
Abstract
This study investigated the effect of silymarin on human sperm quality during cryopreservation. Samples were collected from 20 normospermic individuals, and each sample was divided into different concentrations of silymarin comprising the following groups: (0, 20, 100, 500, and 1000 μg/mL silymarin). Sperm quality parameters, such as plasma membrane integrity, mitochondrial membrane potential, acrosomal membrane integrity, and caspase 3 were estimated. Silymarin concentrations of 100–500 μg/mL significantly increased motility, plasma membrane integrity, and mitochondrial activity compared with the frozen control group. Acrosomal integrity was increased in the 1000 μg/mL silymarin group. Moreover, 20 and 100 μg/mL concentrations significantly decreased the percentage of caspase 3. The addition of silymarin antioxidant to the frozen medium reduced damage in the sperm after freezing and thawing. This is the first study that showed silymarin can be useful in cryopreservation of human sperm.
Introduction
Sperm cryopreservation involves freezing and storing seminal plasma fluid that contains sperm cells. 1 The cryopreservation of human sperm is a routine procedure used in assisted reproductive centers, andrology laboratories, and sperm banks. Cryopreservation is widely used to preserve fertility before treatments such as radiation and chemotherapy, in individuals with diseases that may cause testicular damage, and in azoospermic patients undergoing extraction to prevent repetitive processes.2–4
Sperm cryodamage is caused by cellular dehydration followed by osmotic shock, physical damage from intracellular formation of ice crystals, thermal shock, oxidative stress (OS), or the formation of reactive oxygen species (ROS).5,6 Increased ROS production can induce lipid peroxidation (LPO), structural changes, chromatin dysfunction, and DNA damage.7,8 Seminal plasma contains a large amount of antioxidant enzymes (superoxide dismutase [SOD] and catalase) and nonenzymatic scavengers (albumin, taurine, urate, and ascorbate) to avoid damage caused by ROS. To reduce this negative effect, freezing media are commonly supplemented with various antioxidants. 9
Silymarin, a flavonoid obtained from the extract of seeds and fruits of the milk thistle (Silybum marianum) plant, has been clinically used for over 30 years in the treatment of alcohol-related liver diseases and as an antihepatotoxic agent. 10 However, the main activity of silymarin is its strong antioxidant properties with the flavonolignans and other polyphenolic compounds it contains. 11 Studies have demonstrated that silymarin causes an increase in cellular GSH levels, increases SOD activity, and inhibits LPO. 12 We aimed to evaluate the cryodamage that occurs in the sperm cryopreservation process through parameters such as motility, plasma membrane vitality, mitochondrial activity, acrosomal integrity, and apoptosis, and to investigate the effects of silymarin, which has strong antioxidant properties, on cryodamage.
Materials and Methods
Sample collection and study group formation
For the collection of semen samples, 20 male individuals who applied to the Selçuk University Medical Faculty Hospital In vitro Fertilization Center for spermiogram analysis and whose semen parameters were classified as normospermia according to World Health Organization (WHO) 2010 criteria, 13 and voluntarily consented to study participation were recruited.
For our study, the approval of the noninvasive Clinical Research Ethics Committee (2017/80) of Selçuk University Medical Faculty was obtained. Each of the selected individuals were informed of the context of the study before the spermiogram test and signed a voluntary consent form. The individuals were all in the active reproductive period (between 20 and 40 years of age), and did not have any systemic disease, febrile illness in childhood, testicular trauma, or varicocele that affected reproductive functionality. Samples were obtained through masturbation after three to 5 days of abstinence.
Each of the fresh semen samples obtained from patients were divided into six different groups for assessment (Table 1).
Groups Created for the Study
Semen analysis
The collected semen sample was kept in an incubator at 37°C until liquefied, and was divided into six equal volumes. In the first group, the prefreeze control group, 10 μL fresh semen sample was placed on the Makler Chamber and all squares were counted under a phase-contrast microscope (CX31; Olympus) at 20 × objective to evaluate concentration and motility. 13 Individuals with a sperm count in the range of 60–120 × 106/mL were included in the study.
The motility of each spermatozoon was graded as +4 progressive motile for sperm with linear forward motility, +3 progressive motile for wide circular motility, +2 nonprogressive motile for motility without progression, and +1 immotile for nonmoving sperm. The results were evaluated as % total motility, which represents the percentage of progressive and nonprogressive motile sperm.
Eosin Y solution (Merck) was used to evaluate plasma membrane vitality. For this evaluation, 15 μL of semen sample was dropped onto a slide and 15 μL of dye was added, and covered with a coverslip. Under the phase-contrast microscope (CX31; Olympus) at 40 × objective, for each test group, five different regions were randomly selected on the entire slide. At least 200 cells were evaluated in each region and the average percentage values of the unstained sperm cells were calculated 13 for each region, and thus the viable cell ratio was calculated.
The pH (Merck ColorpHast test strips) was determined and the values were recorded. After semen analysis of the first group, acrosomal integrity, mitochondrial activity, and apoptosis assessments were evaluated. The semen samples reserved for the other groups were prepared for freezing.
Freezing and thawing of semen
To form Groups 2, 3, 4, 5, and 6, fresh semen samples were mixed with freezing medium (1067-Origio sperm freezing medium) at a 1:1 ratio in sterile cryotubes (Cryo Vial, T3082A). Silymarin dissolved in a small amount of dimethyl sulfoxide (DMSO) was added to the freezing medium at different concentrations for each group. Origio sperm freezing medium was used for dilution. This medium contains only chemically defined components, including glycerol and sucrose as the cryoprotective agents.
The added silymarin concentrations were as follows: 1000, 500, 100, and 20 μg/mL. Group 6 was the control group and no silymarin was added to its medium. The samples were immersed in liquid nitrogen (−196°C) 14 and the thawing process was initiated 72 hours later, in which they were placed in a 37°C water bath for 1 minute. After adding phosphate-buffered saline (PBS; Sigma) at a ratio of 1:1, a sample was centrifuged at 1500 rpm for 15 minutes, then the supernatant was removed from the cryoprotectant and discarded. 15
Analysis of acrosomal integrity with fluorescent microscopy
Fluorescein isothiocyanate-labeled peanut agglutinin (FITC-PNA; L7381 Sigma) was used to evaluate the acrosomal membrane integrity. 16 A 30 μL semen sample was spread on the slide and air dried. It was fixed with absolute methanol. PBS solution containing FITC-PNA (100 μL/mL) was dropped onto the sample and left at 37°C for 30 minutes, then rewashed with PBS and examined under an epifluorescence microscope (Bx51; Olympus) at 100 × magnification. Impairment of the external acrosomal membrane structure reduces the binding of lectin to the membrane.
While green fluorescence in the acrosomal cap of the sperm head indicated the presence of an undamaged acrosome, green fluorescence only in the equatorial segment, or no staining indicated that the acrosomal integrity was impaired. During the evaluation, average values were taken by counting 200 cells in 5 different randomly selected regions.
Analysis of mitochondrial activity with fluorescent microscopy
MitoTracker™ Red FM (M22425; Thermo Fisher Scientific) was used to determine mitochondrial activity. 17 One vial of MitoTracker Red FM was diluted with PBS (1:2000 ratio) and 50 μL of diluted MitoTracker solution was mixed with 1 × 106 cells in the Eppendorf tube and incubated at 37°C for 30 minutes. After incubation, the sperm sample was spread on a slide and evaluated under the epifluorescence microscope (Bx51; Olympus) at 100 × magnification.
Assessment of apoptosis with caspase 3
The Caspase 3 immunohistochemical staining method was used to determine the presence of apoptotic cells. 18 The semen sample was spread and dried on a slide with lysine coating, then fixed with absolute methanol. Anti-Caspase-3 antibody (ab13847) was used as the primary antibody. The Histostain-Plus IHC Kit, Dab, broad spectrum (859643; Invitrogen) was used for immunohistochemical staining. The procedures were done according to the kit protocol and cross-stained with Mayer's Hematoxylin (MHS1; Sigma), then closed with the mounting medium, Entellan. Sperm cells that displayed brown staining indicated Caspase 3 activation, while sperm cells that were only stained with Mayer's Hematoxylin (black arrow) indicated there was no apoptotic process. The results were calculated by counting 200 cells in 5 different regions under the light microscope (Zeiss Axio Imager.A1) at 100 × magnification.
Statistical analysis
For statistical analysis, motility, plasma membrane vitality, mitochondrial activity, acrosomal integrity, and apoptosis were compared among groups using the one-way analysis of variance test. Correlations between the sperm parameters (motility and plasma membrane vitality) of the groups were determined using Pearson's correlation test.
Results
Concentration and motility
In the sperm samples collected from 20 normospermic individuals, the mean sperm count was 85.6 ± 5 × 106 with a minimum value of 61 × 106 and a maximum of 115 × 106.
The mean percentage of total motility, comprising progressive and nonprogressive motility, was significantly higher in Group 3 and 4 compared with the other groups (Groups 2, 5, and 6; p < 0.05). There was a statistically significant difference between Group 2 (semen + freezing medium +1000 μg/mL silymarin) and Group 5 (semen + freezing medium +20 μg/mL silymarin), and Group 6 (semen + freezing medium) in terms of total motility percentages (p = 0.925, p = 0.256) (Fig. 1).

Prefreezing (Group 1 fresh semen) and freeze–thawed sperm samples, showing total motility, both progressive and nonprogressive, and plasma membrane integrity. Values for each group (n = 20) are presented as percentage totals ± standard deviation, each different letter (a–c for total motility; d–g for plasma membrane integrity) represents statistical significance between the groups (p < 0.05).
Plasma membrane vitality
The mean plasma membrane vitality was 82 ± 8.5 standard deviation (SD) % in prefreezing fresh sperm samples, and 27 ± 6 SD % after thawing. According to the comparison between groups, plasma membrane vitality was 38.6 ± 6.2 SD % in Group 4 (100 μg/mL silymarin added) and was significantly different compared with all other groups (p < 0.05) except for Group 3 (p = 0.997). There was no significant difference between plasma membrane vitality percentage values of Group 2, 5, and 6 (p = 0.784, p = 0.213) (Fig. 1).
Mitochondrial activity
In the process with sperm MitoTracker Red, the presence of intense red fluorescence in the center where the mitochondria are located indicates sperm with an active and undamaged mitochondrial membrane.
Mitochondrial activity was 78.9 ± 9.3 SD % in prefreezing sperm samples. When the groups that were subjected to cryopreservation were compared among themselves, Group 4 had significantly more mitochondrial activity compared with the other groups that had added silymarin and the control group (p < 0.05). Group 2 had no significantly different mitochondrial activity percentage compared with Group 3, 5, and 6 (p = 0.989, p = 0.997, p = 0.293, respectively).
Acrosomal integrity
Sperm that preserved acrosomal integrity displayed bright green fluorescence in the acrosomal cap, whereas sperm with damaged acrosomal membrane only displayed green fluorescence in the equatorial segment or were unstained (Fig. 2).

After cryopreservation, it was found that Group 2 had the highest rate of sperm that preserved acrosomal integrity, at a rate of 60% ± 5.9%. When the other groups were compared in terms of acrosomal integrity, there was a significant difference between all groups except for Group 4 and Group 5 (p < 0.05).
Apoptosis
When we examined the freeze–thawed sperm samples, the lowest rate of caspase-3 activation (Fig. 3) was demonstrated in Group 4 (cryopreservation with 100 μg/mL silymarin added), which was 24.4 ± 3.7 SD % and was significantly lower than all other groups that were subjected to cryopreservation (p < 0.05), except for Group 5. Regarding cryopreservation, the highest rate of caspase-3 activation was seen in the control group (Fig. 4).
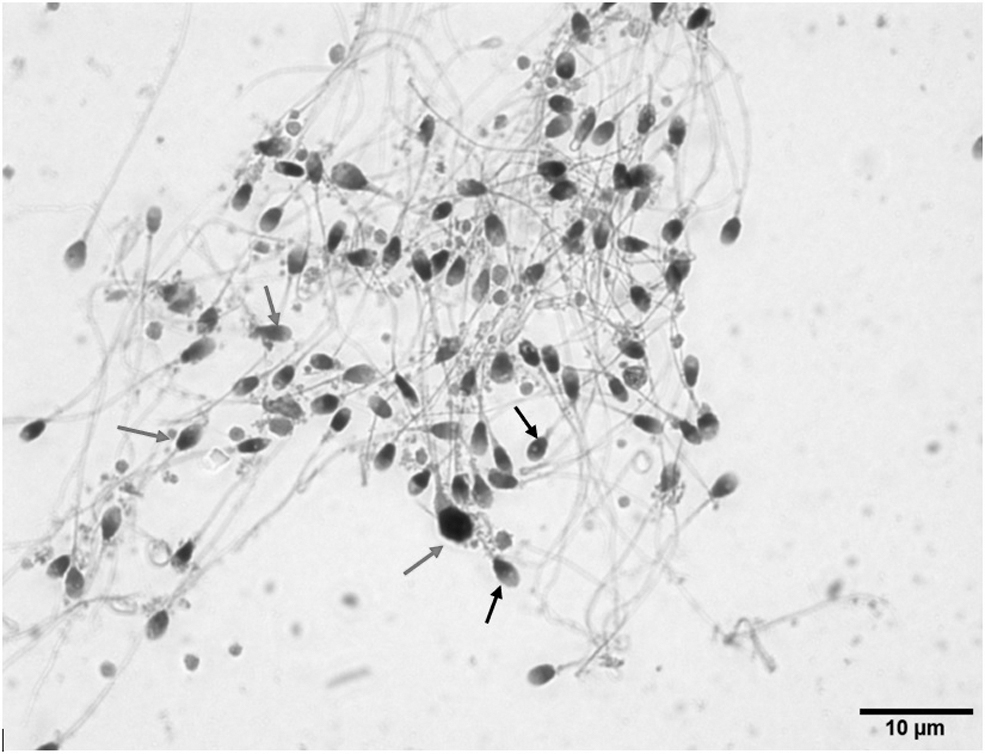
Light microscope image of immunohistochemically stained sperm. Sperm cells that displayed brown staining (light black arrow) indicated Caspase 3 activation, while sperm cells that were only stained with Mayer's Hematoxylin (dark black arrow) indicated there was no apoptotic process. 100 × Magnification.
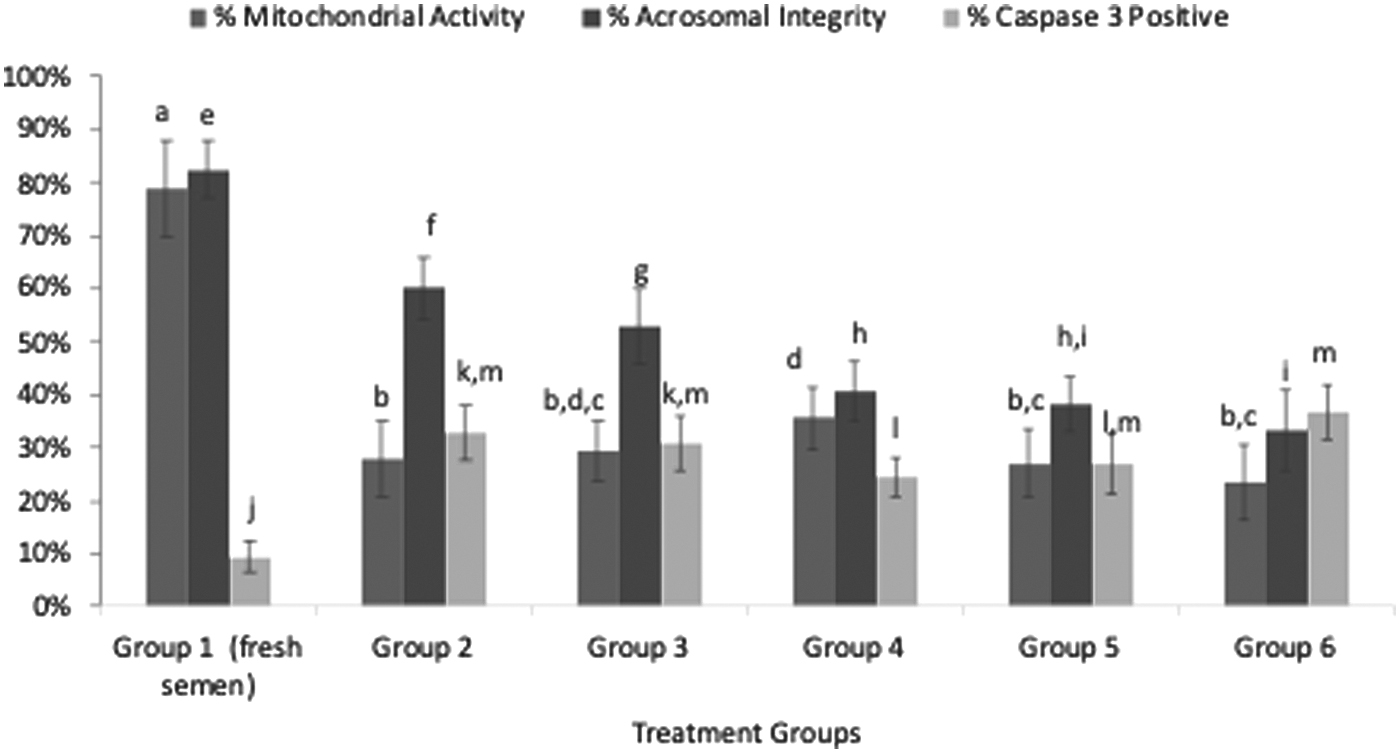
Prefreezing (Group 1 fresh semen) and freeze–thawed sperm samples, showing percentage of mitochondrial activity; acrosomal integrity; and Caspase-3 percentage. Values for each group (n = 20) are presented as percentage totals ± standard deviation, each different letter (a–d for percentage of mitochondrial activity; e–i for acrosomal integrity; j–m for Caspase-3 percentage
Analysis of sperm parameters
Correlation analysis was made to assess all parameters of the fresh sperm samples and the groups that were cryopreserved; there was a significant positive correlation between motility, plasma membrane vitality, and mitochondrial activity percentages in all groups (p < 0.01). In Group 4, in addition to the positive correlation among motility, plasma membrane vitality, and mitochondrial activity percentage values, a significant inverse correlation between acrosomal integrity and caspase-3 values (r = −0.57, p = 0.008) was also found. In Group 5, caspase-3 positively correlated with plasma membrane vitality and mitochondrial activity (r = 0.569, p = 0.09; r = 0.544, p = 0.13, respectively) (Fig. 5 and Table 2).
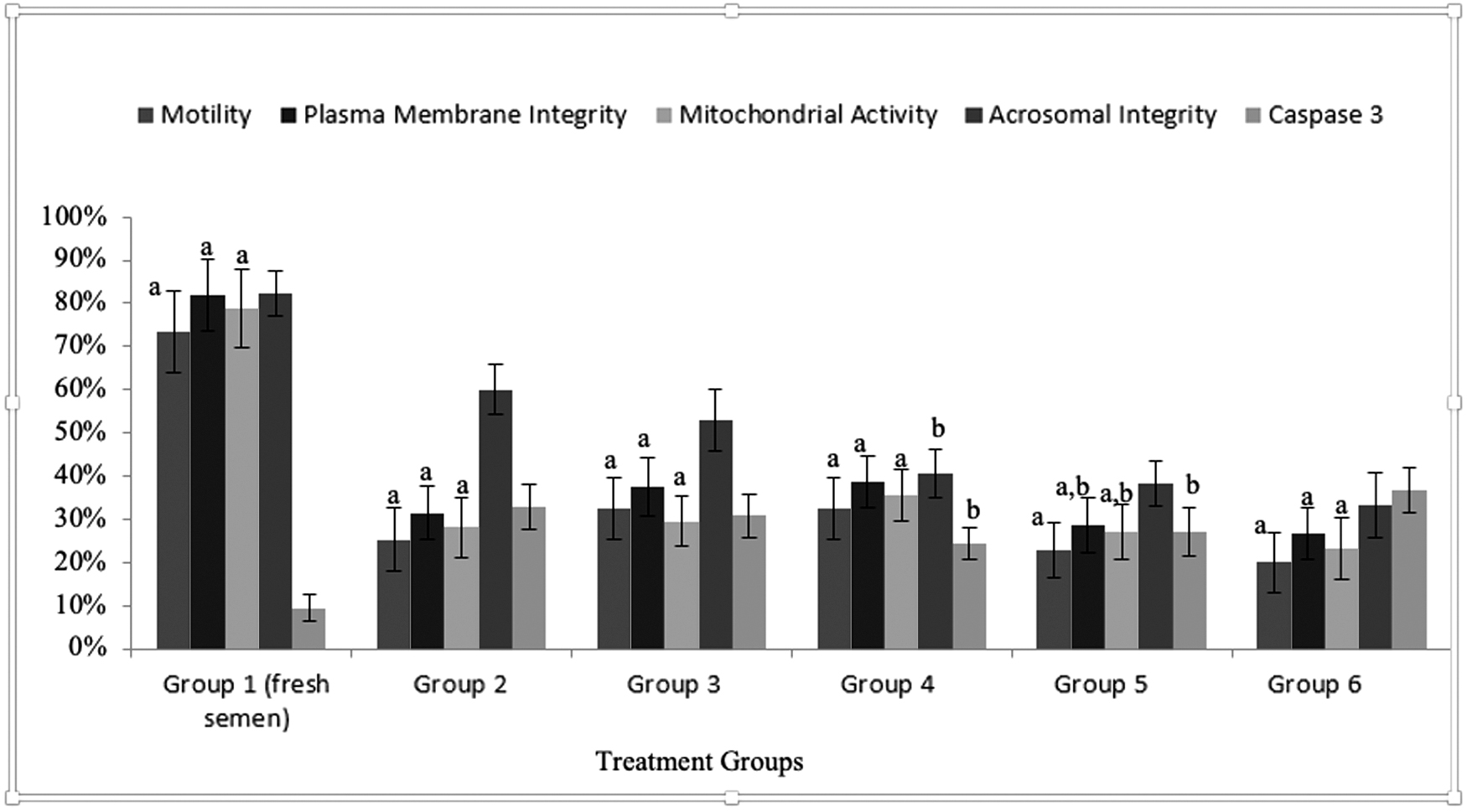
Comparison of sperm parameters according to groups. Values are presented as mean value ± standard deviation for each group (n = 20), and each different symbol (a, b) represents the statistical significance (p < 0.05) among groups.
Pearson's Correlation Coefficient Between Caspase 3 and Motility, Vitality, Mitochondrial Activity, and Acrosomal Integrity
Bold values in the table indicate statistically significant relationship between the parameters.
Discussion
According to the results of our study, adding silymarin to semen cryopreservation medium in different concentrations (20, 100, 500, 1000 μg/mL) had positive effects on sperm motility, plasma membrane vitality, mitochondrial activity, acrosomal integrity, and Caspase 3 activation after freezing and thawing.
Sperm is subjected to severe stress during both the freezing and thawing processes. 19 Osmotic and thermal stress increase ROS during cryopreservation and cause severe damage to sperm structure and function.20,21 ROS, such as hydrogen peroxide (H2O2), superoxide anions (O−2), and hydroxyl radicals (OH−), may induce apoptosis, membrane LPO, distribution of mitochondria, and DNA damage. 22 According to our results and in accordance with the literature, a statistically significant (p < 0.05) decrease in the motility, plasma membrane integrity, acrosomal integrity, and overall viability were observed in the freeze–thawed sperm.
Researchers have attempted to develop strategies to minimize various stress related freeze–thaw damages.23,24 Results of the studies showed that supplementing the sperm medium with antioxidants reduced the effect of ROS-induced damage on sperm during freezing and thawing.6,25 Some plant extracts have an antioxidant property such as Rhodiola sacra aqueous extract, 26 Resveratrol, 27 Quercetin, 28 Lycium barbarum, and Laminaria japonica plant 29 and they have been used in semen extenders to enhance post-thaw semen quality. It is known that silymarin (milk thistle-Silybum marianum extract) is a strong antioxidant that scavenges free radicals and provides potent protection against LPO. In our study, like other studies with natural antioxidants, the groups that contained added silymarin had significantly better results during cryopreservation. It was observed that the optimal concentrations for maintaining motility and plasma membrane vitality was between 100 and 500 μg/mL silymarin and demonstrated no significant difference in parameters (p < 0.05).
Our findings agreed with M Roostaei-Ali Mehr and Parisoush 30 who reported the addition of silymarin as a supplement for ram semen storage showed that sperm quality was improved. They found that sperm motility, acrosome integrity, and membrane integrity were higher in 100, 150, and 200 μg/mL silymarin than the control group. In another similar study, the addition of silymarin to sperm maintenance medium showed positive effects on bull sperm preservation, the addition of silymarin in concentrations 0.36 and 0.54 mg/mL gave post-thawing sperm motility as high as the control, while other concentrations have lower effects on motility as compared with the control. Addition of silymarin improved the percentage alive rate for the post-thawing process and was significantly higher than the rate from the control group. 31
Since cryopreservation is associated with changes in intracellular Ca2+ concentrations as well as oxidative and osmotic stress, mitochondria play a major role in the formation of cryodamage, in which an excessive decrease in mitochondrial membrane potential (early apoptosis-like phenomenon) has been detected. 32 In our study, the protective effect of silymarin on mitochondria against damage that may occur during sperm cryopreservation was investigated. According to our results, the optimal concentration range is 100–500 μg/mL, as is the case with motility, plasma membrane integrity, and mitochondrial activity; 20 μg/mL silymarin is insufficient in preventing cryodamage, whereas 1000 μg/mL silymarin suppresses cellular pathways and has a negative effect.
Eskandari and Momeni reported that silymarin could compensate for the adverse effect of sodium arsenite, which induces OS-like cryopreservation on viability, nonprogressive motility, and the mitochondrial membrane potential of ram sperm. 33 Pearson's correlation analysis demonstrated a positive correlation between sperm parameters in all groups, indicating a strong relationship among motility, plasma membrane vitality, and mitochondrial activity (p < 0.01).
In our study, it was observed that an increase in silymarin concentration provided rather good protection of acrosomal membrane integrity, contrary to motility, membrane vitality, and mitochondrial activity. While an optimal concentration range was 100–500 μg/mL for other parameters, this was not the case for acrosomal integrity. It was observed that the medium, which had 1000 μg/mL silymarin added, preserved the sperm acrosomal integrity at the highest level. However, Hu et al. reported that boar spermatozoa freezing extenders with the presence of alginate significantly improved post-thaw spermatozoa motility, plasma membrane integrity, acrosomal integrity, and mitochondrial activity compared with that of the control (p < 0.05), with various concentrations of alginate (0.2, 0.4, 0.6, 0.8, 1.0 mg/mL) added to the extender. 34
The percentages of spermatozoa classified with intact acrosomes were significantly (p < 0.05) higher in the samples cryopreserved in the presence of alginate at concentrations ranging from 0.4 to 0.8 mg/mL than 1 mg/mL. Another study reported a significantly higher (p < 0.01) percentage of progressive motility, viability, and acrosomal integrity in two
The degree of apoptosis in spermatozoa has been associated with the type of cryoprotectant that was used and the applied protocols. Although apoptosis mechanisms of human spermatozoa are not fully understood, the activation of caspases in spermatozoa are among the best indicators of cellular apoptosis. Activation of Caspase-3, an effector caspase, is one of the fundamental caspases in the apoptotic cascade and irreversibly induces cell death. As a result of our study, a significant increase in caspase-3 activation was observed after the freezing and thawing process of fresh semen (p < 0.05). Najafi et al. agreed that caspase-3 activation was markedly increased in the spermatozoa during cryopreservation compared with the control in their studies as well. 36 These findings agreed with that of some other studies: Caspases 8, 9, 1, and 3 were activated after thawing and were associated with alterations in the outer layer of the human spermatozoa plasma membrane. 37 These increases in the percentage of caspase 3 proved that the cryopreservation process triggered the apoptotic process to a significant extent.
We observed that the added silymarin, dependent on concentration, significantly decreased the percentage of caspase-3 compared with the control group. Najafi et al. reported that caspase-3 activity was found to be lower in the melatonin-supplemented group compared with that in the control group. 36 In another study, the pretreatment of ejaculated human spermatozoa with 1 mM melatonin decreased the H2O2-mediated activation of caspases 9 and 3. 38 The group with added 1000 μg/mL silymarin did not yield a higher percentage caspase-3 values than the control group indicating that silymarin did not cause any toxic effect leading the cell to apoptosis.
There are no data regarding the protective effect of silymarin on frozen human spermatozoa, to which our results could be compared. According to the parameters we evaluated, it is evident that different concentrations have a more positive effect on different parameters. Another result that was not encountered in the literature was that the group to which we added 1000 μg/mL silymarin preserved the acrosomal integrity better. This may be because the chemical structure of silymarin may impact acrosomal integrity through different mechanisms compared with other supplemented antioxidants. The results of past studies are highly variable. These differences may be attributed to multivariable parameters such as structural differences between human and animal-derived sperm, the chemical structure of the antioxidants used, the selected concentration ranges, and the cryopreservation protocols that are applied. In conclusion, we analyzed the effect of silymarin on sperm cryopreservation with five different concentrations in five different parameters.
Cryodamage during the freezing and thawing process has many different causes. While the damage occurs, the reasons affecting the parameters may be different. Although there are interrelated parameters, it can be considered as proof that cryodamage formation and silymarin affect them in different ways. We hope that our results will shed light for future studies on cryodamage and antioxidant mechanisms.
Footnotes
Acknowledgments
This article is extracted from the MSc thesis of S.Ş. The authors thank the Selçuk University Faculty Member Training Program for supporting this project.
Authors' Contributions
S.Ş.: Conceptualization (lead); and writing—original draft (lead). E.E.: Supervision (lead); conceptualization (supporting); and review and editing (lead).
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research was supported by the Selçuk University Faculty Member Training Program Coordinator with the project number 2016-ÖYP-001.