol without AFP I (A0) was termed as the control. In this way, a total of six groups were made as to the factorial of 2 glycerol concentrations (G5 and G7) × 3 AFP I concentrations (A0, A5, and A10). After preparation, the osmolality of the different experimental extenders (A0G7, A5G7, A10G7, A0G5, A5G5, and A10G5) was measured as 320, 323, 325, 315, 319, and 321 mOsm/kg, respectively. The pH was maintained at 7.2 for all the extenders. For the preparation of the AFP solution, AFP I (A/F Protein Inc.) was dissolved in tris–citrate acid–glucose buffer (313.8 mM tris, 103.1 mM citric acid, and 33.3 mM glucose) 36 and added to semen at a concentration of 0 (control), 5, and 10 μg/mL.
The freezing procedure was carried out in two steps. In the first step, the extender and semen samples were cooled slowly at 5°C for 2 hours to reach the thermal equilibrium. Then, a semen extender (precooled at 5°C) was added to the semen to provide a final concentration of 100 × 106 sperm/mL. After 1 hour, the samples were aspirated into 0.25 mL French straws with a suction pump and sealed with polyvinyl alcohol powder.39–41 Then, the straws were frozen in liquid nitrogen vapor, 5 cm above the liquid nitrogen for 12 minutes and then plunged into the liquid nitrogen for storage. The frozen straws were thawed individually (37°C) for 30 seconds in a water bath for evaluation.
Semen quality assessment assays
Sperm motility and velocity parameters
Motility characteristics were assessed using the computer-assisted sperm motility analysis system (CASA: IVOS version 12; Hamilton-Thorne Biosciences, Beverly, MA). CASA was set up as follows: phase contrast; frame rate −60 Hz; minimum contrast −70; low and high static size gates −0.6 to 4.32; low and high intensity gates −0.20 to 1.92; low and high elongation gates 7–91; default cell size −10 pixels; default cell intensity −80. Thawed semen was diluted with a Tris-based extender (5 μL semen +95 μL extender without soybean lecithin and glycerol) and evaluated immediately after dilution. A 5 μL sample of diluted semen was put onto a prewarmed chamber slide (Leja 4, Leja Products Luzernestraat B.V., Holland), and sperm motion characteristics were determined using a 10 × objective at 37°C as follows.
Total motility (%), progressive motility (%), average path velocity μm/s (VAP), straight linear velocity μm/s (VSL), curvilinear velocity μm/s (VCL), amplitude of lateral head displacement, μm (ALH), [VSL/VCL] × 100 (Linearity, [LIN]), [VAP/VCL] × 100 (Wobbing [WOB]), [VSL/VAP] × 100 (Straightness [STR]). For each evaluation, 10 microscopic fields were analyzed to include at least 200 cells.
Plasma membrane integrity
The Hypo-Osmotic Swelling test was used to assess the functional membrane integrity of sperm cells. For this purpose, 20 μL of semen were mixed with 200 μL of hypoosmotic solution [fructose (9 g/L) and sodium citrate (4.9 g/L), 100 mOsm/kg] and kept at 37°C for 60 minutes. A drop of this mixture was placed on a prewarmed slide, and a cover slip was then put on it. A total of 200 sperm cells were counted in at least five different microscopic fields (400 × Olympus CX41, Tokyo, Japan). Next, the percentages of sperms with swollen and curled tails representing sperms with intact plasma membranes were determined. 14
Sperm viability
The vitality of spermatozoa was evaluated by the vital stain Trypan blue, which penetrates the postacrosomal region of dead cells. 42 The sperm sample was diluted with 0.4% trypan blue solution in a 1:1.2 (v/v) ratio and then incubated at 37°C for 15 minutes. To stop the movement of spermatozoa during evaluation, 10 μL of formol saline [0.9% (w/v) NaCl and 0.1% (v/v) 40% formaldehyde in distilled water] 43 was added to the sample. Microscopic slides were prepared by placing a drop of the stained sample on the slide and covering it with a coverslip. Slides were examined under the 1000 × magnification, with immersion oil, of a bright-field microscope. A total of 200 sperm heads was counted, and the percentage of viable (unstained) spermatozoa was calculated and noted.
Lipid peroxidation
Lipid peroxidation in spermatozoa was measured by the reaction of thiobarbituric acid (TBA) with malondialdehyde (MDA) according to Esterbauer and Cheeseman. 44 One milliliter of the thawed sample was added to a tube containing 1 mL of ethylenediaminetetraacetic acid (0.037 g ethylenediaminetetraacetic acid [EDTA] in 10 mL distilled water), 1 mL butylated hydroxytoluene (BHT) (0.2 g BHT in 10 mL ethanol), and 2 mL trichloroacetic acid (TCA) (3 g TCA in 30 mL distilled water). The mixture was centrifuged for 15 minutes at 1200 g. One milliliter of supernatant was incubated with 1 mL of TBA (0.134 g TBA in 20 mL distilled water), heating for 20 minutes in a water bath at 90°C. After cooling, the absorbance was determined by a spectrophotometer (Bausch and Lomb Supertonic 70, Feldkirchen, Germany) at a wavelength of 532 nm.
Results were compared to a standard curve previously prepared with a standard solution of malondialdehyde. The malondialdehyde concentration (nmol/mL) was determined using the value of 1.56 × 105/M cm3 as the MDA extinction coefficient. Results were also calculated for Thiobarbituric Acid Reactive Substances levels normalized to sperm concentration, as more sperm may present more substrate for lipid peroxidation.
DNA integrity
A sperm chromatin structure assay and flow cytometer were used to analyze the DNA integrity of the stored spermatozoa, as described by Evenson and Jost. 45 Semen samples were diluted in basic extender to a final concentration of 1 × 106 spermatozoa per mL. The suspension (200 μL) was subjected to brief acid denaturation by mixing with 400 μL of lysis solution (0.1% Triton X-100 (v/v), 0.15 M NaCl, 0.08 M HCl). Exactly 30 seconds later, 1.2 mL of acridine orange (AO) staining solution (6 μg/mL AO, 0.037 M citric acid, 0.126 M Na2HPO4, 1 mM disodium EDTA, and 0.15 M NaCl, pH 6.0; final stain concentration of 4 μg/mL) was added. After 3 minutes samples were aspirated into the flow cytometer. The sperm chromatin damage of spermatozoa was quantified by the metachromatic shift from green (native, double-stranded DNA) to red (denatured, single-stranded DNA) fluorescence and displayed as red vs. green. Green fluorescence was detected on the FL1 detector and red fluorescence with detector FL3 on the flow cytometer. Using CellQuest software (BD, version 5.2.1) cell populations were gated as (1) green-fluorescing double-stranded DNA (classified as intact) and (2) red-fluorescing cells-outside-main-population denatured DNA. 46
Sperm chromatin structure assay
For each sample, 5000 sperm cells were analyzed to determine the extent of DNA denaturation. The DNA fragmentation index (DFI) was calculated based on the ratio of red/total (red + green) fluorescence for each sperm cell in the sample. For each sample, the percentage of spermatozoa with detectable DFI (%DFI) was evaluated based on the level of red fluorescence. 47
Mitochondrial membrane potential
Mitochondrial membrane potential (MMP) was evaluated by a JC-1 (5,5,6,6-tetrachloro-1,1,3,3-tetraethylbenzimidazolylcarbocyanine iodide) probe that was able to selectively penetrate mitochondria. Briefly, samples were diluted with phosphate-buffered saline buffer to a concentration of 1 × 106 spermatozoa/mL, then 300 μL of each sample was transferred to a polypropylene tube, and 1.2 μL of JC-1 stock solution (1.5 mM JC-1 in dimethyl sulfoxide) was added. The tubes were incubated at 37°C for 30 minutes in the dark. The stained sperm samples were then analyzed by flow cytometry. According to the manufacturer (Molecular Probes), JC-1 reversibly changes its fluorescence from green (monomeric status) to orange (multimeric status) when the mitochondrial membrane becomes more polarized.
In viable cells with normal membrane potential, JC-1 is in the mitochondrial membrane in the form of aggregates emitting an orange fluorescence, while in cells with low membrane potential, it remains in the cytoplasm in a monomeric form, giving a green fluorescence. Therefore, it is possible to distinguish two cell populations: cells with damaged MMP where JC-1 remains in the cytoplasm in a monomeric form, giving a green fluorescence, and cells with normal MMP with a double fluorescence where JC-1 (in addition to emitting green in the cytosol) is also in the mitochondrial membrane in the form of aggregates emitting an orange fluorescence, which identifies cells with high MMP (orange-stained cells).48,49
Acrosome membrane integrity
Acrosome-reacted sperm were evaluated using fluorescein-labeled Pisum sativum agglutinin (FITC-PSA) as previously described by Thys et al. 50 In brief, 15 μL of sperm was smeared onto a slide, allowed to dry, and then fixed and permeabilized in absolute ethanol (purity: 96%) for 10 minutes. After fixation, the smears were covered with 50 μL FITC-PSA (100 μg/mL) and placed in a moist environment in the dark at room temperature for 30 minutes. The slides were rinsed with Tris-buffered saline (213 mM NaCl, 50 mM Tris-HCl, pH 7.0), then mounted with glycerol and a coverslip. Two hundred sperm per slide were counted by a fluorescent microscope (BX51, Olympus) equipped with fluorescence illumination and a fluorescein isothiocyanate (FITC) filter (excitation at 455–500 nm and emission at 560–570 nm) at × 400 magnification. Sperm heads that fluoresced green were considered to have intact acrosomes, and those with no fluorescence were recorded as damaged or disrupted acrosomes.51,52
Phosphatidylserine translocation assay
The percentage of apoptotic cells in spermatozoa was assessed through the detection of phosphatidylserine (PS) translocation through a commercial kit and according to the manufacturer's instructions (Immune Quality Products, Groningen, The Netherlands). At the onset of apoptosis, PS is normally found on the cytoplasmic face of the plasma membrane and translocated to the extracellular leaflet. Annexin V Cy3.18 binds to PS on the exterior surface of membranes of cells that are undergoing apoptosis.
A second stain, FITC, is used to assess the viability and differentiate between apoptotic and necrotic cells. A total of 1 × 106 washed spermatozoa/mL (using calcium buffer) were incubated with 10 mL FITC labeled Annexin V, at 37°C for 30 minutes in the darkness followed by adding 10 mL propidium iodide (PI). The sperm sample was analyzed in a FACSCalibur flow cytometer, and subpopulations were classified as follows: (1) Live spermatozoa (Annexin−/PI−), (2) Apoptotic spermatozoa (Annexin+/PI−), and (3) Dead spermatozoa (PI+).51,53
Flow cytometry analysis
All fluorescence signals of labeled spermatozoa were analyzed by the flow cytometer FACSCalibur (Becton Dickinson, San Khosoz, CA) equipped with standard optics. A minimum of 10,000 spermatozoa was examined for each assay at a flow rate of 100 cells/s. The sperm population was gated using 90° and forward angle light scatter to exclude debris and aggregates. The excitation wavelength was 488 nm supplied by an argon laser at 250 mW. Green fluorescence (FL1) was measured using a 530/30 nm bandpass filter, red fluorescence (PI, FL3) was measured by a 585/42 nm filter, and Rh123 was measured using a 527/25 nm filter (FL3). The percentage of PI-positive cells and the mean fluorescence were calculated on a 1023-channel scale. The analysis of flow cytometry data was performed using FlowJo software (Treestar, Inc., San Carlos, CA).
Statistical analyses
The experiment was conducted as a completely randomized 2 × 3 factorial design, and each treatment was replicated six times. The data were analyzed by the Generalized Linear Model procedure of SAS 9.2. Results were expressed as mean ± standard error of the mean, and they were compared by the Duncan test at p < 0.05. All data were evaluated for normal distribution by the Shapiro–Wilk test. The model used in the present study was as follows: Yijk = μ + Ti + Dj + (T × D) ij + eijk; where Yi stands for the observed dependent variables, including sperm variables, μ is the mean of the population, Ti is the effect of treatment (Glycerol Concentrations, i = 5, 7), Dj is the effect of treatment (AFP concentrations, j = 0, 5, 10), (T × D) ij is the effect of interaction between glycerol and AFP, and eijk is the random residual error.
Results
Sperm motility and velocity parameters
As shown in Table 1, groups containing 5 μg/mL antifreeze protein (AFP) resulted in the highest (p < 0.05) percentage of total and progressive motility compared to the control and other treatment groups. Both parameters were higher (p > 0.05) in A5 when used in G5 and G7 compared to A10 at the same concentration of glycerol. The lowest (p < 0.05) progressive motility was observed in groups containing 0 μg/mL AFP compared to other groups. VAP was found to be lowest (p < 0.05) in the A0G7 group, and VSL, VCL, and ALH were found to be lowest (p < 0.05) in groups without AFP (A0) compared to other treatment groups.
Effect of Different Concentration of Antifreeze Protein (AFP) and Glycerol on Motility and Velocity Parameters of Post-Thawed Goat Epididymal Spermatozoa (Mean ± Standard Error of the Mean)
The parameters of TM, PM, VAP, VSL, VCL, ALH, STR, and WOB were shown when buck sperm was frozen with six different cryopreservation media. A: AFP (A0: 0 μg/mL AFP, A5: 5 μg/mL AFP, A10: 10 μg/mL AFP), G: Glycerol (G5: 5% Glycerol, G7: 7% Glycerol). Different letters within the same row show significant differences (p < 0.05).
ALH, amplitude of lateral head displacement; LIN, Linearity; PM, progressive motility; STR, straightness; TM, total motility; VAP, average path velocity; VCL, curvilinear velocity; VSL, straight linear velocity; WOB, wobbing.
In addition, VAP, VSL, VCL, and ALH were found to be the highest (p < 0.05) in the group containing 5 μg/mL AFP and 5% glycerol (A5G5) compared to other groups. The lowest (p < 0.05) STR and LIN were observed in groups A0G5 and A0G7, respectively, compared to other groups. Furthermore, there were no differences (p > 0.05) in WOB between the groups.
Plasma membrane integrity and sperm viability
According to the results, post-thaw plasma membrane integrity (Fig. 1) was higher (p < 0.05) when antifreeze protein (AFP) 5 μg/mL and AFP 10 μg/mL were added with 5% glycerol compared to 7% glycerol at the same concentration of AFP and control group (A0G7). In terms of plasma membrane integrity, no difference (p > 0.05) was observed between the A5G7 and A10G7 groups. Plasma membrane integrity is the lowest (p < 0.05) in the control group (A0G7).

Effect of different concentrations of AFP and glycerol in extender on plasma membrane integrity (HOS test) of post-thawed buck epididymal spermatozoa. Different letters
The highest (p < 0.05) value of viability was obtained for the A5G5 group compared to other groups. In terms of sperm viability, no (p > 0.05) differences were observed between A5G7, A0G5, and A10G5 groups. Sperm viability was lower (p < 0.05) in A10G7 and A0G7 groups when 7% glycerol is used without the addition of AFP and 10 μg/mL AFP compared to other treatment groups (Fig. 2).
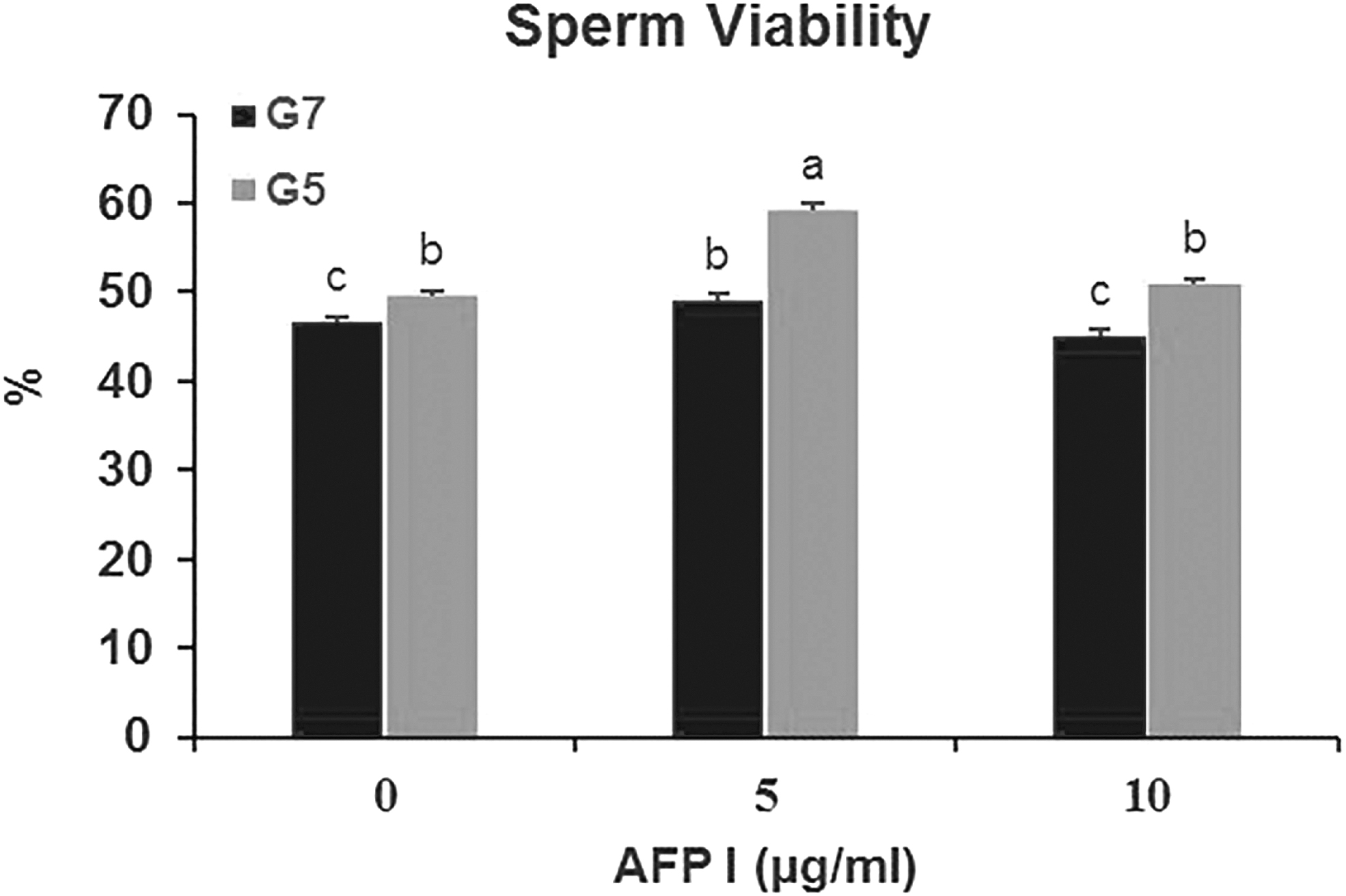
Effect of different concentrations of AFP and glycerol in extender on sperm viability of post-thawed buck epididymal spermatozoa. Different letters
Lipid peroxidation
The obtained results showed that MDA concentration was lower (p < 0.05) in the A5G5 and A5G7 groups compared to other groups, while no (p > 0.05) difference was observed between other treatment groups (Fig. 3).

Effect of different concentrations of AFP and glycerol in extender on malondialdehyde concentration of post-thawed buck epididymal spermatozoa. Different letters
DNA integrity
We observed no difference (p > 0.05) in the percentage of sperm with fragmented DNA post-thawing between treatment groups. However, sperm with fragmented DNA was lower in the A5G5 group and was higher in the A0G5 group (Fig. 4).

Effect of different concentrations of AFP and glycerol in extender on DNA fragmentation index of post-thawed buck epididymal spermatozoa. The same letters
Mitochondrial membrane potential
MMP was found to be higher (p < 0.05) in the A5G5 and A10G5 groups compared to control and other treatment groups. This parameter was lowest (p < 0.05) in the A10G7 group compared to other groups. Moreover, no difference (p > 0.05) was found between the A0G7 and A5G7 groups (Fig. 5).
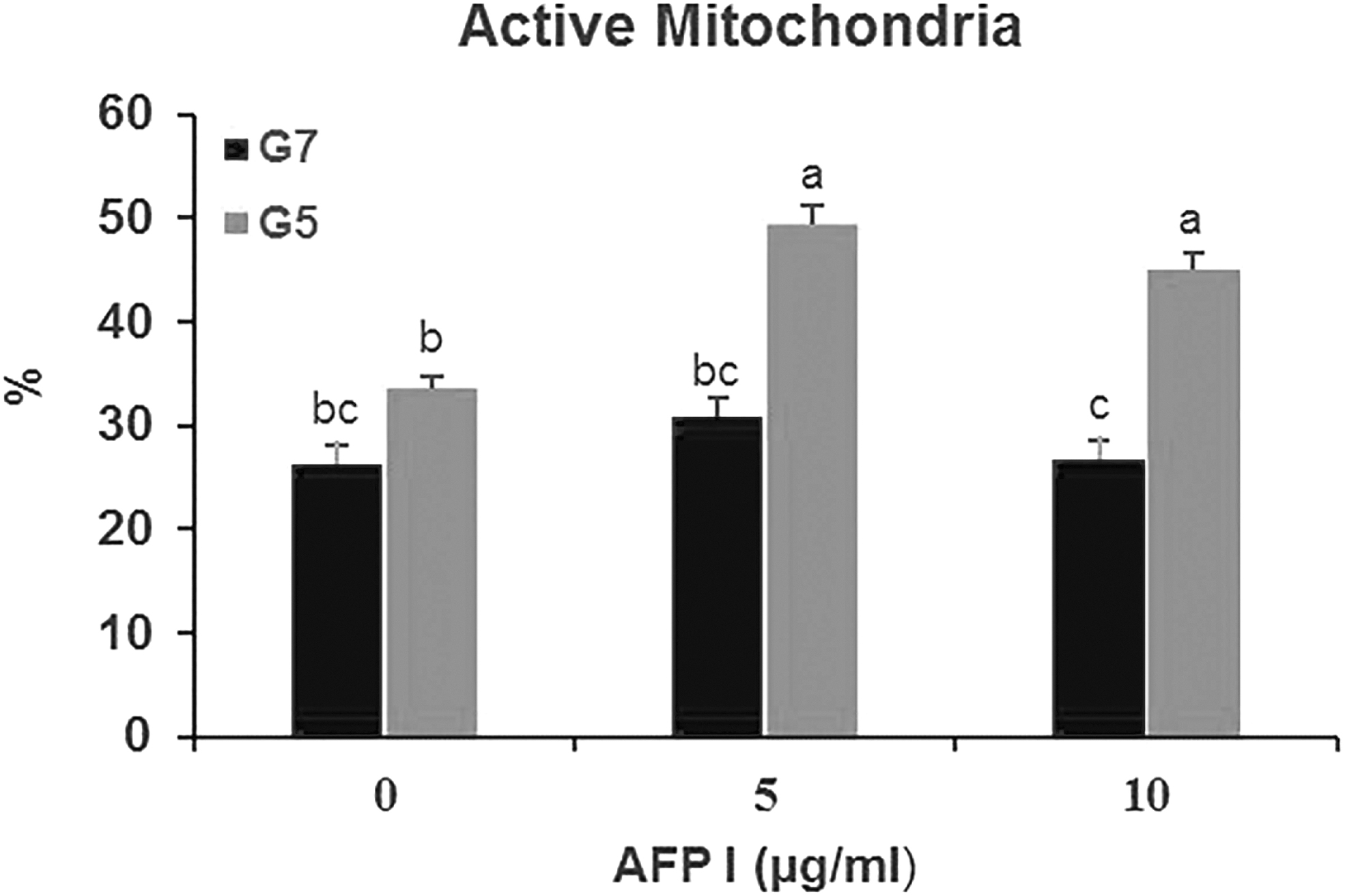
Effect of different concentrations of AFP and glycerol in extender on active mitochondria of post-thawed buck epididymal spermatozoa. Different letters
Acrosome membrane integrity
The results showed that acrosome integrity was higher (p < 0.05) in the A10G5 and A5G5 groups compared to the control and other groups. This parameter was lower (p < 0.05) in the A0G5 and A0G7 groups. Moreover, there was no (p > 0.05) difference between the A10G7 and A5G7 groups (Fig. 6).
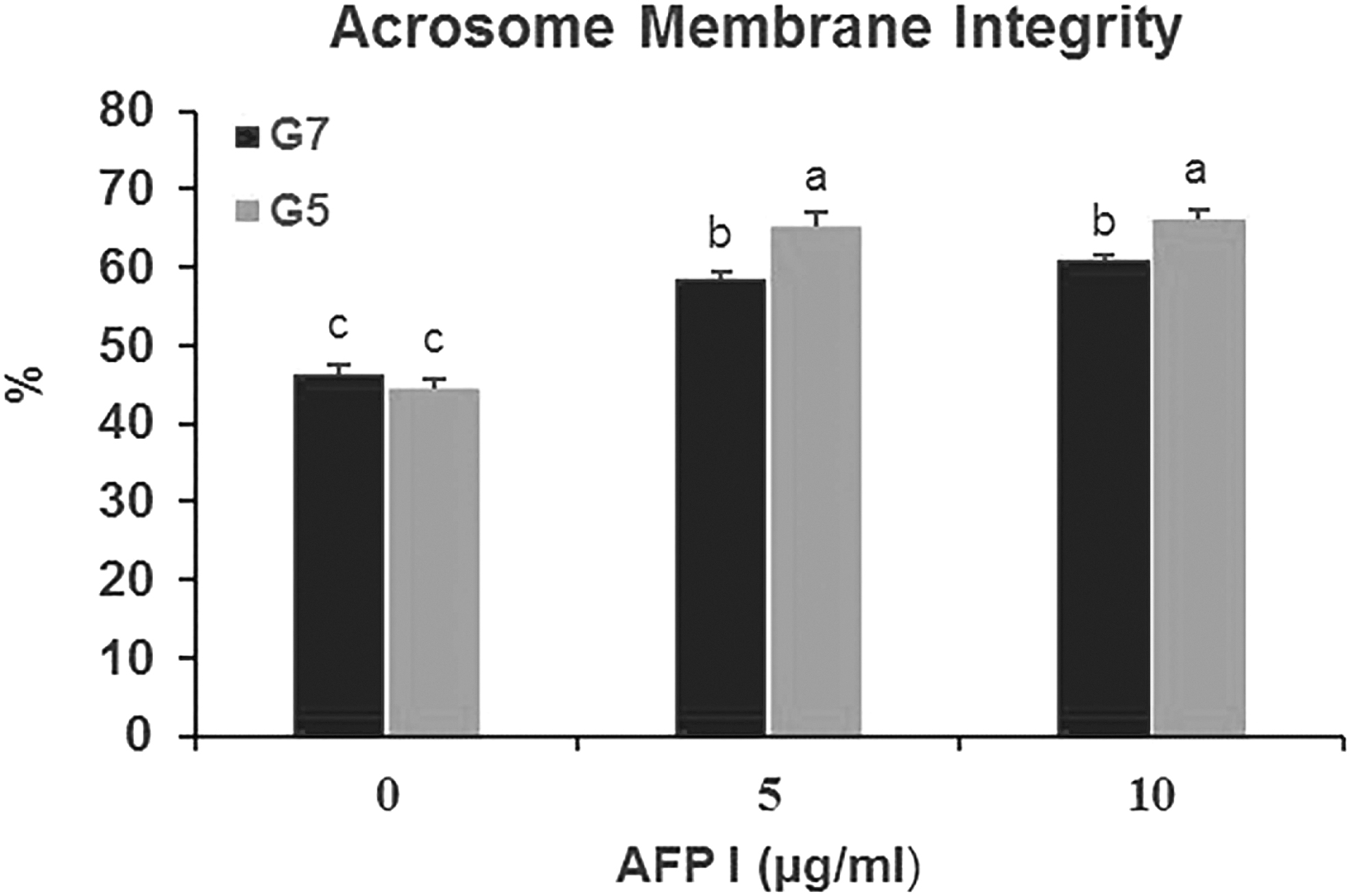
Effect of different concentrations of AFP and glycerol in extender on acrosome membrane integrity of post-thawed buck epididymal spermatozoa. Different letters
PS translocation assay
As represented in Figure 7, the A5G5 and A5G7 groups revealed a higher (p < 0.05) percentage of live and also the lower (p < 0.05) post-thawed dead spermatozoa where the extender was supplemented with 5 μg/mL AFP that is used with the 5% and 7% glycerol compared to other groups. However, A0G5, A10G5, A0G7, and A10G7 yielded the largest proportion of apoptotic spermatozoa with PS translocation being higher (p < 0.05) than other groups.
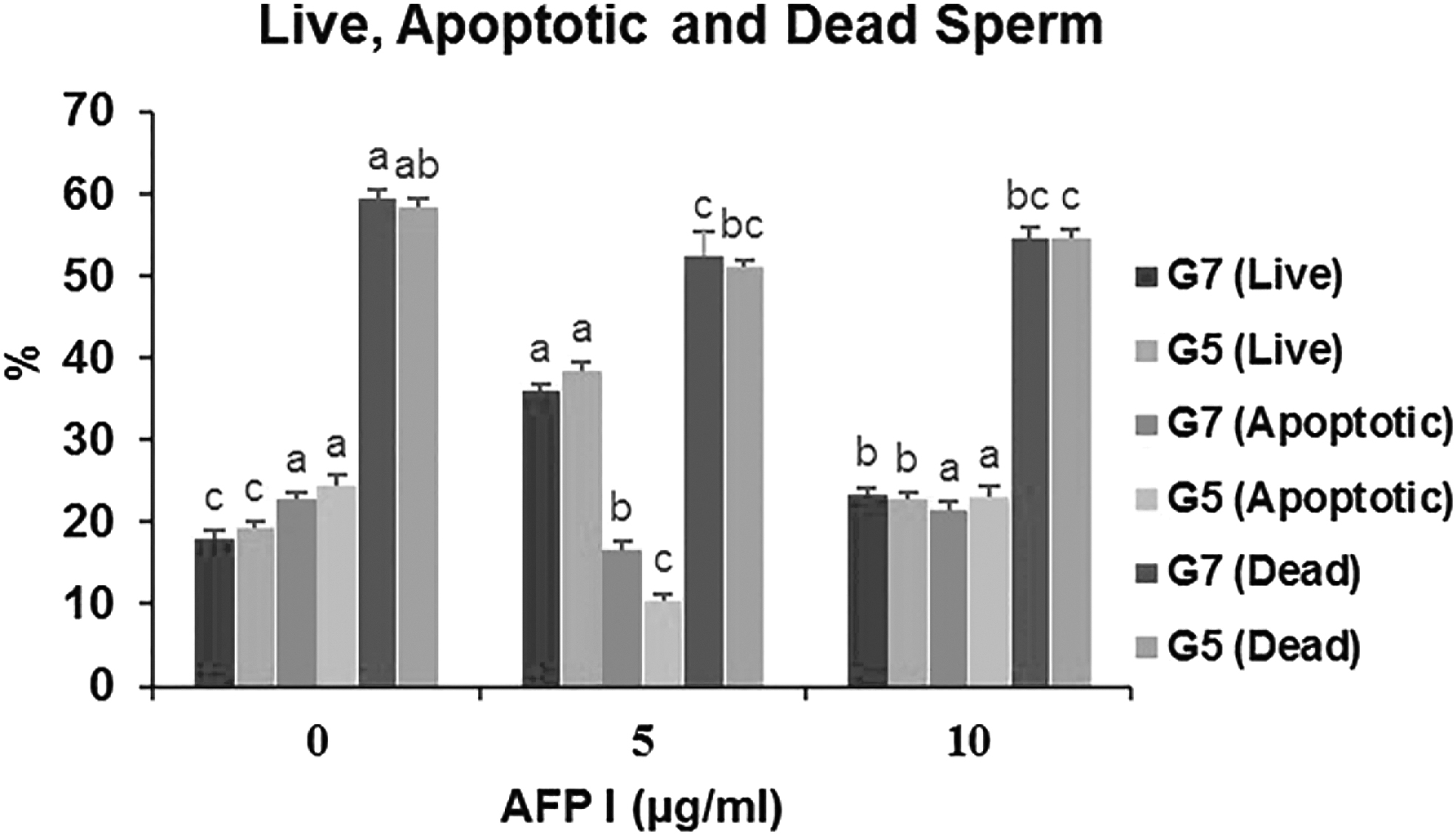
Effect of different concentrations of AFP and glycerol in extender on live, apoptotic, and dead of post-thawed buck epididymal spermatozoa. Different letters
Discussion
Different concentrations of cryoprotectant in the freezing solution were used to elucidate the antifreeze effect of AFP I and to investigate whether AFP I could be replaced by glycerol. Our results show significantly improved post-thaw plasma membrane integrity when AFP was used in combination with a reduced concentration of glycerol. The reason for these results is that AFPs are relatively high molecular mass molecules which can stabilize membranes during chilling and control ice crystal growth during the freezing of cells and tissues.34,54 The AFPs were reported to decrease the temperature at which ice crystals increase in size in a noncolligative manner and exhibit thermal hysteresis. 55 Due to this property, these proteins protect cell membranes by inhibiting ion leakage at hypothermic temperatures.26,29
In the present study, the extenders having 5% glycerol with 5 and 10 μg/mL AFP (A5G5 and A10G5) significantly decreased the sperm cell plasma membrane damage. Xin et al. 56 confirmed that the plasma membrane integrity of frozen-thawed sterlet sperm was greatly enhanced by AFPs (I and III). AFPs might protect the plasma membrane not only by preventing direct ice damage but also by interacting with the phospholipids, preventing the peroxidation of polyunsaturated fatty acids, as well as increasing the saturated fatty acid proportion.57,58
Sperm motility is considered to be the major but not the only prediction parameter for fertility. 59 It has a positive correlation with sperm cell viability and mitochondrial and cell membrane integrity. 60 The high total motility, progressive motility, and STR of sperm after thawing were observed when 5 μg/mL AFP with 5% and 7% glycerol was added to the semen extender. However, excessive AFP concentrations decreased the motility, progressive motility, and other parameters, such as VAP, VSL, VCL, ALH, and STR. The obtained results are consistent with the previous studies, in which AFP at high concentrations was mildly cytotoxic to ram 61 and chimpanzee spermatozoa. 20
Other studies showed protective or stimulating effects of AFPs on sperm motility. Shaliutina-Kolesova et al. 62 demonstrated that supplementing the extender with types I and III AFP (0.1, 1, and 10 mg/mL) had positive effects on the post-thawing motility and velocity of common carp, especially with AFP III. Some similar results have been obtained in sturgeon at 10 mM 63 and buffalo at 0.1 mg/mL. 21 Adding antifreeze protein and glycoprotein type I to semen extender significantly increased post-thaw motility in ram sperm, 61 whereas in bull, supplementation with antifreeze protein type I only improved the osmotic resistance of sperm during cryopreservation. 64
Samples cryopreserved in the presence of 5 μg/mL AFP and 5% glycerol compared to the other frozen groups presented a higher percentage of viable cells, sperm VAP, VSL, VCL, and ALH.
Antifreeze proteins can protect cells by decreasing the freezing point, modifying the ice-crystal formation process, preventing recrystallization, as well as interacting with plasma membranes at low temperatures. 65 Antifreeze proteins bind with ice crystals through a prism on the surface and cause the ice crystals to be more hexagonal. 66 A high sperm cell viability percentage was observed in this study when 5 μg/mL AFP with 5% glycerol was added to the semen extender. This finding is due to the lesser damage to spermatozoa by ice crystals forming in intracellular spaces during cryopreservation.
MDA is a key product of polyunsaturated fatty acid peroxidation in cells 67 and is often used as an indicator of oxidative damage. Sperm membrane integrity, livability, and motility are found to be negatively correlated with the concentration of sperm MDA.68,69 The high lipid peroxidation of the plasma membrane during freezing and thawing induces increased loss of membrane integrity and membrane leakiness. As a result of this successive leakage, loss of intracellular antioxidant enzymes occurs. 18 In the current study, we found a lower concentration of MDA in those groups that had 5 μg/mL AFP with 5% and 7% glycerol in the extender, which consequently led to higher motility in these groups. However, the authors didn't observe a logical relationship between the level of MDA and sperm membrane integrity and livability.
Sperm mitochondria has been specified as the main source of reactive oxygen species (ROS) in spermatozoa 70 and is also considered to be among the sperm structures prone to cryoinjuries during freezing. 71 Lipid peroxidation causes the high levels of ROS that increase sperm damage because sperm plasma membranes are rich in polyunsaturated fatty acids. 72 It has been reported that ROS production during cryopreservation was associated with a reduction in sperm motility and DNA damage. In the present study, we found higher MMP in those groups that had 5% glycerol with 5 and 10 μg/mL AFP in the extender. Cell membranes are vulnerable to damage by the active oxygen produced by lipid oxidation produced during sperm cryopreservation. The stabilization of the cell membrane by AFPs involves the binding of this protein to the biphospholipid layer of the cell membrane. Thus, AFP might have resulted in the binding of this compound to the mid piece where the mitochondria are located. 22 So, there would have been less damage to the mitochondrial membrane by ice crystallization during freezing and thawing.
The acrosomal reaction is an exocytotic calcium-dependent process that is considered the major prerequisite for sperm penetration through the oocyte membranes. 73 Accompanying the reduction in sperm motility, post-thawed sperm loses acrosome integrity, which might be indicative of the decrease in its fertilizing capacity. 22 In addition, we found higher intact acrosomes in those groups that had 5% glycerol with 5 and 10 μg/mL AFP in the extender. The results of this study are consistent with the results of the study done by Younis et al. 20 which showed an increased proportion of acrosome-intact cells in chimpanzee sperm following a freeze–thaw process in the presence of AFP III. The results of that study indicated that the AFPs stabilized the chimpanzee sperm membranes, thereby inhibiting capacitation and the subsequent acrosome reaction.
Rubinsky et al. 74 proposed that AFPs may act by blocking ion channels, thereby stabilizing transmembrane electrolyte gradients. Based on this theory, calcium influx would be decreased, partially inhibiting the capacitation reaction. The current study shows that the addition of 5 μg/mL AFP with a reduced concentration of glycerol (5%) in a freezing extender can enhance the structural and functional parameters of buck sperm.
Footnotes
Acknowledgments
The researchers are grateful to all local slaughterhouse staff situated in Sanandaj, Kurdistan, Iran for their study's sincere collaboration and permission to use the goat testes.
Author Disclosure Statement
The authors declare no conflict of interest prejudging the impartiality of this scientific work.
Funding Information
This research did not receive any specific grant from any funding agency in the public, commercial, or not-for-profit sector.