
Abstract

Introduction
Despite significant advances in sperm and embryo cryopreservation over the past decades, the ability to successfully freeze gonadal tissues and oocytes has remained limited. This is partly due to the challenge of rapid and uniform heat penetration during warming to avoid ice recrystallization particularly between −130°C and 0°C (range of ice nucleation) in large-sized frozen samples. In the domestic cat, an essential nonrodent model for human and endangered felid species, gonadal tissue and oocyte cryopreservation has been investigated for several years. Approximately 20% of morphologically normal follicles can be maintained following vitrification warming and 7 days of in vitro culture.1,2 Around 70% to 80% of testicular germ cell populations remain intact immediately following vitrification warming, recovering to fresh levels following 5 days of in vitro culture. 3
Vitrification of in vitro matured cat oocytes has resulted in the production of live offspring, 4 but equivalent success has not yet been reported after vitrification of immature cumulus–oocyte complexes. While a lot of efforts have been made to preserve the developmental potential of cryopreserved biomaterial during freezing, too few data are available about optimized warming methods to prevent ice recrystallization.
Recently, the development of ultrarapid warming using lasers has improved the survival of large frozen specimens. This approach is achieved by preloading the sample with a wavelength-absorbing material (India ink or nanoparticles such as gold nanorods). Immediately upon removal from liquid nitrogen, the sample is exposed to a laser beam, which evenly and rapidly warms the tissue through the dispersed nanoparticles. This approach has resulted in the improved viability of frozen mouse oocytes,5,6 zebrafish embryos, 7 and coral larvae. 8 In a former study in mice, 92% of laser-warmed oocytes survived (using nonpermeating India ink, with 0.70 M ethylene glycol, 0.71 M acetamide, 0.50 M sucrose, and 0.15 M Ficoll), while conventional warming yielded 76% of oocyte survival in similar vitrification conditions. 5
Another strategy for minimizing the potential harm of ice during vitrification/warming is the use of ice recrystallization inhibitors (IRIs). IRIs represent a vast family of small compounds, both synthetic and biological in nature, which are characterized by their ability to maintain ice crystals at small sizes. IRI antifreeze proteins have been supplemented to murine ovarian tissue cryopreservation to improve postwarming follicle viability, 9 while the addition of polyvinyl alcohol (controlling ice crystal formation) has also shown promise in supporting the developmental potential of vitrified bovine oocytes. 10
In the present study, our main goals were to explore the effect of laser warming versus conventional convective warming on vitrified cat gonadal tissues and oocytes, and assess the benefit of IRIs on vitrified tissues. Specifically, the objectives were to (1) set laser warming conditions for gonadal tissues in the domestic cat (Experiment 1); (2) determine the value of ovarian tissue perforation in reducing refreezing events during laser warming (Experiment 2); (3) evaluate the effect of IRIs on vitrified gonadal tissues (Experiment 3); and (4) compare the efficacy of laser warming versus conventional warming on cat ovarian (Experiment 4a) and testicular tissues (Experiment 4b), as well as on cat cumulus–oocyte complexes (Experiment 5; Fig. 1).

Overall experimental design. Experiment numbers are indicated in white circles. CW, conventional warming; IRI, ice recrystallization inhibitor; IVM, in vitro maturation; IVF, in vitro fertilization; LW, laser warming.
Methods
Domestic cat gonads were obtained from spay clinics and transported to the laboratory on ice for processing within the next 6 to 24 hours. Gonadal biopsies were collected using a standard protocol developed in our laboratory.1,3 Cumulus–oocyte complexes were released from ovarian follicles through tissue mincing with a scalpel blade at room temperature in minimum essential medium (MEM) with Hank's salt-based medium. Tissues and oocytes were then vitrified based on previous protocols1,3,11 (Table 1). Briefly, equilibrated samples were loaded onto either a Cryotop in 1–5 μL droplet of vitrification solution (for laser warming) or threaded onto a 30-gauge needle and submerged into a vitrification solution (for conventional warming), before plunging into liquid nitrogen.
Vitrification and Warming Protocols for Domestic Cat Ovarian and Testicular Tissues and Oocytes
All media solutions utilized minimum essential medium with Hank's salts (Thermo Fisher Scientific) with 20% fetal bovine serum and 0.2 mM
EG, ethylene glycol; IRI, ice recrystallization inhibitors; Me2SO, dimethyl sulfoxide; RT, room temperature.
For the latter, excess medium was removed from the samples by gently dabbing with a Kim wipe before immersion of the needle in liquid nitrogen. Cryotop samples were also exposed to 1.75 × 1011/mL gold nanorods (nanoComposix)7,8 before vitrification. Laser warming was performed with a benchtop iWeld 980 Series, 60 J, Nd:YAG infrared laser (LaserStar Technologies Corporation) with a custom-made cryolaser apparatus (Design Solutions, Inc.), 8 whereas conventionally warmed samples were directly submerged in 1 M sucrose solution (38°C). After both the warming methods, samples were washed in a reverse sucrose gradient (Table 1).
A subset of tissues (2 biopsies/cat) were stained with the Live/Dead assay immediately after warming, while the other tissues (4 biopsies/cat) were cultured for 24 hours in vitro on a 1.5% agarose gel in an MEM-based medium for 24 hours at 38.5°C and 5% CO2 1 and compared with fresh tissues (not vitrified, not cultured; 2 biopsies/cat) and control tissues (not vitrified, but cultured for 24 hours; 4 biopsies/cat). Tissues were then fixed and paraffin-embedded to evaluate apoptosis (TUNEL staining, normalized to DAPI signal) or histomorphometry (ovarian follicle density and testicular tissue tubule area through hematoxylin and eosin staining; Fig. 1). The testicular tissue from one male was also assessed for the spermatogonia/germ cell marker DDX4 (Abcam) through immunohistochemistry. 12
Oocytes were subjected to in vitro maturation (IVM) in Quinn's Blastocyst solution (CooperSurgical) (28 hours at 38.5°C and 5% CO2) 13 and then were coincubated with epididymal sperm (5 million/mL) for 18 hours. Presumptive zygotes were cultured for an additional 24 hours before fixation and chromatin staining (Hoechst 33342) to evaluate the developmental stage.
Results and Discussion
Experiment 1 was designed to set laser warming parameters. Vitrified gonadal tissues (from three female and two male cats) were warmed starting with the maximum setting (360 V, 2.5-ms exposure) and observed for evidence of overheating (bubbles or blowback of droplet; Supplementary Video S1) or underheating/ice recrystallization (flash of white in sample; Supplementary Video S2). Lower voltages (310 and 300 V) and then reduced exposure times (2.0 and 1.5 ms) were also evaluated, and the lowest possible setting that did not result in ice recrystallization was selected for each sample type. Ultimately, 300 V for 2.5-ms (ovarian) and 300 V for 1.5-ms (testicular) were selected. For oocytes (from one female), the lowest possible laser setting (300 V, 1.5 ms) was determined to be sufficient for warming, based on the criteria mentioned above.
Experiment 2 examined the effect of “perforating” the stiff tissue with a 30-gauge needle through a single stab through the center of each tissue piece before equilibration to improve cryoprotectant exposure. Perforation prevented ice recrystallization (observations in 12 biopsies) compared with nonperforated tissues (6 biopsies) and thus was applied to the subsequent ovarian experiments (Experiment 4a).
Experiment 3 explored the exposure to IRIs (small-molecule carbohydrate-based IRI-1 and IRI-2) on ovarian and testicular tissues. IRIs were provided by Dr. Robert Ben's laboratory at the University of Ottawa and selected based on prior success using these compounds in the cryopreservation of other cell and tissue systems. Specifically, the IRIs were gluconamide derivatives with concentrations previously shown to be effective at inhibiting ice recrystallization. 14 Tissue biopsies from one male and one female were incubated in either 15 mM IRI-1 or 10 mM IRI-2 for 15, 30, or 60 minutes at room temperature and were evaluated for overall cell viability (Live/Dead staining; L7011; Invitrogen) using the manufacturer's instructions. Selection of these concentrations was based on previous studies performed on coral larvae (unpublished data). The visible proportion of nonviable cells increased over incubation time (data not shown).
However, ovarian tissues exposed to IRI-1 for 15 minutes maintained slightly better viability than the other conditions and therefore were used in subsequent ovarian tissue experiments. IRI-1 or IRI-2 exposures only resulted in decreased viability in testicular biopsies and were not used in the subsequent testicular tissue experiments. Experiment 4 assessed the survival, morphology, and apoptosis in vitrified-warmed ovarian tissues (1.0–1.5 mm2 cortical sections from three individuals, Experiment 4a) and testicular tissues (0.5–1.0 mm2 from three individuals, Experiment 4b) based on optimizations mentioned above. Treatment groups included Cryotop vitrification followed by laser warming or needle immersion vitrification followed by conventional warming, and fresh tissues that did not undergo any cryopreservation.
A subset of tissues (2 biopsies/cat) were stained with the Live/Dead assay immediately after warming, while the other tissues (4 biopsies/cat) were cultured for 24 hours in vitro 15 and compared with fresh tissues (not vitrified, not cultured, 2 biopsies/cat) and control tissues (not vitrified, but cultured for 24 hours, 4 biopsies/cat). Tissues were then fixed and paraffin-embedded to evaluate apoptosis (TUNEL staining, normalized to DAPI signal) or histomorphometry (hematoxylin and eosin staining). The testicular tissue from one male was also assessed for spermatogonia/germ cell marker DDX4 (Abcam) through immunohistochemistry. 12 In Experiment 4a, no notable differences in ovarian tissue viability were observed between controls and warming treatments immediately following warming (data not shown), regardless of the presence/absence of IRIs. TUNEL staining revealed no significant differences in apoptosis between the warming methods regardless of IRI-1 presence (TUNEL-positive cells ranging from 35% to 60%; analysis of variance, GraphPad Prism 6.0; Fig. 2a).
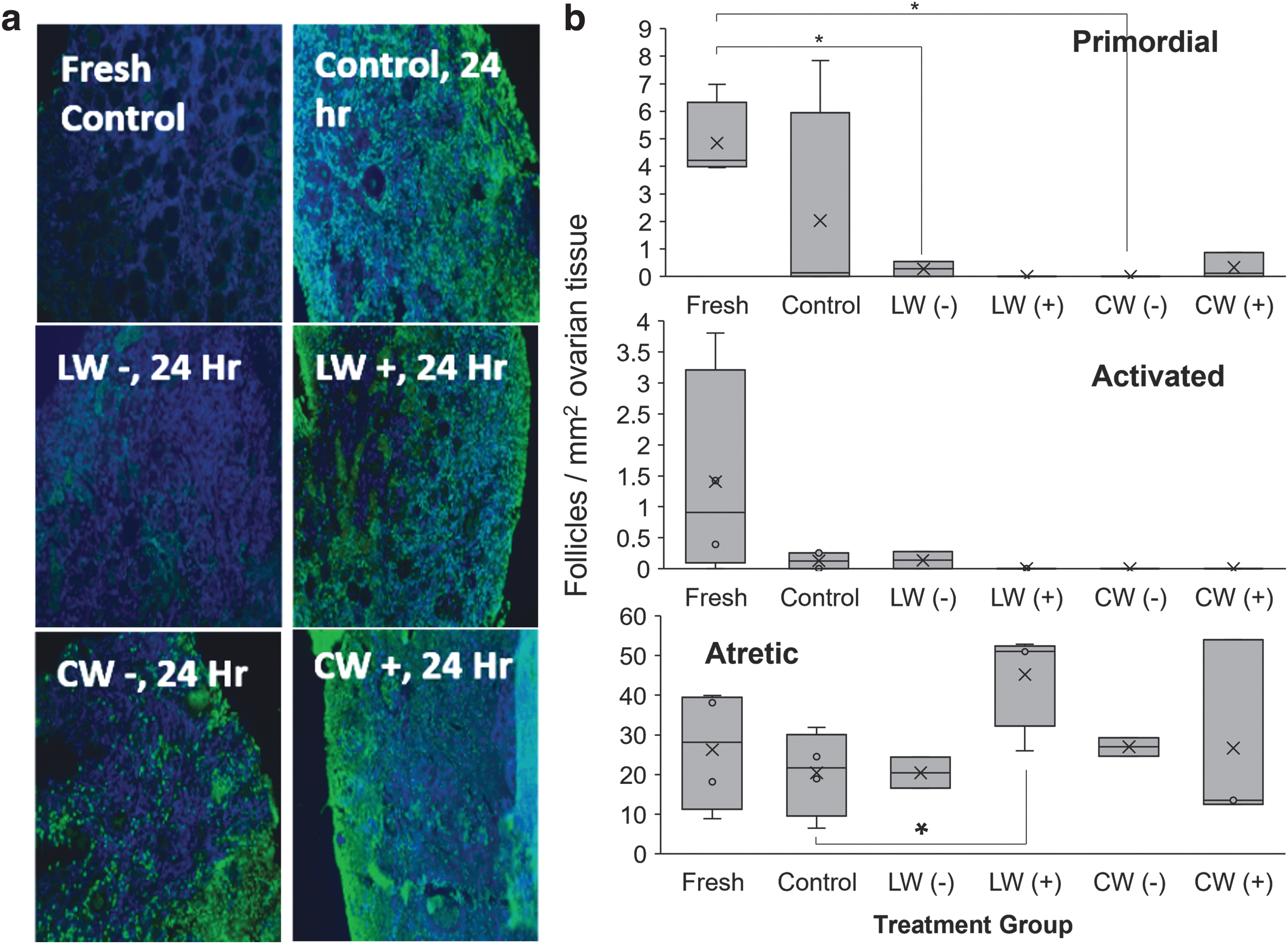
LW versus CW of vitrified ovarian tissues with (+) or without (−) IRI-1.
The density of normal primordial follicles was lower in both the vitrification treatment groups compared with fresh controls (nonparametric Wilcoxon test, R Studio) (Fig. 2b). Conventional warming and culture led to lower density of activated follicles (including transitional, primary, or secondary stages 15 ) compared with fresh controls, but were not statistically different between cultured fresh controls or laser-warmed tissues (Fig. 2b). No differences in atretic follicle densities were observed between warmed tissues (Fig. 2b).
Decreases in morphologically normal follicles, particularly following in vitro culture, have been observed in previous studies on cat ovarian tissue using needle immersion vitrification. However, one study, which used ethylene glycol, trehalose, and Me2SO in serum-free medium and solid surface vitrification, followed by warming at room temperature for 1 minute and immersion at 37°C for 30 seconds, observed no significant differences in either primordial or activated follicle densities in warmed tissues compared with fresh controls. 2 Additional research is warranted to clarify if differences observed among these studies are associated with the different vitrification strategies, vitrification media, warming rate, or combination thereof.
Moreover, there is some ongoing debate regarding the potential toxicity (potentially due to oxidative stress) of gold nanoparticles, which may vary based on particle size, concentration, and cell types. Although the specific type and concentration of gold nanoparticles used in the current study have been demonstrated to support the survival and developmental potential of coral larvae, 8 further exploration of potential toxicity or alternative heat-diffusing options (i.e., India ink) for cat gonadal tissues may be necessary.
In Experiment 4b, testicular tissues were vitrified and warmed through laser or conventional warming. No differences in viability were observed between warmed tissues (data not shown). There were no differences in the percentage of apoptotic cells between warmed tissues (TUNEL-positive cells ranging from 40% to 48%; Fig. 3a). No significant differences were observed in seminiferous tubule areas in control (cultured, not vitrified) compared with both warming groups (p > 0.05; Wilcoxon test; Fig. 3b). In the single individual subjected to spermatogonia/germ cell marker DDX4 staining, the density of DDX4+ cells within tubules was similar among treatments (Fig. 3c).
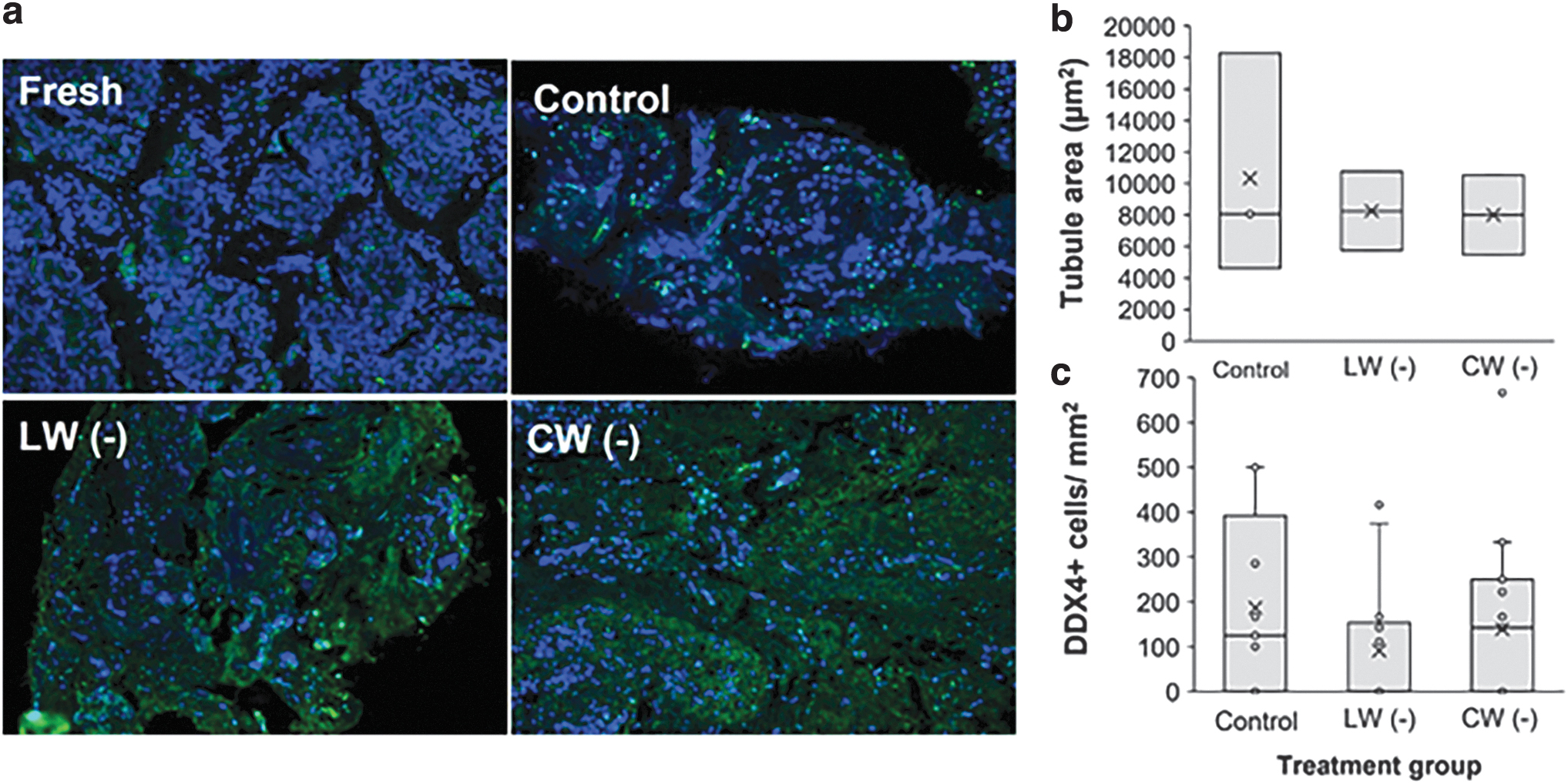
LW versus CW of vitrified testicular tissues.
The conventional warming rate applied in our study was previously compared with a short 5-second warming at 50°C before the standard sucrose gradient exposure at room temperature for vitrified domestic cat testicular tissue. 3 In that study, reduced seminiferous tubule integrity and mitochondrial membrane potential were observed in the slower 37°C warming compared with 50°C. The warming rate under the laser warming conditions in this study was roughly 40 × faster than under conventional warming; yet, no additional benefit was observed on tubule diameter and tissue architecture.
In a study in HeLa cells, it was demonstrated that the temperature range over which cells require rapid warming rates varies significantly based on cryoprotectants—starting at −120°C (i.e., immediately after entering the ice nucleation range) for a medium containing 15% Me2SO and 15% ethylene glycol, but cells in a similar vitrification solution, adding 0.5 M sucrose could be warmed briefly to −60°C without significant ice formation affecting survival. 16 For cat testicular tissues, the 15% glycerol, 15% Me2SO, 0.5 M sucrose cryoprotectant solution that we used could be tolerant to suboptimal warming rates, and would not result in significant ice recrystallization during conventional warming.
Experiment 5 evaluated the efficacy of laser warming versus conventional warming on cumulus–oocyte complexes. Oocytes (44 total oocytes from five cats) were either cultured for IVM in Quinn's blastocyst solution (CooperSurgical) (28 hours at 38.5°C and 5% CO2) 13 or vitrified (Table 1). Following either laser warming (18 oocytes) or conventional warming at 38°C (17 oocytes), oocytes were cultured for IVM and in vitro fertilization. Overall morphology and cumulus cell layers of warmed oocytes were similar among treatment groups. Most fresh oocytes matured and were fertilized (6/7). Although percentages of degeneration were high in both the laser warming (15/20) and conventional warming (16/17) oocytes, two laser-warmed oocytes matured and were fertilized (Fig. 4).
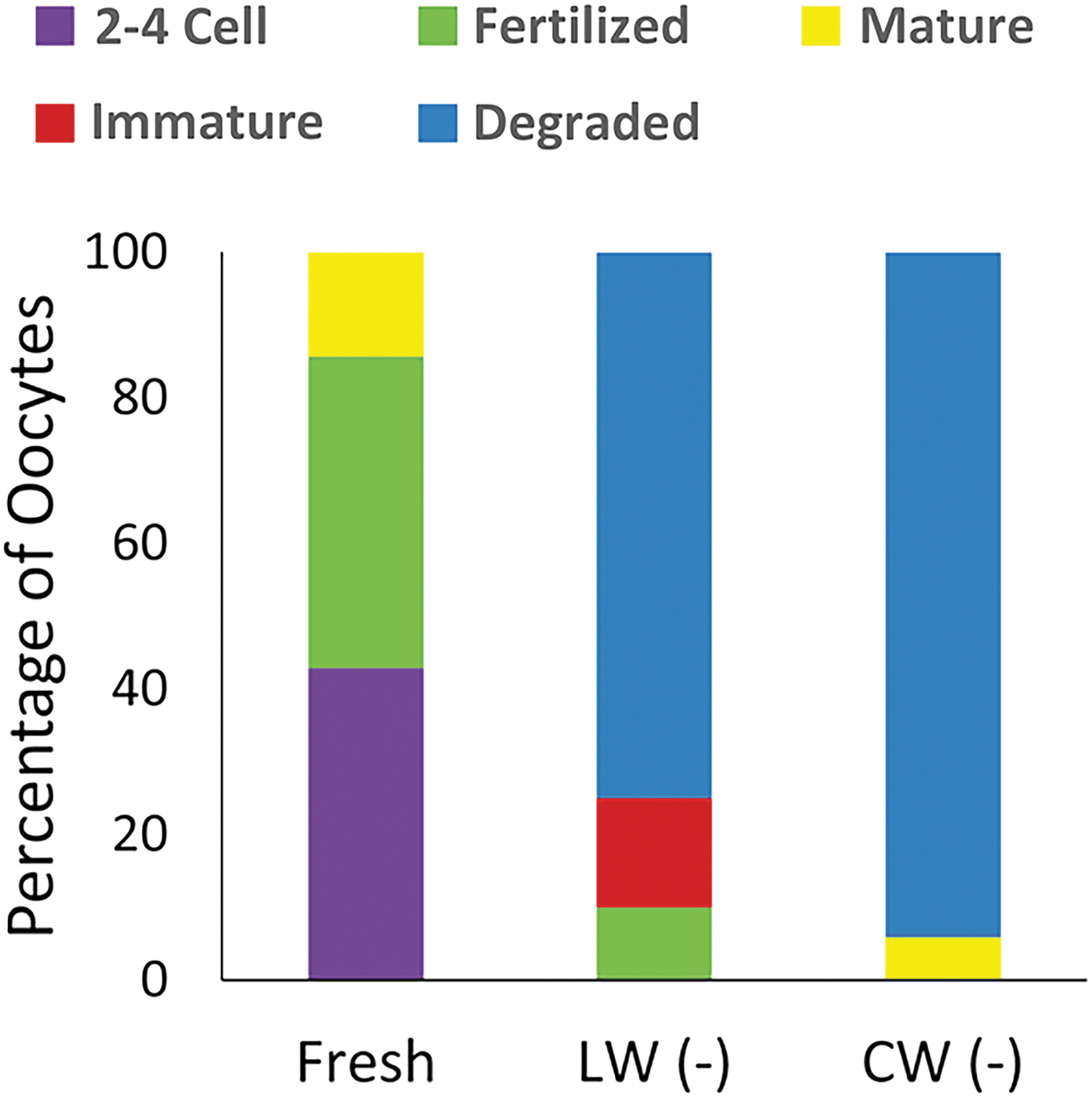
LW versus CW of vitrified cumulus–oocyte complexes in the absence of IRIs (−). Outcome of oocytes after IVM and IVF.
While these results for oocytes are promising, further studies to compare these warming strategies are needed. An extended equilibration period (6–15 minutes, as in previous reports on matured oocyte vitrification 4 ), and additional evaluation of cryoprotectant permeation, will be required to fully assess the value of laser warming for immature oocyte vitrification in the domestic cat.
In sum, we determined that 300 V for 1.5-ms was sufficient for the warming of 0.5 mm3 pieces of testicular tissue and oocytes, while 2.5-ms was preferable for 1.0 mm3 biopsies of ovarian tissue. This difference is likely due to the differences in tissue size and dispersion of seminiferous tubules in testicular tissues, allowing a better penetration of cryoprotectants and gold nanorods than in the denser ovarian stroma. Regardless of warming strategy and tissues, there was no clear advantage of IRIs at the concentrations/durations utilized here. While previous studies on IRIs demonstrated a significant increase in post-thaw integrity of other sample types including human umbilical vein endothelial cells, it is possible there may have been insufficient diffusion/penetration in the gonadal tissues used in the present study. 17
Additional assessments, including dose–response evaluations of IRIs at a more expansive set of incubation periods, are warranted to fully determine the efficacy of these compounds in these tissues. Laser warming may confer a slight advantage over conventional warming for survival of ovarian follicles, but a longer in vitro culture will be needed to further assess the developmental potential of these follicles. Laser warming of testicular tissue appeared to result in a slightly lower viability compared with conventional warming, but variation between individuals also limited the analysis of the difference between treatments. The incongruous shape of the seminiferous tubules in tissue biopsies may also render the laser warming less uniform. Lastly, laser warming of vitrified oocytes resulted in live oocytes achieving maturation and fertilization. However, further analyses are required to validate this very promising result.
Collective results report the first settings of laser warming conditions for vitrified cat gonadal tissues and oocytes. This holds potential application to both human fertility preservation and wild felid genome rescue efforts.
Footnotes
Acknowledgments
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
No funding was received for this study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.