
Abstract
The ability to cryopreserve oocytes without ultrastructural injury has been a concern in the development and use of methods to preserve female reproduction. The stability of the cell membrane must be preserved to reduce the damage caused by ice crystals during vitrification. One approach that has been explored is the use of static magnetic fields (SMFs), which are believed to influence cell membrane stability. In this study, the in vitro effects of SMF that range between 20–80 mT on the vitrification of mice germinal vesicle (GV) oocytes were studied. The viability and mitochondrial (Mt) membrane potential of both vitrified and nonvitrified oocytes were assessed using Trypan blue and JC1 staining. The high in vitro maturation (IVM) rate and high Mt membrane potential in metaphase II (MII) oocytes were taken into account to determine the optimal magnetic field intensity, that is, 20 mT. None of the SMF conditions resulted in intact spindles in MII oocytes. The study also explored the expression of store-operated calcium entry (Stim1, Orai1, and Ip3r) and meiosis resumption (Ccnb, Cdk) genes in GV and MII oocytes of both vitrified and control groups. The results show that the expressions of Orai1 and Ccnb genes in Vit-MII-SMF oocytes were considerably increased. However, no significant difference in Stim1 expression was observed between the groups. The Vit-MII-SMF group exhibited a significantly higher Ccnb expression compared to other groups. In vitro fertilization (IVF) was performed to evaluate the 2 pronuclear (2PN) rates. The findings demonstrated that using 20 mT SMF improved 2PN rates compared to the nonvitrified groups. This study provides a deeper understanding of the effects of moderate SMF and vitrification on the expression of calcium channel genes in GV and MII oocytes. The results suggest that applying a 20 mT SMF can help prevent cryoinjury and enhance the characteristics of vitrified-warmed oocytes.
Introduction
Oocyte vitrification offers a valuable opportunity for fertility preservation in women experiencing secondary infertility due to chemo and radiotherapy or aging. The process can impact various physiological features of oocytes, including calcium signaling. Frequently used cryoprotective agents like ethylene glycol (EG) and dimethyl sulfoxide (DMSO)1,2 have been shown to manipulate intracellular Ca2+ content. 3 Numerous genes implicated in calcium signaling pathways may experience alterations in their expression levels as a result of oocyte vitrification. 4 Specifically, the expression of inositol 1,4,5-trisphosphate receptor (IP3R) 5 which is important for calcium release from intracellular stores decreases during the vitrification of pig oocytes. The release of calcium ions from intracellular reserves and their subsequent binding to various calcium-binding proteins constitute the key steps in the calcium signaling pathway during the maturation and fertilization of oocytes. 6 In the maturation process, calcium homeostasis alters endogenous calcium oscillation and the accumulation of calcium in the endoplasmic reticulum (ER).7,8 The Ca2+ influx occurs through the store-operated Ca2+ entry (SOCE), a combination of stromal interaction molecule 1 (STIM1) and ORAI1. 7
During fertilization, upon sperm-egg fusion, sperm-specific phospholipase C zeta (PLCz) hydrolyzes phosphatidylinositol 4,5-bisphosphate to generate diacylglycerol and IP3R to release Ca2+ from the ER. 9 Long-term and high levels of calcium oscillations may cause fragmentation and apoptosis. 10 To avoid this, the pump and exchanger immediately remove any additional cytosolic Ca2+ into the extracellular environment using the plasma membrane Ca2+-ATPase (PMCA) and Na+/Ca2+ exchange. In addition, sarco-endoplasmic reticulum Ca2+-ATPases (SERCA) restore Ca2+ to the ER until it is filled. 11 In addition to the ER, the mitochondria (Mt) assist in molding the spatiotemporal patterns of Ca2+ responses (Ca2+ oscillations) and simultaneously support a variety of processes that regulate ATP levels in cells. 12 The Mt and the Ca2+-ATPase pump are important factors in regulating [Ca2+]ER during maturation and fertilization. 13 It must be remembered that fertilization and Ca2+ oscillations are critical for the completion of egg activation. Any dysfunction of Mt endangers the future of the fetus. 14
Vitrification affects the functionality of Mt. Recent research has focused on improving the quality of vitrified-warmed oocytes by minimizing ice crystal-induced deterioration of cell membranes, organelles, and structural components. Studies have shown that a static magnetic field (SMF) can alter the orientation of phospholipids, thus benefiting the membrane structure. 15 In 2017, Jasemi et al. showed that the use of a SMF improves the quality of vitrified ovaries after thawing. 16 The use of a 60 mT SMF during vitrification of mice cumulus-oocyte complex (COC) improved post-thawed oocyte functionalities such as viability, Mt potential activity, and ultrastructure17,18 In addition, the SMF affects the function of calcium channels through various mechanisms.19–22 The specific effects of SMF on the cells depend on the cell type and the intensity of the magnetic field. 23 More research is thus needed to fully understand the implications of these changes for cells, particularly regarding oocyte quality and fertility.
The main objective of this study was to examine the effects of moderate SMF on vitrification and calcium variations in oocytes. The research assessed oocyte viability, maturation rates, Mt membrane potential, and gene expression related to calcium and oocyte maturation.
Materials and Methods
This project was conducted following ethical approval from the Royan ethical committee (IR.ACECR.ROYAN.REC.1398.124) and in accordance with laboratory animal guidelines. Neodymium magnetic rings (Bio magnet-China) with varying magnetic field strengths (20.0 ± 0.1, 40.0 ± 0.1, 60.0 ± 0.1, and 80.0 ± 0.1 mT) at the center were used, as recommended by Baniasadi et.al.17,18 The study comprised six experimental groups: fresh oocytes (nVit), vitrified oocytes (Vit), vitrification of oocytes with 20 mT (Vit +20 mT), 40 mT (Vit +40 mT), 60 mT SMF (Vit +60 mT), and 80 mT (Vit +80 mT) SMF. The selection of the optimal magnetic field intensity was based on achieving a high in vitro maturation (IVM) rate, Mt membrane potential, and MII oocyte quality. Other evaluations, such as gene expression and fertilization rate, were conducted using the top-ranked magnetic field intensity. The outline of experimental procedures is presented in Figure 1.
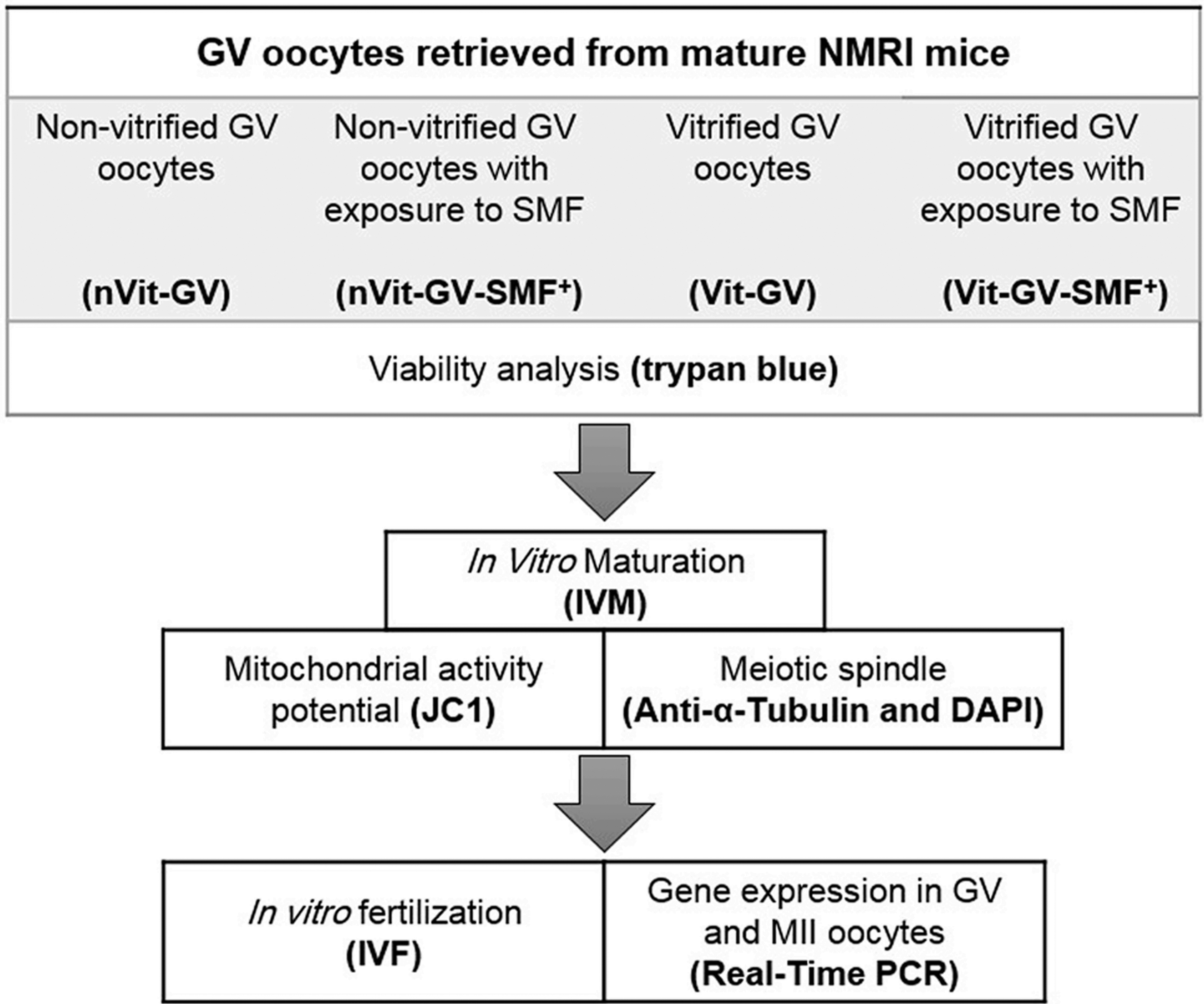
Design of experimental procedure.
Source of oocytes
Twenty-eight female NMRI (Naval Medical Research Institute) mice, aged 6–8 weeks, were used to assess the impact of a SMF on the vitrification of germinal vesicle (GV) oocytes. The mice were housed in the research institute's laboratory in Royan, Tehran, Iran, in a 12-h dark/12-h light cycle and at 40%–60% humidity and a temperature of 20°C–25°C. They had free access to food and water during the study period. To collect GV oocytes, the mice were sacrificed using cervical dislocation (CD) in accordance with the ethical guidelines of the Royan ethics committee. The ovaries were then placed in Alpha Minimum Essential Medium (α-MEM; Gibco) containing fetal bovine serum (FBS; Gibco). Denuded GV oocytes were obtained by dissecting the ovaries using insulin needles, and the oocytes were subsequently divided into groups for further procedures.
Oocyte vitrification and warming
Vitrification-warming processes were done using the Kuwayama procedure.1,2 The procedure entails rapidly freezing the oocytes to −196°C in liquid nitrogen after dehydrating them in a series of cryoprotectant solutions. The following steps are part of the vitrification protocol.
Vitrification
The vitrification process typically uses two separate cryoprotectant solutions.
The equilibrium solution (ES) was made of 7.5% EG, 7.5% DMSO, Ham's F-10 (Sigma), and 10% human serum albumin (HSA; Biotest). The vitrification solution (VS) included 15% EG, 15% DMSO, Ham's F-10, 0.5 M sucrose (Sigma-Aldrich), and 10% HSA. For vitrification, GV oocytes were washed in ES and placed in it for 5 minutes. After that, they were washed in VS for less than 1 minute at ambient temperature. They were quickly loaded on Cryotop and plunged into liquid nitrogen and kept for a day.
Warming
The warming procedure typically involves the use of a warming solution (WS). The WS was prepared by creating a sequence of solutions with gradually decreasing cryoprotectant concentrations in a base medium solution (Ham's F-10 plus 10% HSA).WS1, WS2, and WS3 had 1, 0.5, and 0.25 M sucrose, respectively. For warming, the cryotop was immediately submerged in a WS1 (37°C), and the oocytes were then washed with WS1 for less than a minute. Subsequently, they were transferred to 0.5 and 0.25 M sucrose solutions, in which they were immersed for 3 minutes each.
In the vitrification groups exposed to SMF, the end droplets of ES, VS, and WS were placed inside the center of the magnetic rings. Figure 2, generated using COMSOL Multiphysics software (version 5.2), presents the schematic representation of the droplets in the core of the 20 mT NdFeB magnet.
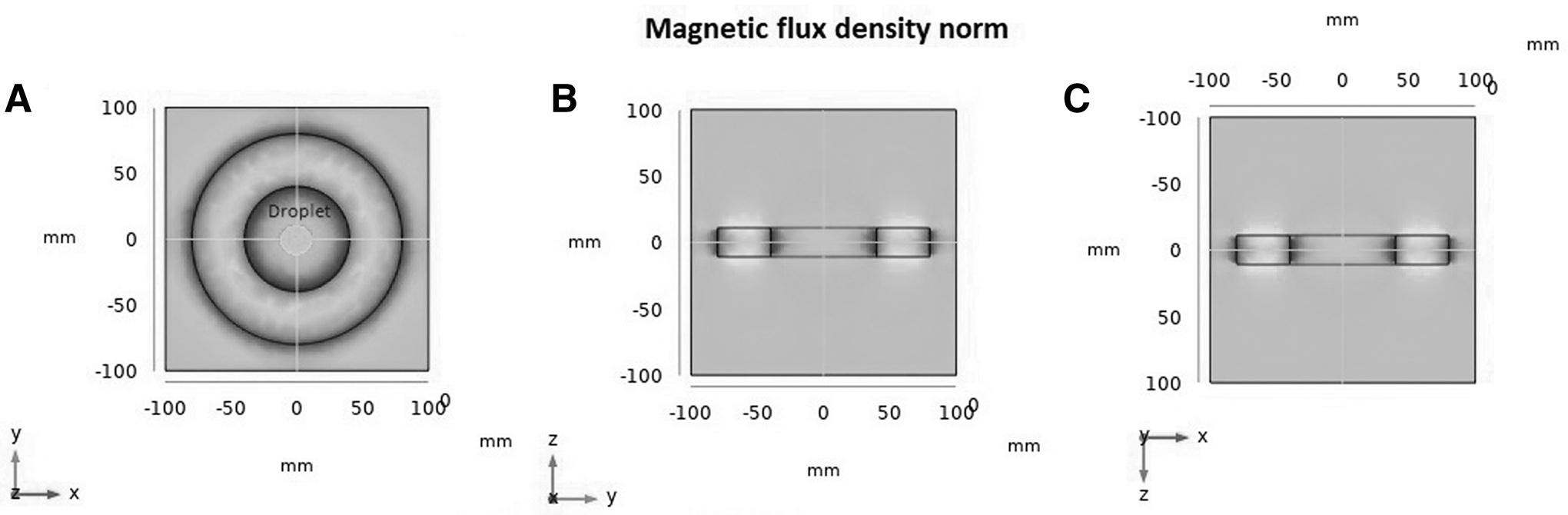
Static magnetic field configuration for 20mT magnetic ring and the placement of the drops.
Oocyte viability
Trypan blue (TBS) staining was used to differentiate live cells from dead cells. Live cells, with intact cell membranes, prevent TBS from entering their cytoplasm. Conversely, in dead cells, TBS passes through the permeable cell membrane and enters the cytoplasm [24]. Following warming, GV oocytes were stained with TBS dye and observed under a light microscope (Nikon; ECLIPSETI-U). Dead cells were characterized by a dark blue membrane, while living cells remained colorless and unstained.
IVM of GV oocytes
The vitrified-warmed GV oocytes were incubated in the α-MEM and 10% FBS medium under 5% CO2, 37, 5°C, and 90% humidity conditions for 30 minutes. Then, they were placed in a maturation medium containing α-MEM 7.5 IU/mL HCG (Merck), 100 mIU/mL follicular stimulating hormone (FSH) (Sigma-Aldrich), and 10% FBS. The MII rates were counted 18 hours later.
Mitochondrial membrane potential
Sensitive fluorescence dye JC-1 (5,5′,6,6′-tetrachloro-1,1,3,3′tetraethylbenzimid azolycarbocyanine iodide) was used to evaluate oocyte Mt membrane potential. At low concentrations, JC-1 is a monomer and emits green fluorescence (∼530 nm). At high concentrations due to membrane polarization, the dye reversibly forms JC-1 aggregates that emit at 590 nm (red color). 24 The red/green ratio indicates the Mt quality. For JC1 staining, the MII oocytes were placed in 0.02% JC1 400 mM at 37°C and 5% CO2 for 30 minutes. After washing, the oocytes were examined under a fluorescent microscope (LSCM; Leica). The red and green signals were quantified using ImageJ software (ImageJ 1.46r, Java 1.6.0_20).
Immunofluorescence for α-Tubulin protein
In all experimental groups, three or four MII oocytes were fixed in 4% paraformaldehyde (Sigma) for at least an hour at 4°C. Thereafter, they were washed in phosphate-buffered saline (PBS) (Sigma) containing 0.01% Tween 20 and permeabilized for 15 minutes at ambient temperature in PBS containing 1% Triton X-100. The membranes were thus permeable to antibodies and dyes. For blocking, the oocytes were washed and put in PBS containing 3% BSA for 1 minute. Then, the oocytes were incubated with an anti-α-Tubulin-FITC antibody (Sigma-Aldrich), and the DNA was stained with 1 g/mL DAPI (Sigma-Aldrich) for 2 minutes. The oocytes were mounted on glass microscope slides using the anti-fade mounting solution. Finally, fluorescent images were captured using a microscope (Nikon; ECLIPSETI-U).
Gene expression evaluation with real-time PCR
A Real-Time PCR method was used to investigate the expression of genes involved in calcium variations (Orai1, Stim1, and Ip3r) and resumption of meiosis (Ccnb, Cdk) in GV (The sequences of Primers has been mentioned in Table 1). The oocytes were categorized into three groups: Nonvitrified GV oocytes (nVit-GV), Vitrified GV oocytes (Vit-GV), and Vitrified GV oocytes exposed to a 20 mT SMF (Vit-GV-SMF). In addition to analyzing gene expression in GV oocytes, the expression of these genes in MII oocytes derived from nVit, Vit, and Vit +20 mT groups was investigated and denoted as nVit-MII, Vit-MII, and Vit-MII-SMF, respectively. To isolate the effects of vitrification and SMF, the nVit-GV-SMF group was included, in which fresh GV oocytes were exposed to the magnetic field for 13 minutes. The total duration that the oocytes were subjected to the field during the freezing and thawing process was 13 minutes. Furthermore, the gene expression in MII oocytes (nVit-MII-SMF) was also examined.
Sequences of Primers for Real-Time Polymerase Chain Reaction Test
The Gapdh gene was used as the housekeeping gene. The RNA Assay Micro Kit (QIAGEN) was used for RNA extraction. The RNA concentrations for the groups containing 20 GV and MII oocytes were measured with a NanoDrop 2000 spectrophotometer (Thermo Scientific) at 260/280 nm. cDNA was also synthesized with Takamed Kit (TaKaRa Bio, Inc., Shiga, Japan). To ensure their specificity, they were BLAST searched against the NCBI site. Real-time polymerase chain reaction (PCR) utilizes fluorescent reporter molecules to quantify the production of amplified products during each PCR reaction cycle. The amount of fluorescence released corresponds directly to the number of products generated in each PCR cycle, enabling quantitative measurement of the PCR response. To enhance result accuracy, each reaction was conducted twice, and four biological replications were included for each experimental group. In addition, n-template control reactions were performed for each gene. The relative expression of the genes was calculated using the ΔΔCt method, where CT represents the threshold cycle at the first significant increase in the PCR product.
In vitro fertilization
The MII oocytes in each group were subjected to the IVF procedure to evaluate the effect of SMF on fertility. The oocytes were examined morphologically using a stereomicroscope (Olympus). Normal MII oocytes were selected for the IVF. The MII oocytes that had spherical regular shapes and showed no abnormal symptoms, including torn zona-pellucida, fragmented cytoplasm, membrane damage, or degeneration, were taken to be normal. The IVF process was carried out as described by Abbasi et al. 25 Male NMRI mice who had already mated were chosen at random.
To sacrifice the male mice, CD was applied. To gather the sperm suspension, the cauda epididymides were dissected and put in 1 mL of preincubated T6 medium with 15 mg/mL bovine serum albumin (BSA; Sigma-Aldrich). For capacitation, spermatozoa were incubated for 1 hour at 37°C with 5% CO2. The average sperm count for the seven male mice was 3–5 106/mL. After capacitation, the sperm suspension was introduced to the MII oocyte in fertilization drops containing T6 medium and 15 mg/mL BSA. Six hours after insemination, the 2PN zygotes were observed under a microscope (SMZ 800; Nikon).
Statistical analyses
Statistical analyses were conducted using the SPSS software (version 22). The Kolmogorov–Smirnov test was initially used to determine whether the data were normal. ANOVA tests were used to assess the differences between the data if the data distribution was normal. Parametric data were shown as mean ± SEM. The alpha (α) level was controlled by the Tukey post hoc test when an ANOVA revealed a significant effect. In addition, data that were not normally distributed were subjected to the nonparametric Kruskal Wallis test. Data were considered statistically significant when *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. All graphs were generated using Prism software (version 7.01).
Result
Oocyte viability
TBS was used to assess the viability of GV-stage oocytes in the nVit, Vit, and Vit-SMF groups. For the vitrified groups with or without SMF, the recovered oocytes were stained. Five oocytes from each group were randomly selected and stained, and this process was repeated five times for statistical replication. No statistical differences were observed based on the TBS results.
Real MII oocyte verification
Maturation rates were assessed through five statistical replications. The results were collected after 18 hours (Fig. 3A). The observed oocytes were categorized into three groups: MII oocytes (showing rupture of the nuclear membrane, meiosis resumption, and reaching metaphase II), GV-arrested oocytes (with a visible nucleus but not resuming meiosis), and dead or degenerated oocytes. Germinal vesicle breakdown (GVBD) oocytes, characterized by the rupture and fading of the nuclear membrane, indicating the completion of meiosis I, were classified as GVBD-arrested oocytes (Fig. 3B). GV to GVBD transition (GGT) and GVBD to MII transition (GMT) rates were also calculated (Fig. 3C) to better understand the role of the SMF in oocyte vitrification and the characteristics of post-thawing oocytes.
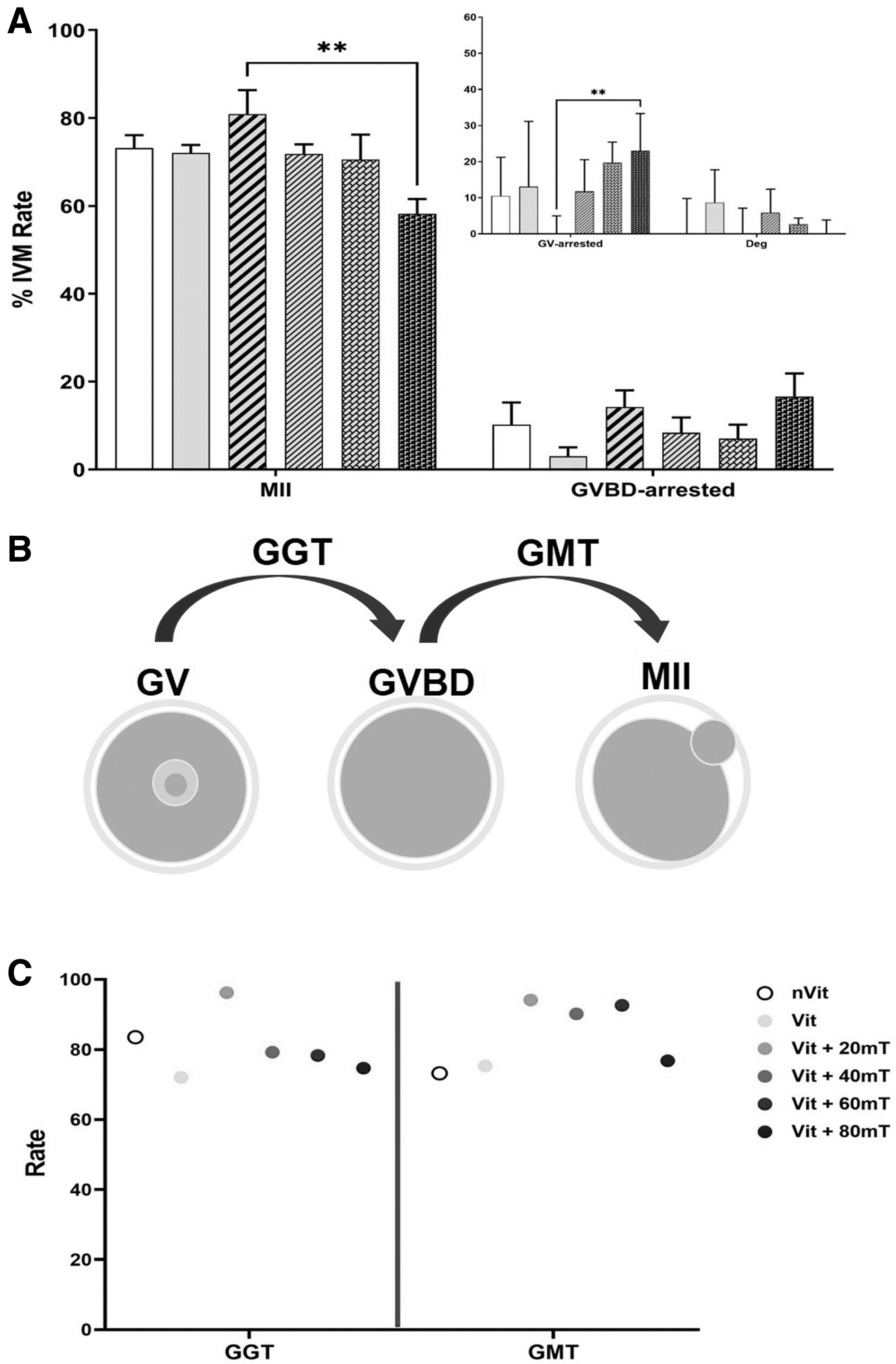
In vitro maturation of mouse GV oocytes.
As seen in Figure 3A, the differences in maturation rates between Vit (72.07% ± 2.96%), nVit (73.13% ± 2.96%), and Vit +20 mT (80.92% ± 5.44%) groups were not statistically significant. However, the maturation rate of the Vit +80 mT group (58.22% ± 3.36%) was significantly different from that of the Vit +20 mT group. Moreover, no significant difference was observed in GVBD-arrested and degeneration rates between experimental groups. The GV-arrested rate in Vit +20 mT decreased compared to the Vit +80 mT group. Furthermore, Figure 3C shows that the GGT value was lower for Vit +80 mT than for Vit +60 mT. Maximum GGT and GMT were obtained when the 20 mT SMF was used during vitrification.
Mitochondrial potential activity
The JC1 experiment was conducted with five statistical replications. Figure 4 illustrates the red and green signals for MII oocytes associated with the nVit, Vit, Vit +20 mT, Vit +40 mT, Vit +60 mT, and Vit +80 mT groups. The statistical results are presented in Figure 5. According to Figure 5, the mean red-to-green signal in the Vit group (47.77% ± 4.24%) significantly decreased compared to the nVit group (70.18% ± 4.84%). Moreover, the value in the Vit +80 mT group (32.86% ± 6.90%) was reduced compared to the nVit and Vit +20 mT (61.51 ± 7.06) groups. No statistical differences were observed between the Vit +40 mT (53.41% ± 3.18%), Vit +60 mT (50.38% ± 3.09%), and Vit +20 mT groups and the nVit group.
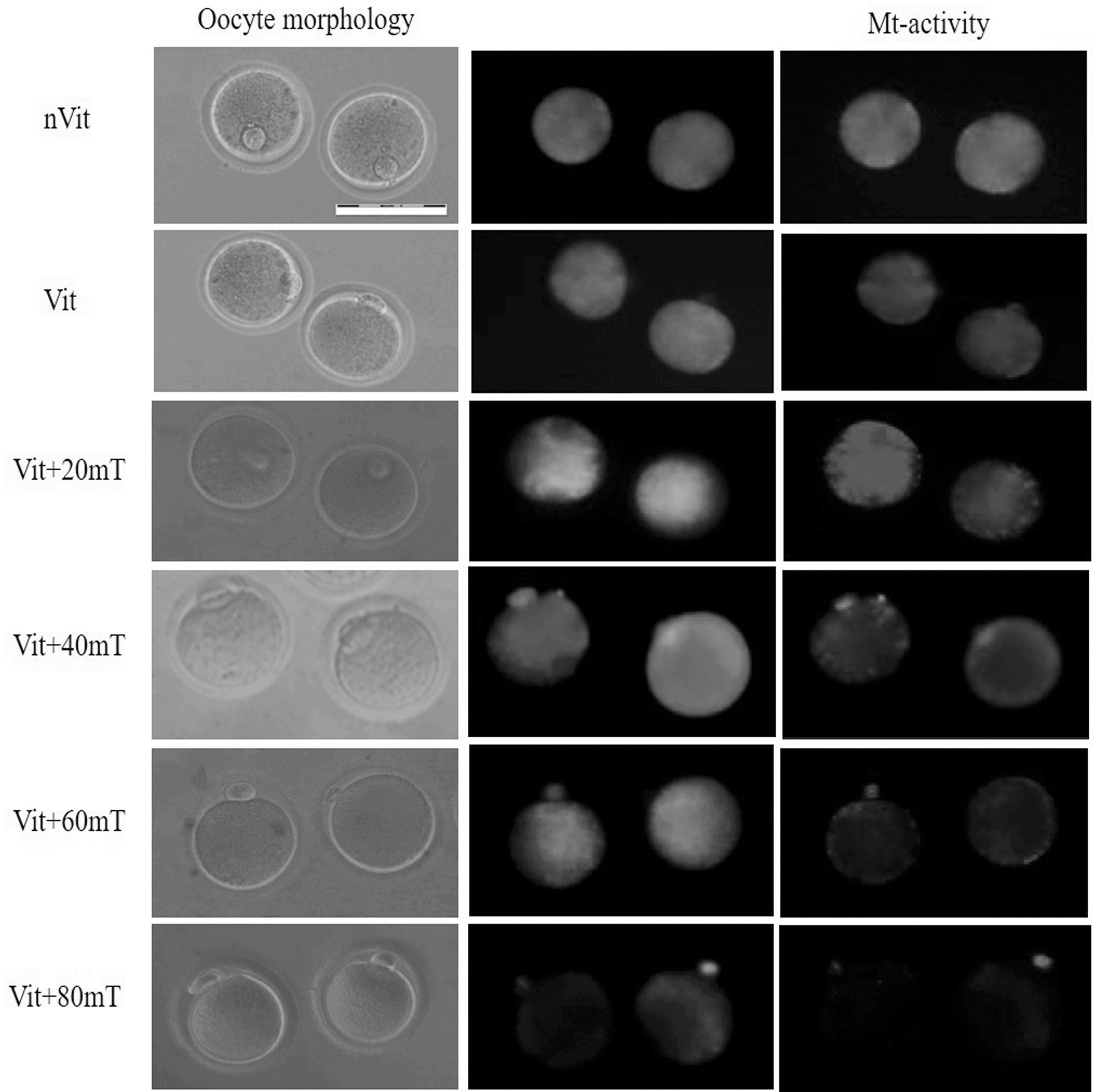
Mitochondrial membrane potential staining for mouse MII oocytes in nVit and Vit-SMF groups. Scale bar = 50 μm. MII, metaphase II.

The mean of red to green signals in MII oocytes that is, data are shown as mean ± SEM. Differences were statistically taken to account as significant at *p < 0.05, **p < 0.01, and ****p < 0.0001.
Mitotic spindle and positioning of the chromosomes
Anti-α-Tubulin and DAPI staining were used to investigate the effect of SMF on the meiotic spindle and oocyte chromosomes (Fig. 6). Spindle staining revealed that an SMF intensity of 60 mT altered the orientation of the spindle, while other intensities had no significant impact on the meiotic spindles.

Microtubule spindle assessment in mouse MII oocytes for nVit, Vit, and Vit +SMF groups at different magnetic field strengths. Green signal and blue signals indicate anti-α-tubulin-FITC antibody and DAPI staining, respectively. Scale bar, 20 mm.
Gene expression level (real-time PCR)
For gene expression analysis, five statistical replications were performed. Figure 7 shows the outcomes. The expression level of the Stim1 gene in the nVit-GV-SMF, Vit-GV, and Vit-GV-SMF groups showed a nonsignificant increase compared to the nVit-GV. For MII oocytes, the increased expression of Stim1 in the Vit-MII-SMF group compared to nVit-MII, nVit-MII-SMF, and Vit-MII groups was not meaningful. The Orai1 expression in Vit-MII-SMF (2.93 ± 0.61) increased significantly compared to nVit-MII (0.22 ± 0.06), nVit-MII-SMF (0.10 ± 0.02), and Vit-MII (0.89 ± 0.09). The expression level of Cdk in the Vit-MII (6.46 ± 0.47) increased significantly compared to nVit-GV (0.66 ± 0.10), nVit-GV-SMF (0.21 ± 0.05), Vit-GV (0.69 ± 0.05), Vit-GV-SMF (0.35 ± 0.19), nVit-MII (0.06 ± 0.02), and Vit-MII-SMF (1.11 ± 0.43).
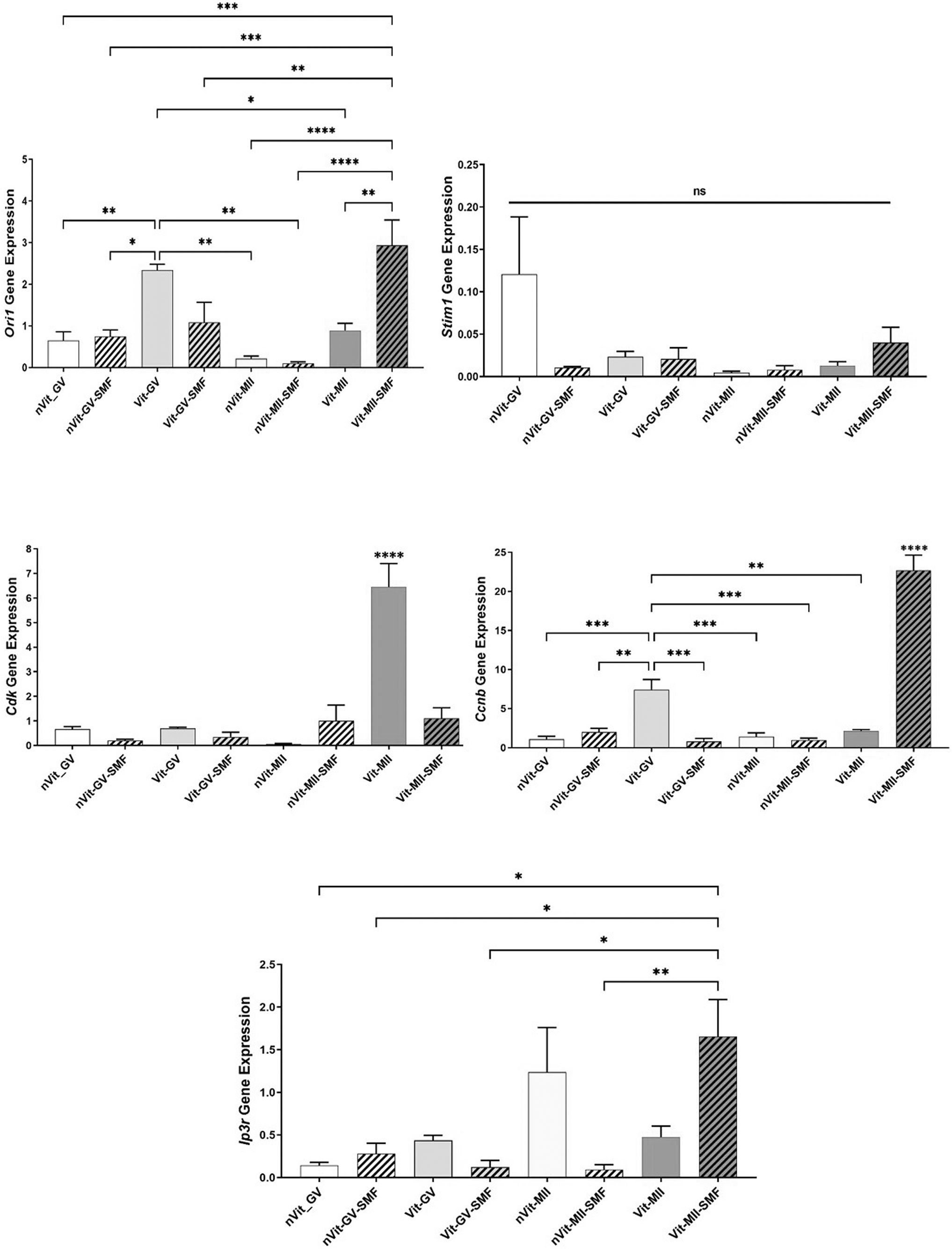
Analysis of Orai1, Stim1, Cdk, Ccnb, and Ip3r gene expression in GV and MII oocytes with real-time PCR. Data are presented as mean ± SEM. Asterisks mean significant difference (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001). The ns indicates not significant.
The Ccnb expression was also significantly increased in the Vit-MII-SMF (22.68 ± 1.96) in comparison to nVit-GV (1.07 ± 0.38), nVit-GV-SMF (1.99 ± 0.49), Vit-GV (7.44 ± 1.29), Vit-GV-SMF (0.79 ± 0.40), nVit-MII (1.41 ± 0.48), nVit-MII-SMF (0.98 ± 0.12), and Vit-MII (2.14 ± 0.09). Compared to all experimental groups except Vit-MII-SMF, Ccnb expression in the Vit-GV was significantly increased. The Ip3r expression in Vit-MII-SMF (1.66 ± 0.43) increased significantly compared to nVit-GV (0.14 ± 0.03), nVit-GV-SMF (0.28 ± 0.12), Vit-GV-SMF (0.13 ± 0.07), and nVit-MII-SMF (0.09 ± 0.03).
In vitro fertilization
For in vitro fertilization, seven statistical replications were used. For the IVF process, the derived MII oocytes belonging to nVit, Vit, and Vit +20 mT (optimal intensity of SMF) groups were considered. The fertilization rate was calculated as 2PNs/MIIs. As seen in Table 2, the number of 2PNs in the Vit +20mT (85.87% ± 3.33) increased significantly compared to the nVit (49.14% ± 6.74) and Vit (63.85% ± 5.14) groups. However, there was no meaningful difference in the 2PN rate between nVit and other groups.
In Vitro Fertilization in Nonvitrified, Vitrified, and Vitrified Oocytes Treated with 20 mT Static Magnetic Fields
Mentions to significant differences with the both other groups.
Differences were statistically taken to account for significance at * p < 0.05.
IVF, in vitro fertilization.
Discussion
Prior studies have already explored the use of 60 mT SMF in COC vitrification. This work offers the first independent validation of the positive effects of moderate 20–80 mT SMF on GV oocyte cryopreservation. The findings indicate an improvement in the quality of vitrified-warmed oocytes after exposure to 20 mT SMF.
The membrane of the oocyte is full of calcium channels, including CRAC (ORAI + STIM), T-type Ca2+ channel, TRPV3, TRPM7, and voltage-activated K+ channels. 26 According to studies by Rosen, SMF causes changes in the calcium channels in the cell membrane, as well as cytoplasmic organelles. 19 In this study, the use of a 20 mT SMF resulted in an increased maturation rate in GV oocytes compared to other groups. The magnetic field likely influenced calcium oscillations by modifying the calcium channels in the cell membrane and organelle membranes, thereby enhancing oocyte maturation.
Mitochondria, the crucial organelles within the cell, not only influence oocyte maturation and development but also play a significant role in fertilization and embryo development. The metabolic activities of mitochondria during cytoplasmic and nuclear maturation are undeniable. Therefore, any dysfunction in mitochondrial structure and function can lead to impaired maturation or incomplete embryo development. 24 Lei et al. reported that vitrified mouse oocytes exhibit a significant reduction in red-to-green fluorescence signals compared to nonvitrified oocytes. 27 Vitrification changes the Mt activity due to an increase in reactive oxygen species.28,29
In this study, the Mt activity in the Vit group was decreased compared to the nVit group. However, using SMF with intensities of 20–60 mT improved the Mt activity. Research by Bernabò et al. demonstrated that exposure of granulosa cells to 2 mT SMF decreased the potential of Mt membranes and increased the entry of sodium and calcium into the cell. 30 It seems that SMF affects Mt activity through sodium and calcium voltage channels, which in turn alter cell metabolism and energy. 31 The ER plays a role alongside mitochondria in forming calcium oscillations, which have an impact on the outer Mt matrix and the maintenance of ATP levels in the cell. Moreover, the relationship between the ER and IP3 receptor, as well as calcium release from the ER, contribute to preserving the stability of this ion in the mitochondria. 13
Following an increase in cytoplasmic calcium levels, Ip3r expression rises during maturation. An opening of channels in the ER promotes these modifications. 32 Because the expression of this gene dramatically increased in Vit-SMF group MII oocytes, it appears that the SMF impacts the ER membrane and causes an increase in calcium oscillations. Lequarre et al., Quetglas et al., and Kumar et al. have shown that the level of expression of the Ccnb does not alter during maturation.33–35 The outcomes of the current study are consistent with these observations. However, Gomez et al. have reported varied outcomes regarding increases in Ccnb expression during oocyte maturation. 36 Zand et al. indicated that the expression level of the Cdk in GV oocytes is low. 37 In addition, in line with the observations of Moore et al., the expression level of Cdk does not alter during maturation because the amount of mRNA for this gene does not change throughout this period. 38 During oocyte maturation from the GV to the MII stage, the Cdk expression level gradually changed in all experimental groups except Vit-MII.
A study by Wang et al. indicates that the expression of Stim1 remains unchanged from GV to MII during maturation. 39 However, Gomez's study suggests an increase in the expression level of this gene from the GV stage to MII. 40 STIM1 recognizes the reduction in Ca2+ concentration inside the ER lumen. Aggregates are formed, and they work along with ORAI1 which resides in the PM as they get closer to it. Ca2+ entrance is facilitated as a result, which helps restock the stores. 41 The 20 mT SMF may likely impact the PM in cells. MII oocytes show increased expression as calcium entry into the cytoplasm is necessary for the maturation process.
Vitrification reduces the IVF rate due to the abnormal calcium ion variation in the ER of the oocyte. The IVF rate depends on Mt changes and ATP levels. Increased permeability of the organelle's membrane to calcium is one of the mechanisms that lower the Mt membrane potential. Thus, raising the calcium level in mitochondria damages the membrane. 42 Optimal vitrification improves the survival rate of the oocytes, increases the rate of fertilization, and, consequently, increases the rate of implantation and live birth. 43 In this study, the fertilization rate in the Vit group decreased with regard to the nVit group. This trend was improved through the 20 mT SMF possibly the maintenance of the Mt functionality.
In addition, the mitochondria have an inner and an exterior membrane. The voltage-dependent anion channel (VDAC) is a key channel found in the outer membrane. These channels allow for the transit of anions and are sensitive to them in the open state. When closed, they are cation sensitive. 44 According to Calabrio's study, the change in orientation of the N-terminal helix in VDAC by SMF causes the channel to be in the open state. The Mt transmembrane potential is reduced. Based on the Nernst–Planck equation, this happens when the gradient concentration between the outer and inner regions of the Mt membrane decreases. 45 It appears that the VDAC is open given the impact of SMF on the opening of the voltage channels. By keeping calcium out of the mitochondria, freezing injury is prevented.
The mitotic spindle, a group of microtubules that are principally responsible for organizing chromosomes during cell division, is another cell organelle that plays a role in embryo formation and development. It is made up of biological substances, including proteins, lipids, and DNA, that exhibit diamagnetic properties. No changes in these materials appear to be caused by the mild SMF (20–60 mT), and this agrees with the findings of Zhao et al. 46
In conclusion, the application of an SMF with a 20 mT intensity increases the IVM, IVF, and membrane potential. Calcium oscillations, which are crucial for maturation and fertilization, are impacted by SMF.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any grant from funding agencies in the public, commercial, or not-for-profit sectors.