
Abstract
Aim:
Artificial propagation of ring-necked pheasant through semen preservation is of significance, as this species is facing enormous threats in its natural habitat. Semen preservation inevitably induces oxidative stress, and exogenous antioxidants need to be investigated for the preservation of ring-necked pheasant semen. Therefore, the current study was conducted to investigate the role of glutathione (GSH) in extender on the liquid storage of ring-necked pheasant semen.
Materials and Methods:
Semen was collected from 10 sexually mature males, evaluated for sperm motility, and pooled. Pooled semen was aliquoted for dilution with Beltsville poultry semen extender (1:5) at 37°C having GSH levels of 0.0 mM (Control), 0.2, 0.4, 0.6, and 0.8 mM. Extended semen was gradually cooled to 4°C and stored in a refrigerator (4°C) for 48 hours. Semen quality, that is, sperm motility, membrane integrity, viability, acrosomal integrity, and DNA integrity, was assessed at 0, 2, 6, 24, and 48 hours.
Results:
Sperm motility (%), plasma membrane integrity (%), viability (%), and acrosomal integrity (%) were recorded higher (p < 0.05), whereas DNA fragmentation (%) was recorded lower in extender supplemented with 0.4 mM GSH up to 48 hours of storage compared with 0.2, 0.6, and 0.8 mM GSH concentrations and control.
Conclusion:
It is concluded that 0.4 mM GSH in extender improves sperm quality parameters of ring-necked pheasant during liquid storage up to 48 hours at 4°C.
Introduction
The ring-necked pheasant also called common pheasant (Phasianus colchicus) belongs to the family “Phasianidae,” 1 native to Eurasia, 2 and is considered a well-known game bird. In addition to its regional significance, it is the ancient one in the world and is listed as least concern under IUCN red list of threatened species. 3 However, the ring-necked pheasant is one of the world's most hunted birds and is used for meat purpose in several countries.4,5
Further, remarkable transformations in farming practices, excessive use of agriculture related chemicals, increased grazing pressures, spraying and mowing of highways, and utility rights-of-ways during the last half of the 20th century have contributed a lot toward reducing this bird's population in the wild. The ring-necked pheasant's high dependence on habitats and niches in or around croplands/agricultural landscapes is driving it toward risk of declining. The major factors include conversion of open, indigenous grasslands and other unused habitats to managed grasslands, removal of luxuriant hedgerows, placement of fence rows, and clean farming acts that lead to a loss of its nests and protective covers.6,7
Conservation of species beforehand has key importance in the management of declining populations. 8 To achieve this, ex-situ conservation in conjunction with in-situ conservation programs may prove useful specifically under current scenario of habitat degradation. Ex-situ in-vitro management practices include germ cell storage for future use in assisted reproductive technologies, more commonly the cryopreserved semen. However, the liquid storage of semen has its own advantages; specifically, it induces lesser damage to sperm compared with freezing procedures. Liquid stored semen (5°C) serves to acclimatize in-vitro sperm storage conditions, primarily suppress the metabolism for days, and lower sperm energy utilization to enable survival for a longer time span.9,10
In addition, liquid storage can generate 10 times higher dose number of ejaculated semen compared with frozen storage, which improves the breeding capacity of animals and brings about the balance of genetic variability among propagated individuals. Hence, the requirement of a lesser number of semen ejaculates with higher dose rates reduces the burden of rearing many males. Further, the expense of processing liquid stored semen is much less compared with cryopreservation due to exclusion of some equipment and continuous supply of liquid nitrogen. 10
The sperm of Galliformes have unique physiology and comprise narrower sperm head width and smaller volume of cytoplasm, which makes them more vulnerable to the damage during preservation. 11 Specifically, avian sperm is more vulnerable to per-oxidative damage due to the high content of poly unsaturated fatty acids in its membrane, which adds on to the pathological state of in-vitro stored sperm.11–13
Naturally, the imbalance of reactive oxygen species (ROS) is prevented by a well-defined antioxidant system of avian semen 14 ; however, during in-vitro storage, this system is affected and brings several damaging effects, including per-oxidative damage to plasma membrane and chipping of DNA strand in sperm.15,16 It is found that the level of antioxidants is reduced due to dilution with the media.17,18 This depletion of natural antioxidants could be managed through augmentation of exogenous alternates.19,20
One of the key antioxidants of semen is glutathione (
Materials and Methods
Ethical statement
The study was approved by the Ethics Committee of Pir Mehr Ali Shah Arid Agriculture University Rawalpindi, Pakistan, for the use of animals.
Animal management
Ten adult male Ring-necked Pheasants (P. colchicus) were managed at Avian Research Station, PMAS Arid Agriculture University Rawalpindi. Birds were reared under a natural photoperiod, that is, 16 hours of light and 8 hours of dark and temperature conditions in individual sand floor pens of 106.68 × 121.92 cm with roof sleeve. Each bird was provided with 100 g of commercially available poultry cock breeder feed daily, and fresh water was provided ad libitum during the experimentation period.
Semen collection and preliminary evaluation
Semen was collected in semen collection vials by massage technique during morning hours of the day. 24 Micropipette (μL) was used to measure semen volume. Each ejaculate's sperm motility was determined initially as described by Zemjanis. 25 Percentage of motile spermatozoa was assessed by placing a drop of semen sample on pre-warmed glass slide (37°C) and observed under a phase-contrast microscope at 400 × (Olympus BX20, Japan). Sperm concentration was measured by using 1 μL of semen and 200 μL of formal citrate solution (1 mL of 37% formaldehyde in 99 mL of 2.9% [w/v] sodium citrate) under a phase-contrast microscope (400 × ; Olympus BX20) through Neubauer's hemocytometer (Marienfeld, Germany).
Extender composition and sample processing
Beltsville Poultry Semen Extender (BPSE) was prepared by adding fructose (0.3 g), potassium citrate (0.0384 g), sodium glutamate (0.5202 g), magnesium chloride (0.0204 g), di-potassium hydrogen phosphate (0.7620 g), potassium di-hydrogen phosphate (0.039 g), sodium acetate (0.258 g), and N-tris(hydroxymethyl) methyl-2-aminoethane sulfonic acid (0.3170 g) to 100 mL of double-distilled water (pH 7.3, osmotic pressure 330 mOsm/kg). BPSE extender was supplemented with five different concentrations of reduced glutathione, that is, 0.0, 0.2, 0.4, 0.6, and 0.8 mM. Qualifying ejaculates (>60% motility) were pooled. Five semen aliquots were made and diluted 1:5 with BPSE. Extended semen was cooled in 2 hours from 37°C to 4°C and kept for 48 hours in a refrigerator (4°C). Hence, samples were assessed for sperm motility, sperm membrane integrity, viability, acrosomal integrity, and sperm DNA fragmentation at 0, 2, 6, 24, and 48 hours of liquid storage.
Semen quality parameters
Sperm motility
Percent sperm motility was measured on a scale of 0% to 100% by placing a droplet of diluted sample on a pre-warmed slide (37°C) that was covered by a cover slip. The slide was examined under a phase-contrast microscope (400 × ). 25
Sperm membrane integrity
Plasma membrane integrity of Ring-necked Pheasant spermatozoa was evaluated using Hypo-osmotic swelling test. 26 Hypo-osmotic swelling (HOS) solution was made by diluting 1 g of sodium citrate with 100 mL of distilled water. A 25 μL diluted semen sample was mixed with 500 μL of HOS solution (100 mOsm/kg) and was incubated at 37°C for 30 minutes. A droplet of this solution was fixed in buffered 2% glutaraldehyde on a pre-warmed glass slide. Spermatozoa with swollen heads and swollen plus coiled tails were considered normal with intact plasma membranes under a phase-contrast microscope (1000 × with oil immersion). A total of 200 spermatozoa were counted at five different fields on a slide.
Sperm viability
Sperm viability of Ring-necked Pheasant spermatozoa was obtained using water soluble eosin (1 g) and water-soluble nigrosine (5 g), which was added to Lake's glutamate solution. 27 Lake's glutamate solution was made by adding potassium citrate (0.00128 g), sodium glutamate (0.01735 g), magnesium chloride (0.000676 g), and sodium acetate (0.0085 g) to 100 mL distilled water. Then, 12 drops of stain were homogenized with 1 droplet of sample; smear was formed and air dried. Smear was observed under a phase-contrast microscope (1000 × with oil immersion). Observed spermatozoa were classified into two categories: alive (physically normal and unstained) and dead (totally or partially stained). A total of 200 spermatozoa were observed at five separate fields on a slide. 18
Sperm acrosomal integrity
Sperm acrosomal integrity of Ring-necked Pheasant was evaluated via Giemsa stain.8,28 Stain was made by mixing Giemsa (3 g) and phosphate-buffered saline (PBS) at pH 7.0 (2 mL) into 35 mL water. Five microliters of semen sample was used to make smear on a glass slide and then air dried and fixed by neutral formal saline (5% formaldehyde) for 30 minutes. Fixed slides were immersed in Giemsa stain for 1.5 hours. Sperm having normal acrosome were evenly stained, whereas sperm having abnormal acrosomes were unevenly stained and sperm with ruptured acrosomes remained unstained. A sum of 200 spermatozoa was observed in five different fields under a phase-contrast microscope ( × 400; Olympus BX20) at 1000 × with oil immersion.
Sperm DNA fragmentation
Sperm DNA fragmentation was determined by aniline blue stain. Staining solution was prepared by mixing 5 g of aniline blue to 100 mL of PBS. Then stain was filtered, and its pH was adjusted to 3.5 with the addition of 2% glacial acetic acid. Ten microliters of semen was smeared on a glass slide and air dried. Slides were stained for 5 minutes with aniline blue stain and then rinsed with distilled water and air dried. A total of 200 spermatozoa was observed using a phase-contrast microscope (1000 × with oil immersion). The spermatozoa stained dark blue were regarded as having fragmented DNA. 29
Statistical analysis
Data are presented as mean ± SEM. The data on different concentrations of reduced glutathione in extender during liquid storage of Ring-necked pheasant semen were analyzed through two-factor factorial analysis of variance in MSTAT-C® software (version 1.42; Michigan State University, East Lansing, MI) in a completely randomized design. When F-ratio was found significant (p < 0.05), a comparison of means was done by fisher-protected least-significant difference test.
Results
Effect of GSH on sperm motility during liquid storage
The data on the effect of different concentrations of GSH in extender on sperm motility at different hours (0, 2, 6, 24, and 48) of liquid storage are presented in Figure 1. Addition of GSH in extender at the concentration of 0.4 mM was found to maintain significantly higher sperm motility (p < 0.05) at all the storage hours compared with other concentrations of GSH, that is, 0.2, 0.6, 0.8 mM, and control. At all hours, lower concentrations of GSH, that is, 0.2 and 0.4 mM were found to increase sperm motility, whereas higher concentrations of GSH, that is, 0.6 and 0.8 mM resulted in a gradual decrease of sperm motility though it remained higher compared with the control.

Effect of GSH concentrations (0.0, 0.2, 0.4, 0.6 and 0.8 mM) on sperm motility (%) of Ring-necked pheasant semen at different hours of liquid storage. Bars with different letters show significant differences (p < 0.05) throughout the storage hours. GSH, glutathione.
Effect of GSH on sperm membrane integrity during liquid storage
The data on the effect of addition of different concentrations of GSH in extender on plasma membrane integrity of spermatozoa at different hours (0, 2, 6, 24, and 48) of liquid storage are presented in Figure 2. At all storage hours, sperm plasma membrane integrity was improved with the supplementation of GSH at all levels compared with control. However, GSH supplementation at lower levels, that is, 0.2 and 0.4 mM were found to improve plasma membrane integrity compared with further higher concentrations of GSH, that is, 0.6 and 0.8 mM. Sperm plasma membrane integrity (%) was found to be significantly higher (p < 0.05) with the addition of GSH at the concentration of 0.4 mM in the extender in comparison to other concentrations (0.2, 0.6, 0.8 mM) and control at all hours of liquid storage.
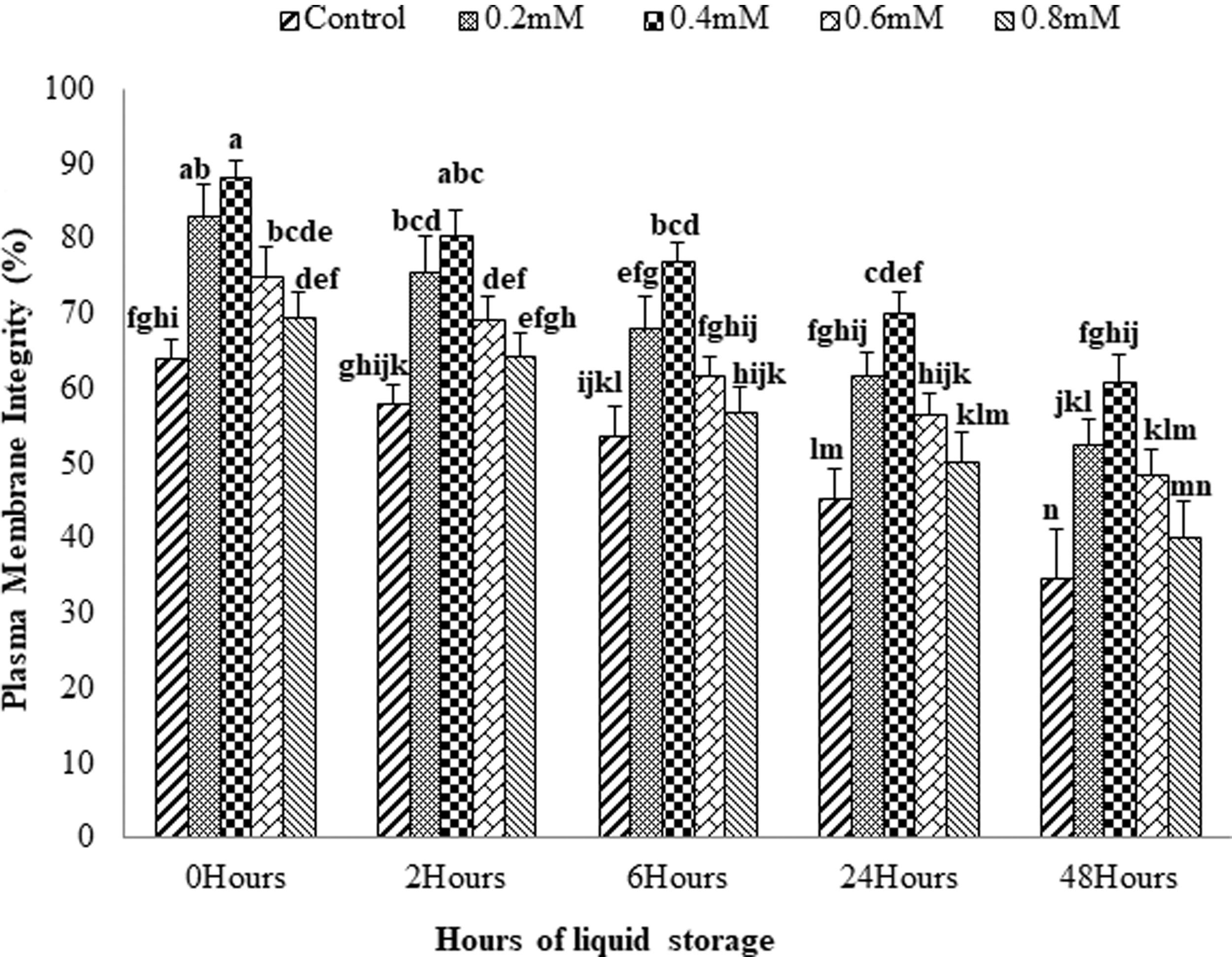
Effect of GSH concentrations (0.0, 0.2, 0.4, 0.6, and 0.8 mM) on plasma membrane integrity (%) of ring-necked pheasant spermatozoa at different hours of liquid storage. Bars with alphabetical letters show significant differences (p < 0.05) throughout the storage hours.
Effect of GSH on sperm viability during liquid storage
The data on the effect of various concentrations of GSH in extender on sperm viability at different hours (0, 2, 6, 24, and 48) of liquid storage are presented in Figure 3. Addition of GSH in extender at 0.4 mM was recorded to maintain higher (p < 0.05) sperm viability at all the storage hours compared with other concentrations of GSH (0.2, 0.6, 0.8 mM) and control. The trend for sperm viability remained the same as for both sperm motility and plasma membrane integrity. The sperm viability was improved with the addition of GSH at lower concentrations (0.2 and 0.4 mM) compared with higher concentrations (0.6 and 0.8 mM), though it remained higher compared with control at all concentrations throughout the storage period.

Effect of GSH concentrations' (0.0, 0.2, 0.4, 0.6, and 0.8 mM) supplementation on sperm viability of ring-necked pheasant spermatozoa at different hours of liquid storage. Bars with various letters represent significant differences (p < 0.05) throughout the storage hours.
Effect of GSH on sperm acrosomal integrity during liquid storage
The data on the effect of supplementing different concentrations of GSH in extender on acrosome integrity of spermatozoa at different hours (0, 2, 6, 24, and 48) of liquid storage are displayed in Figure 4. Addition of GSH in extender at the concentration of 0.4 mM was found to maintain acrosomal integrity significantly higher (p < 0.05) compared with other concentrations of GSH (0.2, 0.6, 0.8 mM) and control at all hours. Improvement in acrosomal integrity was evident at lower concentrations of GSH, that is, 0.2 and 0.4 mM; however, higher concentrations of GSH (0.6 and 0.8 mM) resulted in decreased acrosome integrity at all storage hours though it remained higher compared with control.

Effect of different concentrations of GSH (0.0, 0.2, 0.4, 0.6, and 0.8 mM) on sperm acrosomal integrity of ring-necked pheasants at several hours of liquid storage (4°C). Error bars with letters depict significant differences (p < 0.05) throughout the storage hours.
Effect of GSH on sperm DNA fragmentation during liquid storage
The data on the effect of supplementing various concentrations of GSH in extender on sperm DNA fragmentation at different hours (0, 2, 6, 24, and 48) of liquid storage are presented in Figure 5. Addition of GSH in extender at the concentration of 0.4 mM was found to reduce (p < 0.05) DNA fragmentation compared with other concentrations of GSH (0.2, 0.6, 0.8 mM) and control at all observed hours. It was viewed that with the lower GSH levels there was a decrease in DNA fragmentation, that is, 0.2 and 0.4 mM but further higher concentrations of GSH; that is, 0.6 and 0.8 mM were found to gradually increase the DNA fragmentation at all storage hours. However, DNA fragmentation remained lower than the control.
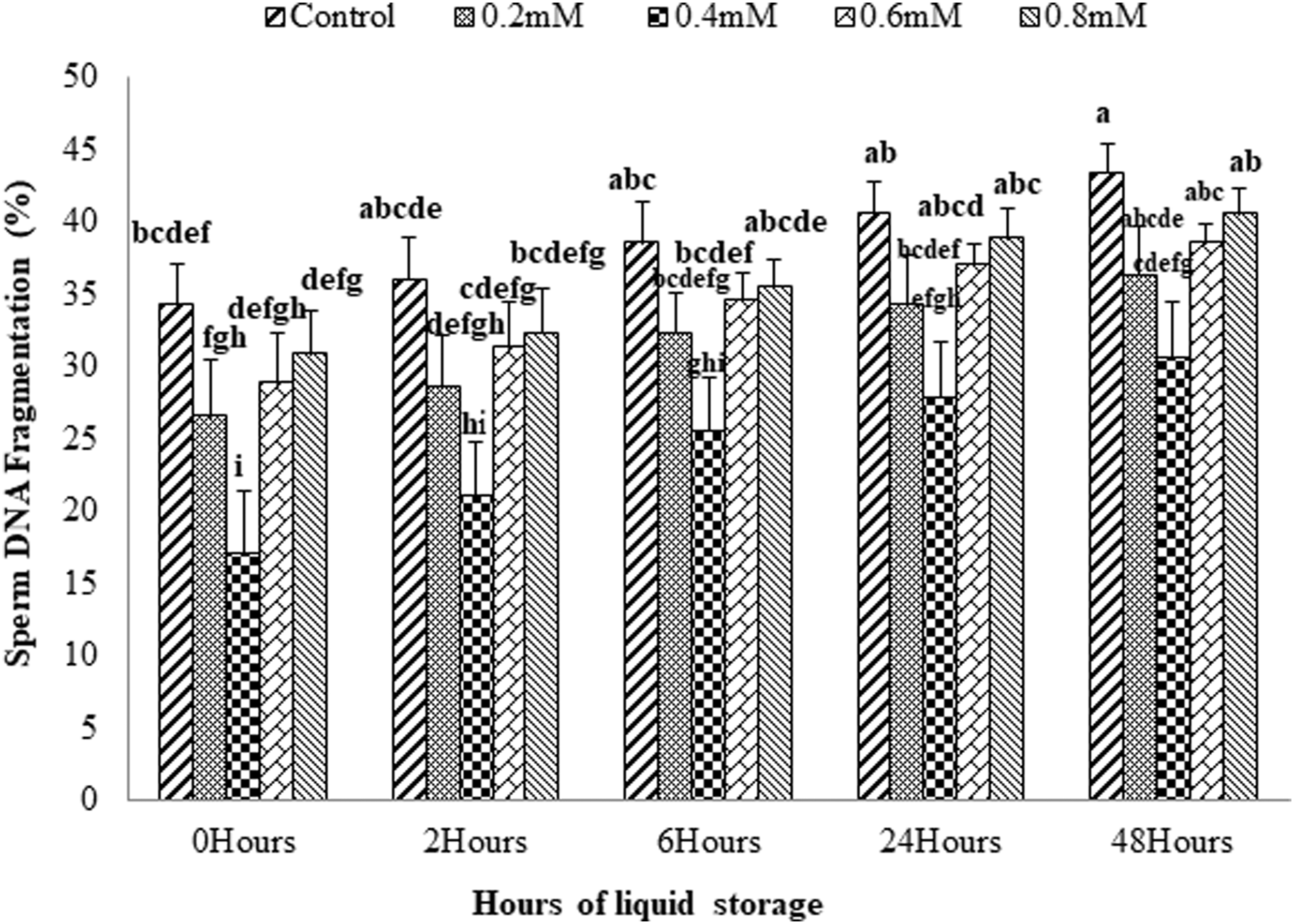
Effect of supplementation of GSH concentrations (0.0, 0.2, 0.4, 0.6, and 0.8 mM) on sperm DNA fragmentation of ring-necked pheasant. Error bars along with letters depict significant variations (p < 0.05) throughout the storage hours.
Discussion
Glutathione is an intracellular defense tool that is used for combating oxidative stress, especially in its reduced form. Many studies have reported the positive impact of GSH supplementation in liquid stored30–33 mammalian sperm, whereas fewer studies are available on liquid-stored avian semen. Further, among avian species, the impact of antioxidants is mainly studied in liquid-stored semen from different chicken breeds.34–36 Therefore, the full potential of avian semen to tolerate conditions of liquid storage is yet to be explored, as avian sperm has a long head and little cytoplasm, making it more sensitive to low temperatures compared with the sperm of other species. 37
In-vitro manipulation of avian semen for successful implementation of assisted reproductive techniques requires dilution with suitable media and appropriate storage temperatures, which could provide sperm with a minimum shelf-life of 2 to 4 days. 10 In the present study, liquid storage of ring-necked pheasant semen was performed for 48 hours at 4°C and time-dependent decline was observed in semen quality at 0, 2, 6, 24, and 48 hours with maximum semen quality recorded at initial storage hours. Extended storage hours and days for liquid preservation of semen severely affect sperm characteristics due to oxidative stress 38 and decline of antioxidants. 39
In such cases, supplementation of exogenous antioxidants is required to maintain the semen quality for maximum utility of liquid-stored semen.40,41 Reduced glutathione is one of the major antioxidants in semen and can be supplemented in media to replenish its level during in-vitro semen preservation.42–44 According to current results, semen quality is observed maximum at 0.4 mM GSH and found to decline above this level; however, it remained higher compared with the control. Structurally, reduced glutathione is a natural molecule in sperm cells, 45 composed of cysteine, glutamate, and glycine amino acids. It is utilized to maintain membrane proteins and nucleic acids due to its redox power.46–48
The acrosome and sperm membranes are considered significant predictors of fertility49–51 and these are the primary sperm structures affected due to altered physiology of in-vitro stored semen.50,52 Lipid peroxidation and protein denaturation in plasma membrane is associated with the loss of sperm motility,47,53 whereas antioxidants provide longevity and acrosomal intactness to spermatozoa by preventing ROS buildup until acrosomal reaction.53,54
The present study reported maintenance of motility, intact acrosome, and plasma membrane altogether with GSH fortification of extender for liquid storage of ring-necked pheasant semen at all stages. The avian sperm are more sensitive to cryodamage due to their elongated shape; hence, the anti-oxidative impact of GSH on avian sperm amplifies during initial storage time, 18 whereas for mammalian sperm GSH is reported to show significant effects after temperature decline/freezing. 55 Still there are minor reports of immediate GSH influence on mammalian semen in terms of livability and intact acrosome. 56
Further, GSH was reported to exert beneficial effects on diluted/cooled cockerel semen in terms of semen quality parameters regardless of the prolonged storage period (72 hours) compared with control. 54 It has previously been reported that without exogenous antioxidants imbalance of saturated to unsaturated fatty acids ratio and loss of cholesterol 57 results in the leakage of cell constituents.
The viable sperm with intact DNA succeed toward infundibulum and fuse with the ovum8,58,59; however, the ROS react with normal metabolic components in the absence of anti-oxidative checkpoints and cause telomere reduction, DNA epigenetic changes, and Y chromosomal micro-deletions 60 with serious repercussions on post-fertilization development. 61 It is well known that GSH protects against ROS and electrophiles by its sulfhydryl group, which is a strong nucleophile. 62 The current results showed that sperm viability and DNA integrity consistently remained higher with 0.2 and 0.4 mM GSH from 0 to 48 hours.
It indicates that GSH maintained sperm physiology and DNA structure in its original state soon after dilution and during liquid storage. Previous studies reported reduced DNA damage and higher fertility rates due to GSH addition in liquid storage of cockerel, 54 turkey, 37 rooster, 63 and buffalo semen.64,65 The DNA damage is correlated with ROS levels37,66 and the role of GSH as antioxidant for in-vitro stored semen has been reported.
Possibly, the natural GSH in control was exhausted in the current study, resulting in a faster decline of sperm DNA integrity compared with GSH-treated groups. However, excessive GSH may disrupt the normal metabolic needs of cell and produce ill-effects due to the absence of physiological levels of ROS. 18 This study reports the efficiency of glutathione for liquid storage of ring-necked pheasant semen and its inclusion at 0.4 mM of extender was identified best for storage up to 48 hours. These findings pave a way toward investigation beyond cooling and for evaluation of reduced glutathione in extender in terms of quality and fertility of cryopreserved ring-necked pheasant semen.
Conclusion
It is concluded that 0.4 mM of GSH in extender enhances motility, membrane integrity, viability, and acrosomal integrity whereas it lowers DNA fragmentation of ring-necked pheasant sperm during liquid storage at 4°C.
Footnotes
Authors' Contributions
S.Z.: Management of Study Animals (lead), Investigation (lead), Formal Analysis (lead), Methodology (lead), Writing-Original draft (lead). B.A.R.: Conceptualization (lead), Project administration (lead), Methodology (lead), Supervision (lead). S.A.: Resources (lead), Visualization (lead), Validation (lead), Writing-Review and editing (equal). M.S.A.: Data Curation (lead), Validation (supporting), Writing-Review and Editing (equal) K.W.: Management of Study Animals (Supporting), Formal Analysis (Supporting).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.