
Abstract
Background:
Plasma cell-free RNAs (cfRNAs) can serve as noninvasive biomarkers for the diagnosis and monitoring of diseases. However, the delay in blood processing may lead to unreliable results. Therefore, an unbiased evaluation based on the whole transcriptome under different storage conditions is needed.
Methods:
Here, blood samples were collected in ethylenediaminetetraacetic acid tubes and processed immediately (0 hour), or stored at room temperature (RT) or 4°C for different time intervals (2, 6, and 24 hours) before plasma separation. High-throughput sequencing was applied to assess the effects of storage conditions on the transcript profiles and fragment characteristics of plasma cell-free mRNA, long noncoding RNA (lncRNA), and small RNAs.
Results:
More genes changed their expression levels with time when blood was stored at RT compared with those at 4°C. Cell-free mRNA and lncRNA were relatively stable in blood preserved at 4°C for 6 hours, while cell-free microRNA (miRNA) and piwi-interacting RNA (piRNA) remained stable at 4°C for 24 hours. After 24 hours, more contamination of the leukocyte-derived RNAs occurred at RT, possibly due to apoptosis. Meanwhile, significant changes were also observed regarding the characteristics of the RNA fragments, including fragment size, the proportion of intron, and the pyrimidine frequency of the fragmented 3′ end. Fifteen tissue-enriched genes were detected in the plasma but not expressed in leukocytes. The expression level and fragment length of these genes gradually decreased during storage, suggesting the degradation of the cfRNA and the dilution of leukocyte-derived RNA with other tissue-derived cfRNA.
Conclusions:
Our results suggest that the contamination of leukocyte-derived RNA and the degradation of original cfRNA contribute to the changes in the cfRNA expression profiles and the fragment characteristics during short-term storage. The storage of blood at 4°C for 6 hours allows plasma cfRNA to remain relatively stable, which will be useful for further studies or clinical applications where adequate quantification or the fragment signature of cfRNA is required.
Introduction
Plasma cell-free messenger RNA (cf-mRNA),1,2 long noncoding RNA (cf-lncRNA), 3 microRNAs (cf-miRNA), 4 transfer RNA (cf-tRNA), 5 and piwi-interacting RNA (cf-piRNA) 6 have been used as noninvasive circulating biomarkers for the monitoring or diagnosis of a wide variety of diseases, such as cancers, pregnancy complications, and neurological diseases. However, due to the short half-life of cell-free RNA (cfRNA) and the potential contamination of cellular RNA, preanalytical variables may lead to discordant results of cfRNA-based liquid biopsies, limiting its clinical application. 7 Standardization of sample processing workflow will benefit by reducing these variations in the clinical application of plasma cfRNA.
Short-term storage and transportation of blood samples from the collection site to the laboratory are commonly required during research or clinical applications. Blood cells may undergo a series of alterations during the temporary storage, including cell apoptosis and stress response. 8 These alterations of the cells may impact the expression pattern of the original transcriptome in the plasma through cellular RNAs release. Plasma contains RNA from other tissues, 9 which is important for monitoring the healthy and diseased status of tissues. These RNAs cannot be replenished by blood cells, and the release of cellular RNA may dilute these RNAs, making them difficult to detect in subsequent analysis. Another possible mechanism that affects the identification of these genes in plasma is the degradation of cfRNA by ribonucleases (RNases). RNases exist in plasma 10 and can rapidly degrade exogenous RNA.11,12 RNase A, as one of the most abundant extracellular RNases, is released into the extracellular compartment as a secreted protein through various pathways. 13 RNase A prefers to cleave the phosphodiester bond where the 3′ end residue is a pyrimidine,14,15 and participates in RNA degradation.
Previous studies have shown that plasma cfRNA may exist inside vesicles, or form complexes with proteins or lipids,11,16 thus avoiding degradation by RNase. The endogenous cfRNA is more stable compared with the exogenously incorporated RNA, supporting the concept that cfRNA is protected by specific mechanisms.11,12 However, the concentrations of plasma miRNA were significantly decreased when isolated plasma was stored at 4°C for 2 weeks, 17 suggesting the possibility of degradation during storage. Elucidating the effect of short-term storage of whole blood on the expression profile of cfRNA can help develop a proper procedure for sample processing.
Several studies have been conducted to determine the optimal blood storage temperature and time for plasma cf-mRNA and cf-miRNA.18–20 However, these studies mainly focused on specific genes. Thus, an unbiased evaluation based on the whole transcriptome is needed. In addition, the fragment characteristics of plasma nucleic acids, such as fragment size and end motif, have been considered as molecular signatures in liquid biopsies. 21 However, no study has yet evaluated the effects of storage conditions on the fragment characteristics of plasma cfRNA.
Therefore, we used high-throughput sequencing to investigate the effects of different blood storage temperatures (room temperature [RT] and 4°C) and times (0, 2, 6, and 24 hours) on the transcript profiles and fragment characteristics of multiple cfRNAs. We also analyzed the leukocyte contamination and the decay of original plasma cfRNA by integrating the corresponding leukocyte transcriptome.
Materials and Methods
Sample collection, processing, and sequencing
A total of four healthy individuals were recruited in this study, including two males (M2 and M3) and two females (F2 and F3). Peripheral blood samples were collected in ethylenediaminetetraacetic acid (EDTA) tubes (BD, 0202992058), and treated immediately (0 hour) or stored at RT (18°C–22°C) or 4°C for various time intervals (2, 6, and 24 hours) before plasma separation (Fig. 1).
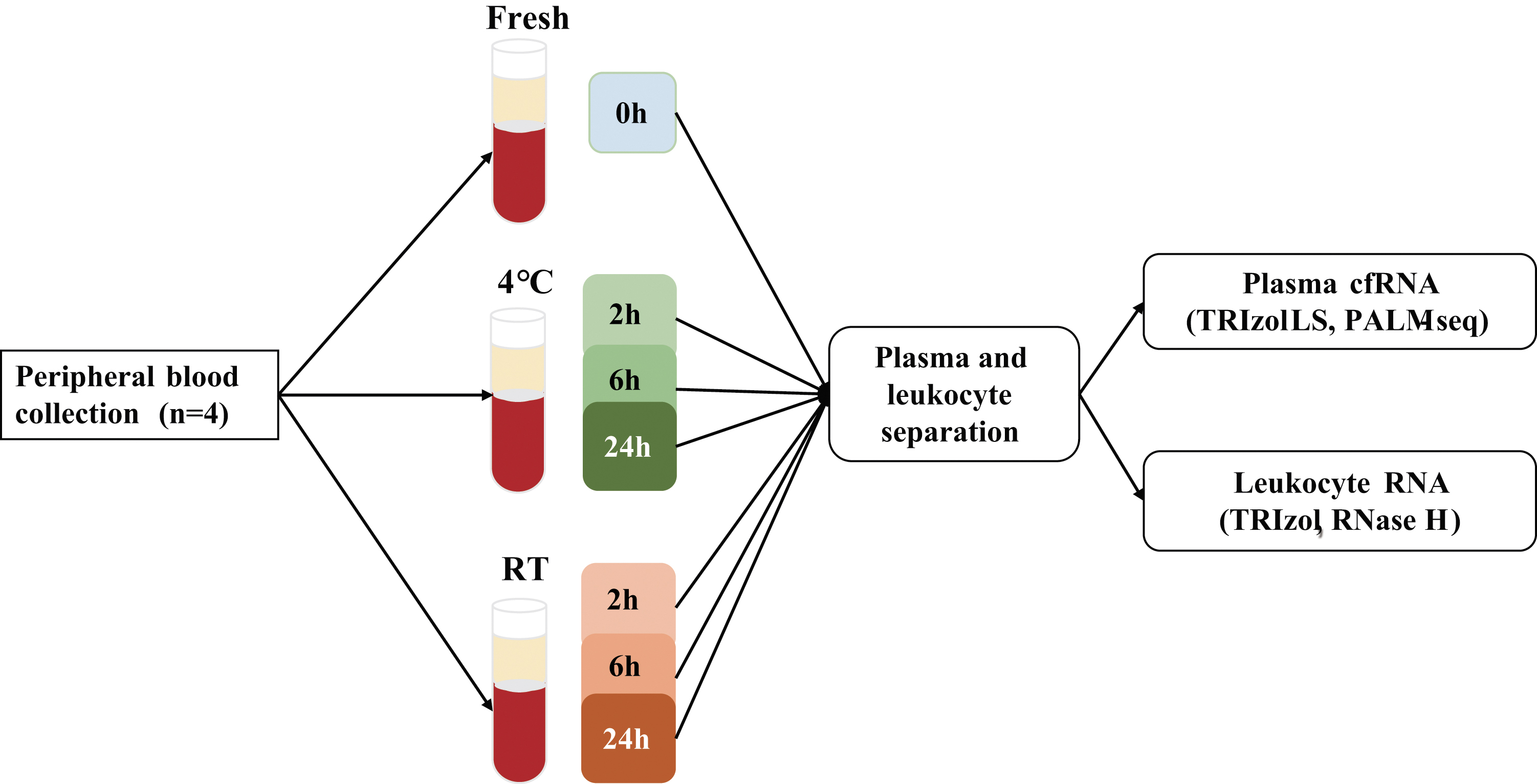
Samples processing. Peripheral blood samples were collected from four healthy subjects and treated immediately (0 hour), or stored at RT or 4°C for various time intervals (2, 6, and 24 hours) before plasma separation. Plasma cfRNA and leukocyte RNA data are integrated to elucidate the impact of short-term storage on plasma cfRNA. cfRNA, cell-free RNA; PALM-seq, PolyAdenylation Ligation Mediated-Seq; RT, room temperature.
For each storage condition, 1 mL blood was removed, and 300 μL plasma was separated according to the two-step centrifugation protocol: 1600 g for 10 minutes at 4°C followed by 12,000 g for 10 minutes at 4°C. Then, the supernatant was collected, and 900 μL TRIZOL LS Reagent (cat. 10296028; Thermo Fisher Scientific) was added and mixed immediately. cfRNA was extracted according to the manufacturer's protocol (cat. 10296028, Part No. 10296010.PPS; Thermo Fisher Scientific). RNase-free glycogen (cat. AM9510; Thermo Fisher Scientific) was added as a carrier. The transcriptome sequencing library was constructed using the PolyAdenylation Ligation Mediated-Seq (PALM-Seq) method, 22 which enables simultaneous sequencing of cell-free coding and noncoding RNA. Sequencing was performed on BGISEQ-500RS (single-end 100 bp) platform.
The corresponding leukocyte RNA was isolated and sequenced based on the ribonuclease (RNase) H method as previously described by Xing et al. 23 This study was approved by the Institutional Review Board of Shenzhen Maternity and Child Healthcare Hospital (SFYLS 2019 No. 139), and all participants provided written informed consent before blood collection.
Alignment and quantification
cfRNA sequencing data were processed following the previously published procedure. 22 In brief, adapter and low-quality reads were trimmed, then reads with “N” base (>10%) or <17 bp were filtered by cutadapt. 24 The clean reads were aligned to rRNA, vault RNA, and Y RNA reference (https://www.ncbi.nlm.nih.gov) by bowtie. 25 Then, the unmapped reads were further aligned to a transcriptome reference in the following order: miRNA (www.mirbase.org), tRNA (http://gtrnadb.ucsc.edu/index.html), piRNA (www.regulatoryrna.org/database/piRNA), and finally mRNA and lncRNA (https://www.gencodegenes.org). For miRNA, we combined mature and hairpin miRNA as a reference, but only mature miRNAs were retained for further analysis. The reads count of miRNA, tRNA, and piRNA was normalized to reads per million mapped reads. The expression levels of mRNA and lncRNA were estimated by RNA-seq by expectation-maximization (RSEM) 26 and normalized to transcripts per million mapped reads (TPM). Finally, the reads unmapped to transcriptome were aligned to the genome (hg19) using STAR. 27
Clean reads of leukocyte RNA were obtained as previously described. 23 Then, the clean reads were aligned to the mRNA and lncRNA transcriptome by bowtie2, 28 and quantified at the gene level by RSEM. 26
Differentially expressed gene analysis and functional analysis
Differentially expressed gene (DEG) analyses between immediately processed samples and samples stored for a period of time were performed using the DESeq2. 29 Genes were considered as DEGs if fold change ≥2 with an adjusted p-value <0.05. The pathway enrichment analysis was performed using Metascape. 30 Pathways with a q-value <0.01 were considered significant.
Platelet marker genes, housekeeping genes, and tissue-enriched genes
Platelet marker genes were downloaded from the PanglaoDB database. 31 Genes with a ubiquitousness index = 0 and marker count <4 were retained for further analysis. Eighteen mRNA housekeeping genes 32 and 6 miRNA housekeeping genes 33 were collected to examine the effects of short-term storage. Tissue-enriched genes (TEGs) were downloaded from the human protein atlas database. 34 The genes with expression levels greater than one and having at least fivefold higher expression levels in a particular tissue compared with all other tissues were classified as TEGs. Twenty-three tissues with ∼10 TEGs were considered for further analysis. TEGs with a TPM >0 in at least two plasma samples and TPM ≥1 in at least one plasma sample were included in the TEG analysis. Plasma-specific TEGs were defined as TEGs that were only detected in plasma without expression in leukocytes under any storage conditions.
Fragment characterization of plasma cfRNA
The transcript relative coverage across all transcripts was produced by RSeQC 35 with geneBody_coverage.py. The nucleotide of fragmental 3′ end was defined as the previous nucleotide at the 5′ end of each fragment. The distribution of read tags in the genome was calculated by RSeQC 35 with read_distribution.py.
Deconvolution analysis of plasma cfRNA
The relative proportion of the leukocyte subset in plasma was estimated by CIBERSORT 36 with 100 permutations and quantile normalization disabled. The same type of cells in the resting state and activated state were merged. For example, resting CD4 memory T cells and activated CD4 memory T cells are merged into CD4 memory T cells.
Statistical analysis
Statistics were performed using R tool (3.5.1). The comparison analyses of reads distribution across the genome, nucleotide frequency of the fragmental 3′ end, and leukocyte subsets were performed using the two-tailed paired t-test (for paired samples), and p-values <0.05 were considered statistically significant. Principal component analysis (PCA) plots were generated using the function prcomp(). Pearson's correlation coefficients (PCCs) were calculated by the lm() function in R.
Data availability
The data that support the findings of this study have been deposited into CNGB Sequence Archive 37 of the China National GeneBank DataBase 38 with accession number CNP0001320.
Results
The effect of blood storage on the whole-transcriptome profile of plasma cfRNA
In total, we obtained 28 cell-free transcriptomes by high-throughput sequencing to determine the effect of different storage conditions (Fig. 1). On average, each plasma sample had ∼64 million clean reads after the removal of rRNA, vault RNA, and Y RNA. We identified ∼4804 lncRNA genes, 13,701 mRNA genes, 818 miRNA genes, 412 tRNA genes, and 895 piRNA genes per sample.
To investigate the effects of storage conditions on the transcriptome profile of plasma, we performed PCA on all samples. All types of cfRNAs stored at RT for 24 hours were distinguishable from those under other conditions except cf-tRNA, while only cf-mRNA and cf-lncRNA stored at 4°C for 24 hours differed from other storage conditions (Fig. 2a). We then calculated the PCCs and performed DEG analysis between immediately processed plasma samples and samples stored for a period of time. As expected, we observed that PCCs were decreasing and the numbers of DEG were increasing during storage, no matter whether the samples were stored at RT or 4°C (Fig. 2b, c). Compared with those at 4°C, samples stored at RT showed lower PCCs and more DEGs (Fig. 2b, c).
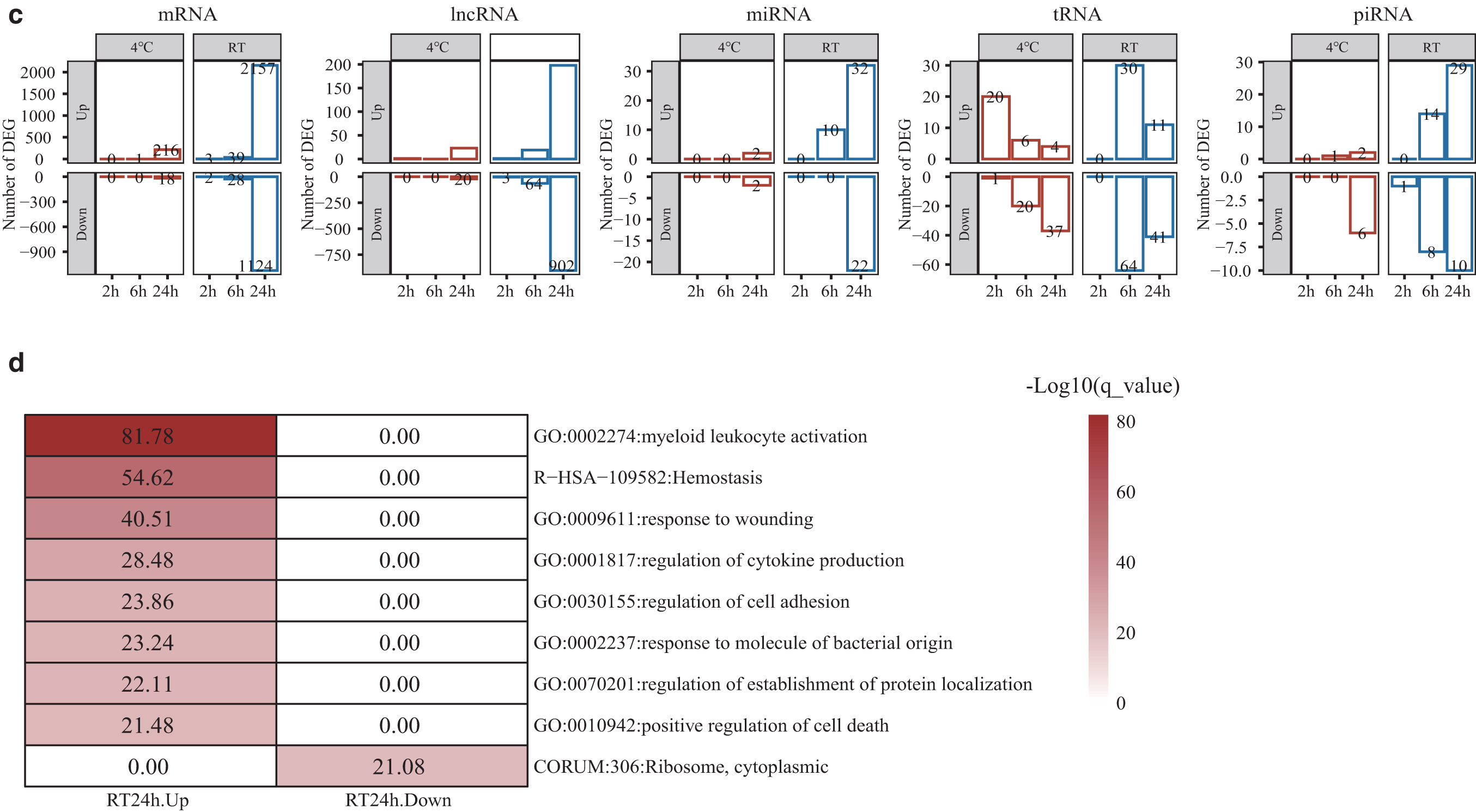
The impact of blood storage on whole-transcriptome profile of plasma cfRNA.
Cf-mRNA and cf-lncRNA showed little variation at 4°C within 6 hours and at RT within 2 hours, with high PCCs (average R2 > 0.81) and a small number of DEGs (<5) (Fig. 2b, c and Supplementary Table S1). For small RNA, 10 cf-miRNA genes (10/1253 = 0.80%) and 22 cf-piRNA genes (22/2343 = 0.94%) showed significant changes in expression when samples were stored at RT for 6 hours or at 4°C for 24 hours (Fig. 2c and Supplementary Table S1). Cf-tRNA maintained high PCCs during storage, but the numbers of differentially expressed tRNAs were fluctuant and increased to 94 (94/419 = 22.43%) at RT-6 hours (Fig. 2b, c and Supplementary Table S1).
To reveal the biological processes that may occur during blood storage, we conducted gene functional enrichment analysis on cf-mRNA DEGs. The genes with significantly increased expression levels at RT-24 hours were mainly involved in the biological processes of myeloid leukocyte activation, hemostasis, regulation of cytokine production, and positive regulation of cell death (Fig. 2d). And the genes with significantly decreased expression levels at RT-24 hours were involved in the cytoplasmic ribosomal protein complex (Fig. 2d). DEGs identified under other conditions were not enriched in any pathways.
We then examined the effects of temporary storage on platelet marker genes, and found that the expression of eight platelet marker genes significantly increased at RT-24 hours (Supplementary Fig. S1). We also explored the effects on established housekeeping genes, which are widely used as internal controls for reverse transcription polymerase chain reaction. For cf-mRNA, no housekeeping gene was significantly altered in expression level at 4°C, while four genes had significantly increased expression levels at RT-24 hours, including GUSB, ACTB, RAB7A, and HSP90AA1 (Supplementary Fig. S2a). The expression levels of CHMP2A, TUBB, C1orf43, VPS29, and PSMB2 were relatively stable at both 4°C and RT for 24 hours (Supplementary Fig. S2a). For miRNA housekeeping genes, the expression levels of hsa-miR-16-5p, hsa-miR-191-5p, and hsa-miR-25-3p were relatively stable at both 4°C and RT over time, while the hsa-miR-106a-5p expression level was reduced at RT after 24 hours storage (Supplementary Fig. S2b).
The effect of blood storage on fragment characteristics of plasma cfRNA
By assessing the variation in gene body coverage across all transcripts, we found that transcript coverages of cf-lncRNA and cf-miRNA were highly concordant for all conditions, while cf-mRNA and cf-tRNA had slight fluctuations (Fig. 3a). Subsequently, we compared the size of cfRNA fragments during storage. The results showed that most of the fragments of cf-mRNA and cf-lncRNA were <50nt at 0 hour (Supplementary Fig. S3a). However, the proportions of 96 nt cf-mRNA and cf-lncRNA (the upper limit of single-end 100 nt) as well as 53 nt cf-tRNA increased at 24 hours, especially at RT (Fig. 3b). Genes with increased expression may contribute to the increase in fragment size (Supplementary Fig. S3b). In contrast, no obvious change in fragment size was detected in cf-miRNA and cf-piRNA (Supplementary Fig. S3a). We further evaluated the nucleotide frequency of fragmental 3′ ends to infer the cfRNA cleavage by RNase A, which has a cleaving preference for pyrimidine nucleotides (T and C). As expected, a higher frequency of pyrimidine nucleotides was observed at the fragmental 3′ ends (Fig. 3c). However, the frequency of pyrimidine nucleotides was significantly decreased at RT-24 hours, suggesting that intact RNAs were released into plasma (Fig. 3c). In addition, we found that the proportion of introns increased with time, indicating the release of immature RNA from blood cells (Fig. 3d).
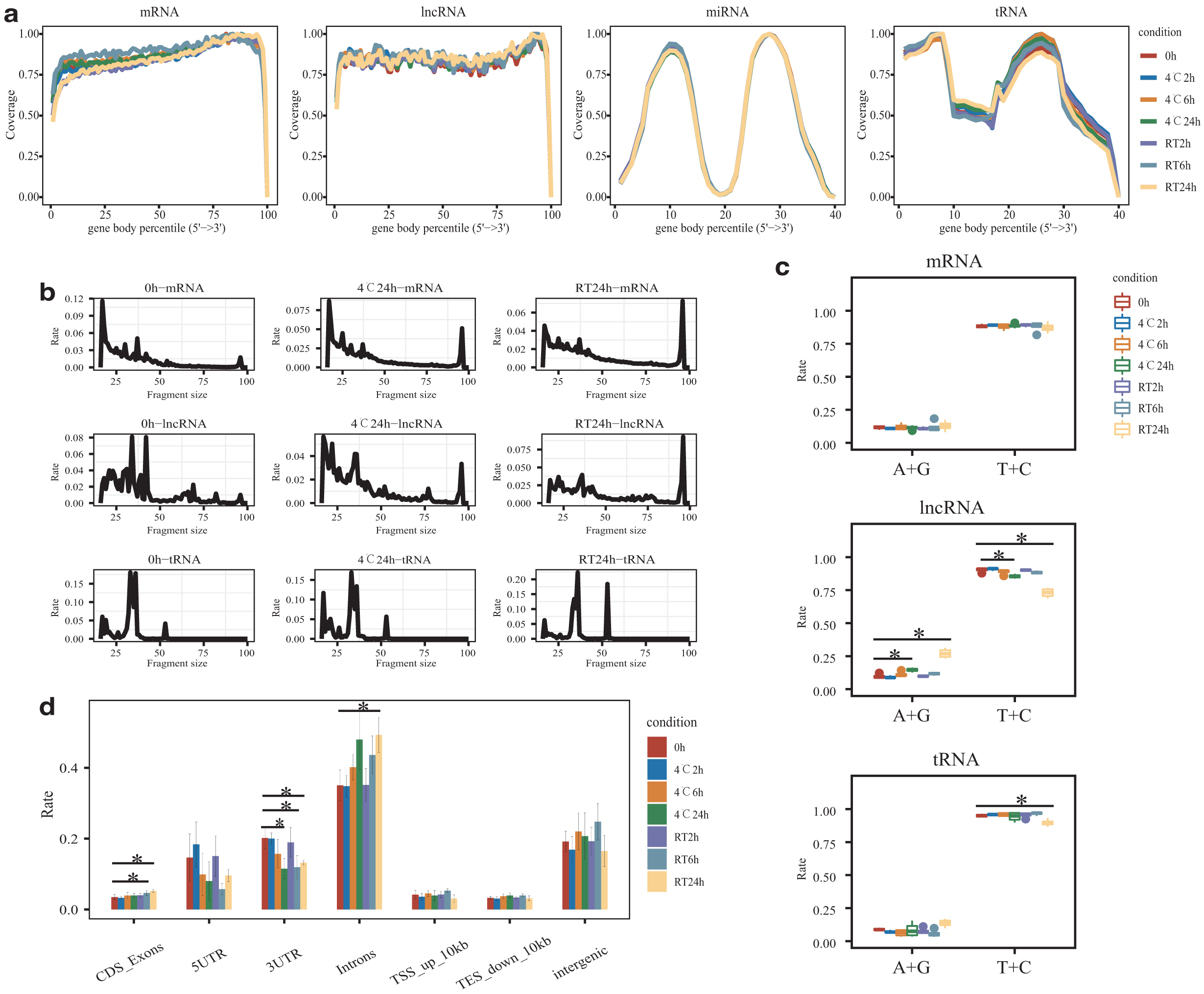
The impact of blood storage on sequence characteristics of plasma cfRNA.
The effects of leukocytes on plasma cfRNA during blood storage
To evaluate the contamination of leukocyte RNA during storage, we integrated paired leukocyte RNA data from a previous study. 23 Increasing correlations of cfRNA and leukocyte RNA were observed over time, at both RT and 4°C (Fig. 4a), indicating that the leukocyte-derived RNA contamination was increasing. Further, deconvolution on plasma cf-mRNA showed that the relative proportions of neutrophils increased from 14.3% to 61.2% (p = 0.004) at RT-24 hours (Fig. 4b), suggesting that the release of neutrophils-derived RNA increased after blood storage for 24 hours at RT.
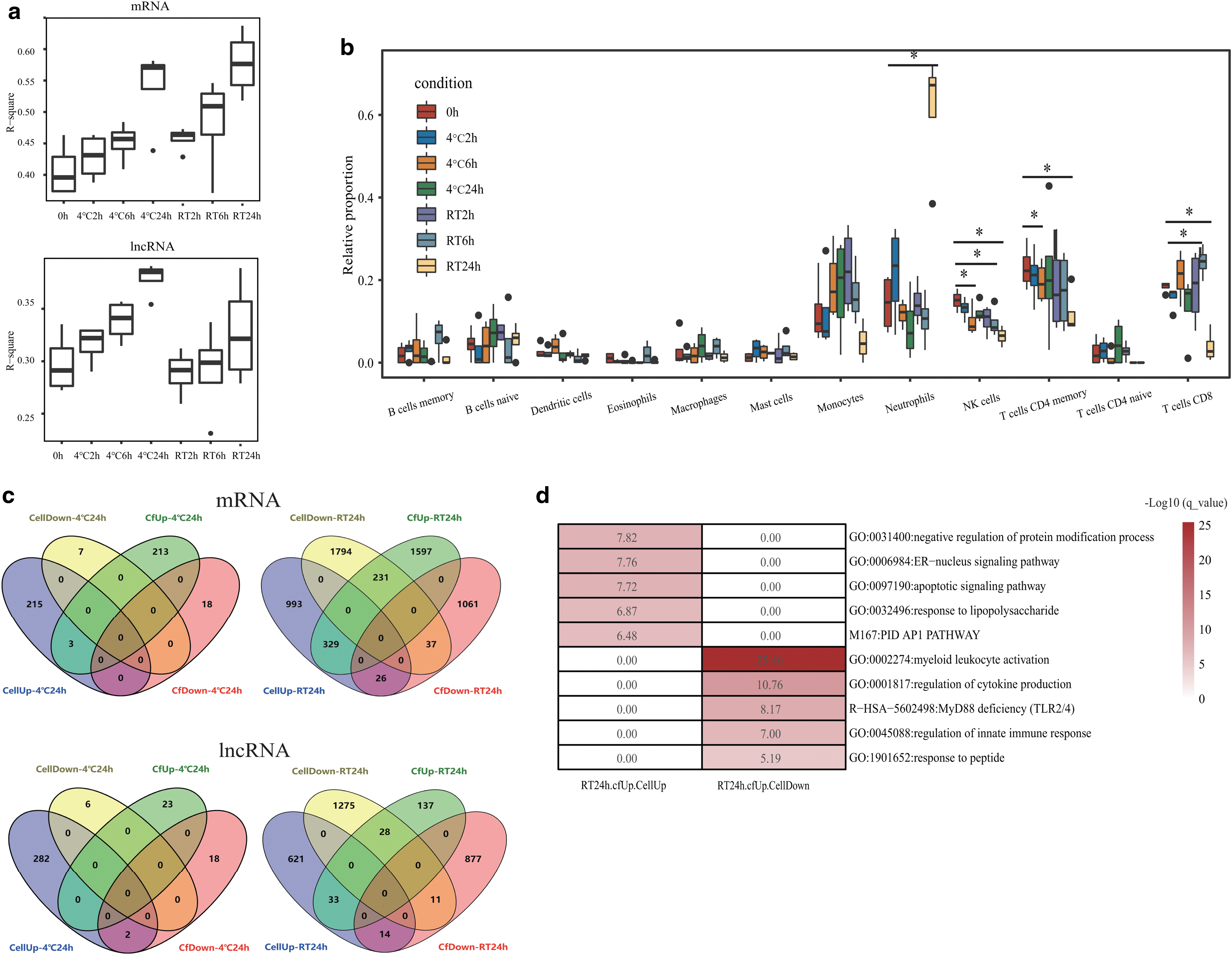
The impact of leukocyte on plasma cfRNA during blood storage.
To confirm these findings, we evaluated the DEGs shared by leukocytes and plasma under the same storage conditions (Supplementary Table S2). As a result, there were few DEGs shared by leukocytes and plasma at 4°C for 24 hours or at RT for 6 hours (Fig. 4c). However, when blood was stored at RT for 24 hours, there were 329 mRNA genes and 33 lncRNA genes that increased their expression in both plasma and leukocytes (Fig. 4c). The shared protein-coding DEGs were enriched in the apoptosis-related pathways, apoptotic and endoplasmic reticulum-nucleus signaling pathways (Fig. 4d). At the same time, the expression of 231 mRNA genes increased in plasma but decreased in leukocytes (Fig. 4c). These genes were mainly enriched in the leukocyte- and immune-related pathways (Fig. 4d). We noticed that the neutrophil activation and degranulation pathway is a member of the myeloid leukocyte activation pathway (Supplementary Table S3).
The effects of blood storage on TEGs
TEGs from 23 tissues were used to evaluate the effects of blood storage, and the highest detection rates of TEGs were lymphoid tissue and bone marrow, followed by the liver, with detection rates of 59.3%, 58.6%, and 32.2%, respectively (Fig. 5a). After removing the TEGs expressed in leukocytes, 15 TEGs were only detected in plasma (plasma-specific TEGs). Among them, 12 genes are liver specific, 1 gene is pancreas specific, 1 gene is testis specific, and 1 gene is intestine specific (Fig. 5b). The expressions of most plasma-specific TEGs gradually decreased at both 4°C and RT. Moreover, the expression level of FGA and TMIGD1 significantly decreased after 24 hours storage at 4°C and RT, respectively (Fig. 5c). We further analyzed the fragment size distribution of plasma-specific TEGs, and found that the fragment size of these genes was smaller than that of total cfRNA at 0 hour. Besides, the fragments were getting shorter with storage time (Fig. 5d), suggesting that the original cfRNA in plasma was degrading over time.

The impact of blood storage on TEGs in plasma.
Discussion
Here, we comprehensively characterized the effects of storage temperature and time on whole transcriptome and fragment characteristics of plasma cfRNA, and further investigated the contamination of blood cell-derived RNA and the degradation of the original cfRNA by integrating leukocyte RNA data. The results will help us understand the dynamics of cfRNA during storage and provide a feasible storage condition.
A few studies have evaluated the effect of blood storage on plasma cf-mRNA and cf-miRNA, and most of them focused on specific genes. Previous studies indicated that plasma cf-mRNA was relatively stable at 4°C for 24 hours 12 or at RT for 3 hours 11 by evaluating the concentration of specific genes, such as ACTB and GAPDH. Several studies believed that there were minimal effects on cf-miRNA within 24 hours of blood collection in EDTA tubes at RT by analyzing specific miRNA genes, such as miR-15b, miR-16, and miR-21,39,40 while Ward Gahlawat et al. reported that the expression level of breast cancer-associated miRNA genes (miR-148b and miR-652) significantly increased before 24 hours at RT. 18 We observed similar expression trends of these genes in our study. Moreover, we could reach an unbiased conclusion by analyzing the cfRNA landscapes during temporary storage. PCA, PCC, and DEG analyses showed that a larger change happened at RT than at 4°C and accumulated with storage time. Different types of cfRNAs underwent different paces of change, and cf-mRNA and cf-lncRNA were more unstable under the same storage conditions compared with small RNA. Cf-tRNA appears to be stable during storage as it kept high PCCs, but high proportions of differentially expressed tRNAs were observed when blood was stored at RT for 6 or 24 hours. Therefore, more samples are needed for tRNA evaluation. Blood stored at 4°C and separating plasma within 6 hours could almost retain the original cfRNA transcriptome, and are suggested for cf-RNA transcriptome study. In addition, the storage condition-sensitive genes and pathways should be cautioned in the cfRNA-related study. Preserved tubes with RNA stabilizers appear to mitigate the effects of storage temperature on RNA quality, potentially extending storage time.41–43
Cf-mRNA, cf-lncRNA, and cf-miRNA of plasma stored at RT for 24 hours were shown as distinguishable clusters in PCA, indicating that their expression profiles underwent dramatic changes. Several studies have shown that plasma RNA concentration increased over time when plasma is incubated with blood cells at RT, suggesting that RNA is released from components of blood.18,44 Consistently, the increased correlation between leukocyte and plasma RNA expression profiles, as well as the enrichment of leukocyte-associated pathways, suggests that the release of leukocyte-derived RNA is increased during storage. Neutrophils are the most abundant type of leukocytes in human. When blood was stored at RT for 24 hours, the relative proportion of neutrophil RNAs significantly increased in plasma cfRNA, but significantly decreased in leukocytes. Considering the short half-life of neutrophils, we speculate that this phenomenon might be due to the apoptosis of neutrophils.23,45,46 Meanwhile, genes with expression significantly increased in both leukocytes and plasma were enriched in the pathways associated with cellular apoptosis (apoptotic and ER-nucleus signaling pathways), supporting the apoptosis of neutrophils at RT after 24 hours storage. The release of leukocyte-derived RNA can dilute RNA from other tissues, resulting in a decrease in their relative expression, as we observed in the plasma-specific TEGs. The enrichment of platelet-associated pathways and increased expression level of platelet marker genes suggest that platelets also release platelet-specific RNA into the plasma during blood storage. In conclusion, the RNA released by leukocytes and platelets during storage contributes to the dramatic change of cfRNA transcriptome when blood was stored at RT for 24 hours. After cellular apoptosis, cellular DNA and proteins are also released into plasma during these processes.47,48 Therefore, it does not matter whether DNA, RNA, protein, or metabolites are used as biomarkers, liquid biopsy samples should be preprocessed with caution.
With the obvious contamination of leukocytes and platelets, the characteristics of plasma cfRNA fragments changed when blood was stored at RT for 24 hours. In plasma at 0 hour, we found that most cf-mRNA and cf-lncRNA fragments were <50 nt. The proportion of cf-mRNA and cf-lncRNA fragments >96 nt increased with time, and the mapping rate of introns was also significantly raised, suggesting the addition of cellular long and intact RNA fragments. Furthermore, we found that there were fewer fragments with 3′-pyrimidine after storage. As RNase A cleaves the 3′ end of unpaired C and U residues, 49 the decreased proportions of 3′-pyrimidine reflected the increase of less degraded RNA fragments. Similar changes occurred in tRNA. All these observations supported that most of the original plasma cf-mRNA and cf-lncRNA fragments are very short, while the longer mRNA and lncRNA are later released from leukocytes with the inclusion of immature mRNA. Therefore, the release of leukocyte-derived RNA affects the fragment characteristics of cfRNA.
Although other studies have demonstrated that cfRNA is protected by certain mechanisms and its concentration remains stable during storage,12,39 these studies did not demonstrate that RNA degradation is absent. It is still possible that the RNA degradation rate is slower than the release rate of leukocyte-derived RNA, resulting in an unobserved effect of RNA degradation. In this study, we considered only TEGs and excluded genes expressed in leukocytes, which reduces the effect of leukocyte-releasing RNA on degradation. Contrary to the fragment length change of overall cfRNA, plasma-specific TEG transcripts degraded to shorter fragments with storage time, implying a gradual degradation of original plasma cfRNA, which may also contribute to the decreased expression of plasma-specific TEGs. However, this finding needs to be verified in more samples. Fragment signatures of cell-free DNA, including fragment size and end motifs, can provide the nucleosome organization and nuclease content of the tissue of origin.21,50 Therefore, it had been used as a molecular marker in liquid biopsy. 51 Likewise, the fragment characteristics of cfRNA may also be used as another noninvasive marker for detecting tissue-specific lesions. In this study, only healthy individuals were included. However, the processes of leukocyte-derived RNA release or extracellular RNA degradation may be affected by disease conditions, which may result in different characteristics of changes in plasma cfRNA during blood storage.52,53 The effect of blood storage conditions on plasma cfRNA in these patients needs to be characterized in further studies.
Conclusions
In summary, plasma cfRNA undergoes decay of the original RNA and contamination of leukocyte-derived RNA, presenting changes in expression profiling and fragment characteristics during blood storage in EDTA tubes. Blood samples stored at 4°C and processed within 6 hours largely maintained the original transcriptome of cf-mRNA, cf-lncRNA, cf-miRNA, cf-Trna, and cf-piRNA. This study highlights the importance of rigorous preprocessing of samples for liquid biopsy tests and will help future clinical studies in which adequate quantification of cfRNA or fragment signature is required.
Footnotes
Acknowledgments
We appreciate the help of Dr. Sunil Kumar Sahu, Dr. Zhongzhen Liu, and Dr. Xiaoming Yin for grammar modifications in the article. We also sincerely thank the support provided by China National GeneBank.
Authors' Contributions
W-J.W. and F.C. provided conceptualization; J.Z. and S.Z. designed the methodology; J.S. and Q.Z. imparted software; X.Y., H.C., and Y.X. provided validation; X.Y. and T.W. performed formal analysis; J.S. and W-J.W. performed investigation; W-J.W. contributed to resources and project administration; J.S. and T.W. performed data curation; J.S. contributed to writing—original draft preparation and visualization; W-J.W. and Q.Z. contributed to writing—review and editing; F.C. and X.Z. provided supervision. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project is supported by the National Key Research and Development Program of China (No.2018YFC1004900), the Science, Technology and Innovation Commission of Shenzhen Municipality under the grant (Nos. JCYJ20170412152854656, JCYJ20180703093402288).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.