
Abstract
The cryopreservation-thawing process of spermatozoa cells has negative impacts on their structure, function, and fertility parameters, which are known as cryoinjury. Asthenozoospermia patients are more susceptible to cryoinjury. Granulocyte-macrophage colony-stimulating factor (GM-CSF) increases sperm glucose uptake via the induction of glucose transporters, resulting in increased sperm motility. This study aimed to investigate the efficiency of GM-CSF supplementation of the cryopreservation media for semen samples of asthenoteratozoospermia patients. The study was carried out on 20 semen samples from infertile men referred to diagnosing semen analysis. To avoid subjective bias, two main sperm motility parameters, including velocity along the curvilinear path and velocity along the straight-line path were considered by the computer-assisted sperm analysis system. Afterward, each semen sample was divided into three equal aliquots and randomly assigned to one of the following groups: group I (control, freezing media only), group II (+GM-CSF, freezing medium supplemented with 2 μL/mL GM-CSF), or group III (GM-CSF added after thawing and washing). Following semen thawing, standard parameters, mitochondrial membrane potential (MMP), and the DNA Fragmentation Index were analyzed. Total sperm motility (progressive and non-progressive) improved significantly in group III samples after a 30-minute incubation with GM-CSF compared with the control group (26.5% ± 3.1% vs. 17.51% ± 2.59%). However, no differences in progressive motility or sperm morphology were found among the three thawed samples. The percentage of vitality was significantly higher in group III compared with the other two groups (28.38% ± 3.4% vs. 22.4% ± 3.08% and 22.14% ± 2.77%, respectively) (p < 0.05). JC-1 levels (a marker of MMP) were not significantly different between the examined groups (44.95% ± 8.26% vs. 36.61% ± 6.95% vs. 46.67% ± 7.7%, for control, group II, and group III, respectively) (p > 0.05). GM-CSF may be advantageous as an additive after freezing, improving total motility and viability after 30 minutes of post-thaw incubation; however, when supplied to the freezing media before cryopreservation, it is unable to protect against cryoinjury.
Introduction
Sperm cryopreservation is routinely used in assisted reproductive technology programs for patients suffering from male infertility, ejaculatory disorders, severe oligozoospermia, and cancer, as well as those with conditions resulting in testicular damage (undergoing chemotherapy or radiotherapy). 1
Despite its usefulness, cryopreservation can adversely alter sperm functional parameters, DNA fragmentation, and morphology. Alteration in metabolic status, growth factors, and cytokines, with the elevated level of oxidative stress, are the main causes of sperm dysfunction during cryopreservation.2–5 The cryoinjury can be even more severe in patients with asthenozoospermia.6,7 Asthenozoospermia is defined as <30% progressive motile spermatozoa or <42% progressive and nonprogressive motile spermatozoa in fresh semen. 8
Various approaches reduce the stress-related cryoinjury during the freeze–thaw process.9,10 Some of these strategies include the following: adding cholesterol, which enhances membrane fluidity and prevents cold shock alterations 11 ; and adding antifreeze proteins, which reduce ice nucleation and recrystallization damages. 12 In addition, oxidative stress is an important source of damages, which is minimized by neutralizing reactive oxygen species (ROS) with enzymatic, nonenzymatic, antioxidants or reductants. 9 On the contrary, the composition changes of the cryopreservation medium have been studied to achieve optimal cryopreservation results. Supplementation with antioxidants such as vitamin C, myoinositol, vitamin E, glutathione, cerium oxide nanoparticles, and cytokines has been shown to alleviate sperm cryopreservation issues.13,14
Glucose uptake and the subsequent adenosine triphosphate (ATP) synthesis are critical for sperm motility, and so, any alteration in this feature compromises sperm quality.15,16 Substrate availability and engagement of particular hexose membrane carriers are necessary for sperm to produce ATP. 17 The granulocyte-macrophage colony-stimulating factor (GM-CSF) is a glycoprotein with different functions in stimulating hematopoietic and nonhematopoietic cell proliferation, maturation, and viability. 18 Upon binding to its receptor, the phosphatidylinositol 3-kinase (PI 3-kinase)/protein kinase B (PKB) pathway is induced by which cellular glucose uptake is promoted. In addition, GM-CSF directly (via induction of the PKB/Akt pathway) and indirectly (Akt pathway) can regulate several processes involved in redistribution of glucose transporters and glucose metabolism, finally resulting in cell survival activity and also inactivation of some proapoptotic factors including BAD, caspase 9, and forkhead. 19
Interestingly, male reproductive tissues (testes, epididymis, and accessory glands) and different stages of germ cells (immature and mature spermatozoa) express GM-CSF.20,21 GM-CSF has been shown to increase sperm glucose uptake via induction of glucose transporters, resulting in increased sperm motility.22,23
On the contrary, the GM-CSF concentrations are significantly low in seminal fluid from infertile men than the normal fertile group. 24 Furthermore, many of the cytokines are unstable in semen samples stored at low temperatures (−20°C and −80°C); therefore, preserving the precise levels of the cytokines in long-term freeze storage conditions should be adjusted because during the freezing process, structural changes in many cytokines alter their quantification. 25 In animal studies, GM-CSF supplementation to freeze the medium can reverse total sperm motility, mitochondrial membrane potential (MMP), and intracellular ATP levels. 26 There is even a hypothesis that activation of the cytokine signaling pathway leads to increased glucose and vitamin C uptake, which can reverse motility in live but immotile sperms after cryopreservation. 22
Furthermore, our previous in vitro study showed that sperm media supplemented with GM-CSF can improve sperm motility via PI3K/AKT pathway activation in oligoasthenoteratozoospermia (OAT) patients. 27 Therefore, this study aimed to investigate the effects of GM-CSF supplementation on semen parameters of asthenoteratozoospermic patients during the sperm cryopreservation process.
Materials and Methods
The study was carried out on 20 semen samples from infertile men referring to the infertility clinic of Akbarabadi Hospital, Tehran, Iran, for diagnostic semen analysis. The study was approved by the Ethics Committee of Iran University of Medical Sciences (IR.IUMS.FMD.REC.1399.087). Written informed consents were obtained from all participants. Semen samples were gathered from male patients aged 24–40 years following 48–72 hours of sexual abstinence in sterile containers by masturbation. The inclusion criteria included asthenoteratozoospermia patients (defined as total sperm motility <42%, or progressive motility <30%, and sperm morphology <4%).
The exclusion criteria were necrozoospermia (a high percentage of immotile and nonviable cells) before and after sperm freezing, severe oligozoospermia, leukocytospermia (or pyospermia), azoospermia, samples from men with a history of varicocele, cryptorchidism, smoking, alcohol intake, and recent hormonal treatment (3 months).
Semen analysis, sample grouping, and freezing
A computer-assisted sperm analysis (CASA) system was used to examine the standard parameters of liquefied semen samples (30 minutes after ejaculation). To avoid subjective bias, two main sperm motility parameters including velocity along the curvilinear path (VCL) and velocity along the straight-line path (VSL) were considered by the CASA system.
Each semen sample was divided into three equal aliquots and randomly assigned to one of following groups: group I (control, freezing media), group II (+GM-CSF, freezing medium supplemented with 2 μL/mL GM-CSF; Sigma-Aldrich), or group III (supplemented with GM-CSF after thawing and semen analyzed 30 minutes later). The concentration of GM-CSF was selected by the preliminary study to find nontoxic, effective cytokine levels (2, 5, and 10 μL/mL).
Semen samples were diluted by a freezing medium (1:0.7; Cook) in a dropwise way and frozen by the method of fast vapor freezing according to the WHO laboratory manual for the examination and processing of human semen (sixth edition). 8 After a 10-minute equilibration period at room temperature, the cryovials were maintained for 15 minutes in liquid nitrogen vapor, and then plunged into liquid nitrogen (−196°C).
Samples were thawed at room temperature for 15 minutes after the cryostorage period (an average 30-day interval). The semen parameters of groups I and III were washed and analyzed using the CASA system (VCL and VSL). However, group III samples received GM-CSF in addition after thawing and washing. Semen was then incubated for 30 minutes before being subjected to standard parameter analysis.
Sperm vitality was assessed by eosine-nigrosin (E-N) staining in all the groups. For this assessment, 10 μL of the semen sample is mixed with 10 μL of E-N dye, placed on a glass slide, incubated at the room temperature for 30 seconds. At least 200 spermatozoa cells were counted under a light microscope (100 × ). Unstained cells (white sperm) were classified as the live sperms, and pink or red cells were classified as dead ones. The survival rate is obtained by dividing the number of live sperm and dead sperm that are pink or red. An overview of sample grouping and study design is shown in Figure 1.

Overview of sample grouping and study design. Created with BioRender.com.
Analysis of the MMP
Mitochondrial membrane potential (MMP) of spermatozoa cells was assessed by 5.5′, 6,6′-tetrachloro-l, la, 3, Y-tetra ethylbcnzimidazolo carbocyanine iodide (JC-1; Invitrogen) dye according to the manufacturer's instructions with flow cytometry. The spermatozoa were incubated with 2 M JC-1 dye for 10–15 minutes at 37°C, and then rinsed in phosphate-buffered saline (PBS). JC-1 aggregates and emits orange fluorescence in spermatozoa with high MMP (590 nm). In sperm with low MMP, it remains monomeric and exhibits green fluorescence (527 nm). By using software, the top right dot plot (Q2) indicates spermatozoa with high MMP, while the bottom right dot plot (Q3) represents spermatozoa with low MMP. The proportion of MMP in spermatozoa was calculated using FlowJo software. 28
Detection of DNA Fragmentation Index
The DNA Fragmentation Index (DFI) of sperm cells was analyzed using the In Situ Cell Death Detection Kit and the terminal deoxynucleotidyl transferase-mediated fluorescein-dUTP nick end labeling (TUNEL) test using the In Situ Cell Death Detection Kit (11684795910ROCHE; Roche, Mannheim, Germany) according to the manufacturer's instructions. After thawing, each semen sample was washed in PBS, resuspended in paraformaldehyde 4%, and incubated at 15°C–25°C for 60 minutes. Then the fixative was removed by washing with PBS. The sperm pellets were resuspended in 50 μL of TUNEL reaction mixture and incubated at 37°C for 60 minutes. PBS was used to wash the specimens twice. The Becton–Dickinson FACScan flow cytometer (excitation, 488 nm) in the FL-1 channel was used for measurement of green fluorescence emission. The outputs of flow cytometry were analyzed by FlowJo software and presented as the percentage of fluorescent spermatozoa.
Statistical analysis
Data were assessed by GraphPad Prism 9 software. Comparing mean values among groups was performed via one-way ANOVA with the Tukey multiple comparison post hoc tests to compare differences among the groups. The paired-sample t-test was utilized to compare before and after treatment. Statistical significance was defined as p ≤ 0.05. The data have been shown in the form of means ± SEMs.
Results
Assessment of standard sperm parameters: motility, morphology, and vitality
Compared with the control group, the total sperm motility (progressive and nonprogressive) in the group III samples after 30 minutes of incubation with GM-CSF improved significantly (p ≤ 0.05) (Fig. 2a). However, the results of progressive motility (Fig. 2a) and sperm morphology tests (Fig. 2b) revealed no difference between the three thawed samples (p > 0.05).

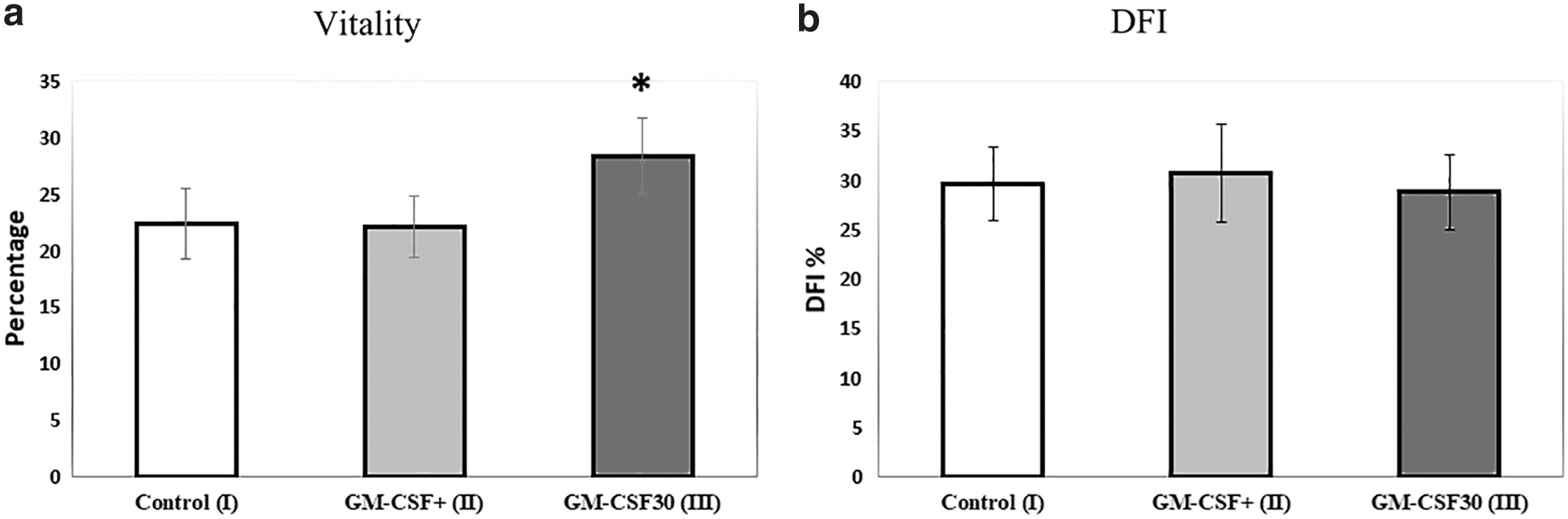
The percentage of vital sperm cells was significantly higher in the group III samples when compared with the other two groups (28.38% ± 3.4% vs. 22.4% ± 3.08% and 22.14% ± 2.77%, respectively) (p < 0.05) (Fig. 3a).
Evaluation of the effect of GM-CSF treatment on sperm DNA fragmentation
Figure 3b shows the analysis of DFI levels in the studied groups. The percentage of DFI in the examined groups did not differ before and after GM-CSF treatment (26.63% ± 3.72% vs. 30.80% ± 4.96% and 28.80% ± 3.82%, for control, group II, and group III, respectively) (p > 0.05).
Assessment of MMP
As shown in Figure 4d, the level of JC-1 (as a marker of MMP) in the studied groups was not significantly different (44.95% ± 8.26% vs. 36.61% ± 6.95% vs. 46.67% ± 7.70%, for control, group II, and group III, respectively) (p > 0.05). The mitochondria in healthy and normal cells are polarized, and JC-1 accumulates in the mitochondria as aggregates emitting red light at 590 nm. The mitochondria in cells with low MMP are depolarized, preventing JC-1 from entering the mitochondria. JC-1 remains in the cytoplasm as a monomeric form with a green fluorescence at 535 nm. 29 Differently from rhodamines and other carbocyanines, JC-1 is a ratiometric probe producing two emission peaks, which correspond to two different forms of the dye, monomer and aggregate.
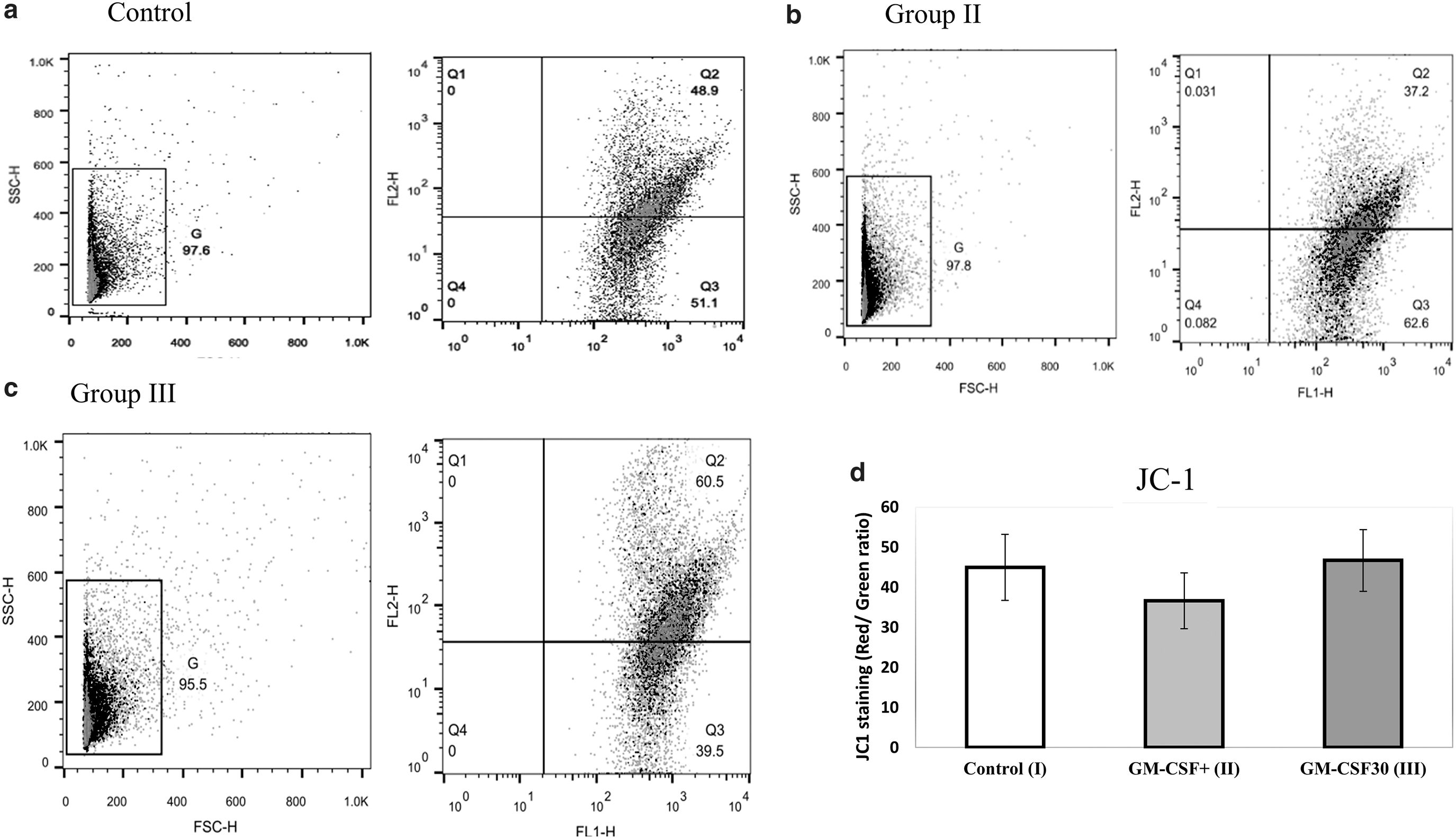
Representative dot plot of JC-1 staining for
The JC-1 monomers are predominant in depolarized mitochondria characterized by a low MMP and emit green fluorescence, whereas the JC-1 aggregates accumulate in hyperpolarized mitochondria with a high MMP and emit red fluorescence. Upon decreasing MMP, the JC-1 aggregates dissipate into monomers causing a shift from red to green fluorescence.
Discussion
The purpose of this study was to determine whether the GM-CSF supplemented to freezing media during semen freezing or after thawing impacts the post-thaw sperm quality of patients with asthenoteratozoospermia. The results of this study suggested that GM-CSF improved the total motility and vitality of spermatozoa when supplemented to washing media after the thawing process.
In addition to impairing sperm motility and vitality, the sperm freezing and thawing process has increased sperm DNA fragmentation. Cold shock, intracellular formation of ice crystal, ROS production, alteration in the semen cytokine component, and the merging of the conditions are all factors that can contribute to cryodamages. Many studies on improving the freezing components have been performed to reduce these cryodamages, for instance, by the addition of various antioxidant compounds, vitamins D, E, C, and glutathione. In general, simulating the freezing environment of sperm to normal seminal conditions can considerably preserve sperm during the freezing process.30–33 GM-CSF plays a critical role in the induction of the antioxidant mechanism via catalase activity for ROS scavenging 34 or suppressing cellular stress response genes and apoptosis pathways. 35
Motility is precisely controlled by the structural and molecular mechanisms of spermatozoa and is an essential indicator to determine male fertility. Studies have shown that GM-CSF is significantly lower in seminal plasma of infertile men than fertile ones. In addition, it contributes to the establishment and maintenance of early inflammatory events for successful pregnancy, prevents apoptosis, and enhances cell survival.24,36 We screened the compensatory effects of GM-CSF on sperm during and after freezing because it is a natural cytokine in seminal plasma and has a beneficial influence on glucose uptake by spermatozoa cells. 19
This study showed that semen GM-CSF incubation after warming for 30 minutes significantly increased the total motility of thawed sperm compared with adding it before cryopreservation. The findings of this investigation were congruent with the results of Vilanova et al.'s study, in which GM-CSF (2 nM) increased cattle sperm motility within 24 minutes of incubation. 20 In addition, human sperm motility also improved after 1 hour of incubation with GM-CSF. 27
Alteration in GM-CSF signaling causes multiple mitochondrial abnormalities, including decreased activity of tricarboxylic acid cycle and glutathione formation, and reduced glycolysis. 37 These major sources of energy are pivotal for sperm functions. 17 On the contrary, GM-CSF can exert pleiotropic effects by regulating profound functions of metabolic pathways involved in cellular proliferation, including mitochondrial turnover, metabolism, and integrity.17,37 In addition, GM-CSF receptors of spermatozoa are located on the midpiece and the tail, through which facilitating the uptake of glucose by glucose transporters. These functions with the activation of GM-CSF receptors improved motility parameters.20,23
The freezing and thawing process has a significant impact on the overall quality of the sperm. Overall, it alters acrosomal integrity, ultrastructure of mitochondria, reduces their MMP, and ATP production.38,39 The status of MMP is a potential indicator and regulator of sperm motility. 40 The assessment of this parameter is accompanied by a vitality test, which evaluates the membrane integrity of the cells, providing valuable information for the sperm's fertility potential. Although group II had a slight but not significant reduction in MMP compared with the control group, samples in group III after 30 minutes of GM-CSF incubation showed the same MMP level of control group (p > 0.05).
Furthermore, our previous study on OAT patients showed that GM-CSF boosted sperm cell MMP, with the percentage of high MMP substantially higher after a 1-hour incubation compared with the control (without incubation). As a result, GM-CSF influences sperm mitochondrial activity and has the potential to improve the process of sperm energy production. 27 Since sperm is a cell with intense energetic demands that fluctuate during its life span, medium supplementation with GM-CSF may deliver adequate energy to the cells following semen thawing by interfering with mitochondrial functions. Incubation with GM-CSF may aid under normal conditions, but it does not prevent the damage caused by freezing. However, it is effective for enhancing mobility, according to its mechanism.
Incubation with GM-CSF does not protect the cryopreserved sperm from cryoinjury if added to a freezing medium before cryopreservation. In contrast, according to a higher rate of motility and viability after 30 minutes of post-thaw incubation, it can be concluded that adding GM-CSF cytokine to the sperm washing medium after the thawing process could be effective as a treatment of sperm cryodamages.
Conclusion
It appears that GM-CSF could be beneficial as an additive after freezing as well as improving the total motility and viability after 30 minutes of post-thaw incubation.
Footnotes
Acknowledgments
All authors are very thankful to the patients for their cooperation.
Authors' Contributions
E.H.: Conceptualization (lead); writing—original draft (lead); formal analysis (lead); and writing—review and editing (equal). P.A.: Software (equal) and review and editing (equal). M.M.: Software (equal) and writing—review and editing (equal). F.A.: Methodology (lead) and writing—review and editing (equal). M.T. and F.T.K.S.: Patient recruitment and methodology—review and editing (equal). Z.Z.: Conceptualization (supporting); writing—original draft (supporting); and writing—review and editing (equal).
Confirmation Statement
Each author confirms that the research is supported by an institution that is primarily involved in education or research.
Data Access Statement
All data generated or analyzed during this study are included in the article. Additional information is available from the corresponding author on reasonable request.
Author Disclosure Statement
Funding Information
This research was supported by the Iran University of Medical Sciences (grant No: 98-4-4-14981).