
Abstract
Somatic cell biobanking is a promising strategy for developing reproductive techniques. Although cryopreservation, a technique used for creating biobanks, has been performed on Galea spixii, structural and physiological damage to its cells highlight the need to optimize the cryoprotective solution being used. Therefore, the osmoprotective activity of 5 mM L-proline was evaluated as an alternative cryoprotectant for G. spixii fibroblast conservation. The concentration was defined based on previous studies conducted on mammalian cells. Cells derived from the skin of six individuals were cultured until the fifth passage were cryopreserved under the following treatments: (i) control (non-cryopreserved); (ii) a solution with 10% dimethyl sulfoxide (Me2SO), 10% fetal bovine serum (FBS), and 0.2 M sucrose; (iii) a solution with 10% Me2SO, 10% FBS, and 5 mM L-proline; and (iv) a solution with 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline. Tests were conducted to analyze cell morphology, viability, metabolism, proliferation, and apoptosis; reactive oxygen species (ROS) levels; and mitochondrial membrane activity (ΔΨm). A reduction in the number of viable cells (72.3% ± 1.2%) was observed in the sucrose-containing group compared to the control (86.7% ± 2.0%) and L-proline (88.4% ± 1.8% and 87.8% ± 2.1%) groups. After apoptotic analysis, a reduction in the number of viable cells was observed in the group with sucrose alone (74.6% ± 4.1%) compared to the control group (88.2% ± 1.1%). The ROS levels (1.03 ± 0.5 and 1.07 ± 0.5, respectively) and ΔΨm values (0.99 ± 0.42 and 1.22 ± 0.73, respectively) observed in the groups with L-proline were similar to that observed in the control group (1.00 ± 0.5 and 1.00 ± 0.4, respectively). Moreover, no difference was observed between groups for cell morphology, metabolism, or proliferation. Thus, L-proline is a cryoprotectant agent that can be used during G. spixii fibroblast cryopreservation, alone or with sucrose. In addition, we developed an adequate biobank for G. spixii, whereby stored cells could be used for reproductive techniques.
Introduction
Cellular biobanks are a collection of isolated in vitro-cultured somatic cells. They enable the formation of an organized and searchable collection of biological samples, 1 and provide relevant information for the management, optimization, and development of protocols for wild species. With respect to laboratory and wild rodents, biobanks offer applications for the treatment of genetic diseases and development of regenerative therapies. 2 Furthermore, somatic cell banks offer an opportunity for species conservation, 3 and genetic reintroduction of populations threatened with extinction through assisted reproduction techniques, 4 as observed in cloning by somatic cell nuclear transfer 5 and the induction of cells to pluripotency. 6 In addition, with the formation of cellular banks for wild rodents, it is possible to implement other techniques irrespective of limiting factors, such as reproductive dysfunction, unexpected deaths, geographical isolation, and the age and sex of the animal. 7
Spix’s yellow-toothed cavy (Galea spixii) is a wild rodent that was initially used to develop a biobank derived from fibroblastic lines. 8 G. spixii is an herbivorous rodent native to the Caatinga biome, but is located in other biomes, such as in Brazil, Paraguay, Bolivia, and east of the Andes Mountains. 9 This species is 22.5–23.5 cm long, 10 with uniform-colored fur, a gray back, white belly, and no tail. 11 G. spixii contributes to the maintenance of local fauna and flora. 12 Despite environmental disturbances associated with hunting this rodent, 13 its population is considered stable 14 because of its adaptive, physiological, reproductive, and genetic characteristics. 15 Therefore, it stands out as an ideal study model for developing protocols involving biobank formation at scientific breeding sites, and among other wild rodents.
For G. spixii, a biobank of somatic tissues has already been developed and used to observe the structural and morphological characteristics of the skin, which is an organ from which somatic cells can be isolated in a minimally invasive, inexpensive, and rapid manner. 16 The establishment and cryopreservation of fibroblastic lines is another technique implemented for wild rodents. 8 After cell cryopreservation, changes are observed in the cell viability and proliferation rate, owing to an imbalance in their cellular redox system and an increased number of apoptotic cells, probably caused by osmotic stress and ice crystal formation during cryopreservation. This may have caused lipid peroxidation, which is an important event in cell membrane damage during cryopreservation. 17 Therefore, it is necessary to optimize this process.
The first parameter evaluated was the selection of the cryoprotective solution. The first protocol established for G. spixii was based on protocols used for other phylogenetically close species,3,18 in which the cryoprotectant solution comprised 10% dimethyl sulfoxide (Me2SO), 10% fetal bovine serum (FBS), and 0.2 M sucrose. 8 However, after in vitro culturing of the cryopreserved fibroblastic lines, 8 their viability, metabolism, and proliferative rate were affected, probably due to the osmotic stress generated by intracellular and extracellular ice crystal formation. 17 Herein, only the cryopreservation mechanisms of the cryoprotectants used were studied, such as Me2SO, which promotes the osmotic dehydration of cells by penetrating into the cytosol and preventing intracellular ice crystal formation. 19 Moreover, sucrose, a disaccharide, promotes the stabilization of the lipid membrane because of direct interactions with the polar components of the cell bilayer, preventing the aggregation of intracellular molecules and osmotic stress. 17
However, it is also necessary to consider species-specific characteristics, as cryopreservation protocols are dependent on these characteristics. This is true for G. spixii, in which physiological characteristics, such as size, weight, and metabolic rate, differ from other laboratory and wild rodents. 15 Therefore, considering the damage observed after G. spixii fibroblast cryopreservation, and the need to optimize the cellular cryopreservation protocol for G. spixii, selecting new cryoprotective agents facilitates the promotion of greater membrane stability and controls osmotic stress. Thus, L-proline may be used as an alternative cryopreserving agent, because in red blood cells, 19 lung carcinoma cells, 20 endothelial cells, 21 and mouse oocytes, 22 it has been found to have multifunctionality after cryopreservation.
L-proline, a natural amino acid, is structurally formed by a non-polar cyclic pyrrolidine side chain with an amino group.19 In endothelial cells, L-proline has been implicated as a cell survival factor at a concentration of 5 mM,21 as it protects cells against osmotic stress and maintains cell cycle progression through the binding and transmission of signal transporters that participate in cell control and death.20,23,24 Furthermore, studies have shown that L-proline balances cell turgor pressure, reduces water stress, and controls the acidity of the cell culture medium, as it stabilizes proteins in the cytosol, preventing cryopreservation stress-induced denaturation, aggregation, or dissociation.23,25
Considering the positive characteristics of 5 mM L-proline, we hypothesized that such an agent could improve the viability and physiological quality of G. spixii fibroblasts. However, the isolated effect of L-proline and its synergistic effect with sucrose need to be investigated. Therefore, this study aimed to evaluate the synergistic interaction of L-proline with sucrose and isolated effect of L-proline on the viability and physiology of G. spixii fibroblasts.
Materials and Methods
Reagents and solutions were purchased from Sigma-Aldrich (St. Louis, MO, USA) and Gibco BRL (Carlsbad, CA, USA). The study protocol was approved by the Animal Use Ethics Committee of the Federal Rural University of Semi-Arid (CEUA/UFERSA, No. 23091.010566/2017-20) and Chico Mendes Institute for Biodiversity Conservation (ICMBio, No. 60428-1).
Animals, collection, and processing skin biopsies
Biopsies from the peripheral ear region were recovered from six adult male G. spixii individuals (1-year old) that were kept in scientific breeding facilities located at the Wild Animal Multiplication Center (CEMAS/UFERSA, Mossoró, RN, Brazil 5° 10′S, 37°10′ W, No. 1478912). To simultaneously collect all skin biopsies, samples sterilized with 70% alcohol with sizes ranging from 1.0 to 2.0 cm2 were obtained using surgical pliers. The samples were then transported to the laboratory in Dulbecco’s modified minimal essential medium (DMEM) with 10% FBS and 2% antibiotic–antimycotic (ATB–ATM) solution at 4°C for up to 1 hour. In the laboratory, skin biopsies were processed by first removing the hair, and then cutting them into sections of 9.0 mm³ (3.0 × 3.0 × 1.0 mm) for in vitro culture.
Primary culture of skin biopsies and cell subculture
Subsequently, a primary culture was set up with incubation at 38.5°C and 6.5% CO2.8 When the cells reached confluency ranging 70%–80%, they were subcultured and cell lines were established. 8 Trypsinization was then performed using 0.25% trypsin with 0.05% EDTA. This was considered the first cell passage. After the fifth passage, the cells were cryopreserved as described in the Experimental design section.
Experimental design
To evaluate the effect of L-proline during cryopreservation, fibroblasts were stored at a low temperature (−196°C) using the slow freezing technique.26,27 The fibroblasts were evaluated under the following experimental conditions: (i) control (non-cryopreserved); (ii) cryopreserved with a solution comprising 10% Me2SO, 10% FBS, and 0.2 M sucrose; (iii) cryopreserved with a solution comprising 10% Me2SO, 10% FBS, and 5 mM L-proline; or (iv) cryopreserved with a solution comprising 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline. The L-proline concentration used in this study was based on the cryopreservation of human endothelial cell lines. 22 In this study, 5 mM L-proline did not cause negative effects after the cells were recultured and stored at low temperatures (−196°C); however, higher concentrations generated high levels of osmotic stress.
We evaluated standard cryopreservation medium with the presence and absence of L-proline and synergism of L-proline.3,18 This allowed us to visualize its effects on reducing or controlling osmotic stress during the cryopreservation of fibroblastic lines derived from G. spixii. Finally, non-cryopreserved and cryopreserved cells were evaluated for their morphology and physiological characteristics, such as viability, metabolism, proliferation, apoptosis, levels of reactive oxygen species (ROS), and mitochondrial membrane potential (ΔΨm).
Slow freezing and thawing of fibroblasts
A concentration of 1.0 × 105 cells/mL was recovered from in vitro cultures, and the cells were counted using a Neubauer chamber. 8 The cells were then added to cryotubes containing DMEM, 10% Me2SO, and 10% FBS, and were maintained for 15 minutes at 4°C. After incubation, 0.2 M sucrose and 5 mM L-proline were added and the cells were further incubated for 15 minutes at 4°C.3,22 The Mr. Frosty system® (Thermo Scientific Nalgene, Rochester, NY, USA), a programmable freezer, was then used to freeze the cells held in a container for 12 hours at −80°C (1°C/min). The samples were stored in cryotubes in liquid nitrogen (−196°C) for up to 2 weeks.
The cryovials were thawed by initially incubating at 25°C for 1 minute, followed by 4 minutes in a temperature-controlled double boiler (35°C–37°C). DMEM containing 0.2 M sucrose or 5 mM L-proline was added to the thawed cells, according to the experimental groups,3,22 and they were maintained at 4°C for 15 minutes. After incubation, the cell suspension was centrifuged for 10 minutes at 600×g. The supernatant was removed, cells were resuspended in DMEM, 10% FBS, and 2% ATB–ATM solution, incubated for 15 minutes at 25°C, followed by centrifugation (10 minutes; 600×g). After centrifugation, the cells were recovered and subjected to in vitro culture using a previously described medium (DMEM, 10% FBS, and 2% ATB–ATM solution) and evaluated according to the experimental design.
Morphological evaluation and cell viability assay using trypan blue
Fibroblast morphology, before and after cryopreservation, nuclear shape, and cytoplasmic extension of the cells were evaluated using an inverted microscope (Nikon TS100, Tokyo, Japan). 28
A cell suspension (20 µL) of fibroblasts was recovered before and after cryopreservation (immediately after thawing and in vitro culturing) to determine the percentage of viable cells. For this, the cells were stained using trypan blue (20 µL; 0.4%), where non-viable cells (dead; stained blue) were those with ruptured membranes and viable cells (alive; colorless) were those with intact cell membranes. 28 A Neubauer chamber was used for cell counting and quantification was performed under an optical light microscope (CX23, Olympus Tokyo, Japan; magnification 10×) using the following formula: number of viable cells/number of total quantified cells.
Metabolic and proliferative activity of cells
The metabolic activity of cells was evaluated using the 3-(4,5-dimethyl-2-thiazolyl)−2,5-diphenyl-2H-tetrazoluim bromide (MTT) assay. 27 A concentration of 5.0 × 104 cells/mL was recovered and seeded in 12-well dishes. The cells were incubated for 5 days in an incubator (38.5°C with 6.5% CO2). After incubation, the MTT solution (5 mg/mL) was added, and after 3 hours under modified temperature and atmosphere (38.5°C and 6.5% CO2), the cells were analyzed at 595 nm using a spectrophotometer (UV-mini-1240; Shimadzu®, Kyoto, Japan).
To evaluate the rate of proliferation, cells were seeded in 24-well dishes at an initial concentration of 1.0 × 104 cells/mL for 7 days (168 hours). At intervals of 24 hours, after trypsinization, the average cell count was obtained, and used to generate a growth curve and calculate the population doubling time (PDT),
26
which was estimated using the following mathematical calculation:
The PDT is expressed in hours (h), where T is the incubation time, ln is Napier’s logarithm, Xe is the number of cells at the end of the incubation period, and Xb is the number of cells at the start of the incubation period.
Assessment of apoptotic levels
The apoptotic levels of fibroblasts were evaluated using a cell suspension (35 µL) in which the cells were stained with ethidium bromide (10 µg/mL) and acridine orange (2 µg/mL) diluted in 8 µL of phosphate-buffered saline. 29 Analysis was performed using a fluorescence microscope (Olympus BX51TF) at an absorbance of 480 nm. Cells were considered to be (i) viable if they had a uniform green nucleus; (ii) at the initial stage of apoptosis if they had non-uniform green nucleus; (iii) at a late stage of apoptosis if they had a uniform orange nucleus; and (iv) undergoing necrosis if they had a uniform red nucleus. 26 Images were captured at a magnification of 20×, and 300 cells were quantified using ImageJ software (National Institutes of Health, Bethesda, MA, USA).
Quantification of ROS and ΔΨm levels
Oxidative stress mediated by ROS molecules and ΔΨm was quantified using the fluorescent probe 2’,7’-dichlorodihydrofluorescein diacetate (H2DCFDA; 10 µM; Invitrogen, Carlsbad, CA, USA) and MitoTracker Red® (CMXRos; 500 nM; Invitrogen). Approximately 50 µL of cell suspension was labeled with fluorescent probes. After 30 minutes of incubation (38.5°C and 6.5% CO2), 100 cells per group were processed and the probes were analyzed using a fluorescence microscope (Olympus BX51TF). 8
Statistical analysis
Data obtained from six adult male G. spixii individuals were expressed as the mean ± standard error (one animal/one replicate) and analyzed using GraphPad Prism software (Version 10.2.3, Graph-Pad Prism Software Incorporation, La Jolla, CA, USA). Normality was assessed using the Shapiro–Wilk test, and homoscedasticity was assessed using Levene’s test. Data that were not normally distributed were transformed using arcsine. Data were compared using analysis of variance, followed by Tukey’s test or an unpaired Student’s t-test. Statistical significance was set at p < 0.05.
Results
No phenotypic change was observed in cellular morphology between the experimental groups (Fig. 1A–D). The cells maintained their fusiform shape, were elongated, and had a centralized nucleus. Moreover, we observed that, after cryopreservation and in vitro culture, fibroblasts exposed to the cryoprotectant solution containing 10% Me2SO, 10% FBS, and 5 mM L-proline or 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline were more adherent to the surface than those exposed to only 10% Me2SO, 10% FBS, and 0.2 M sucrose. This may explain the lower viability observed after the trypan blue assay (Fig. 2A), which demonstrated that, immediately after cryopreservation, the cell viability rate was lower for cells in the solution with 10% Me2SO, 10% FBS, and 0.2 M sucrose (72.3% ± 1.2%) than for non-cryopreserved cells (86.7% ± 2.0%). Moreover, cell viability was similar between cells cryopreserved in 10% Me2SO, 10% FBS, and 5 mM L-proline (88.4% ± 1.8%) and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline (87.8% ± 2.1%) and non-cryopreserved cells (Fig. 2A). However, no difference was observed among the experimental groups after in vitro culture (Fig. 2A).
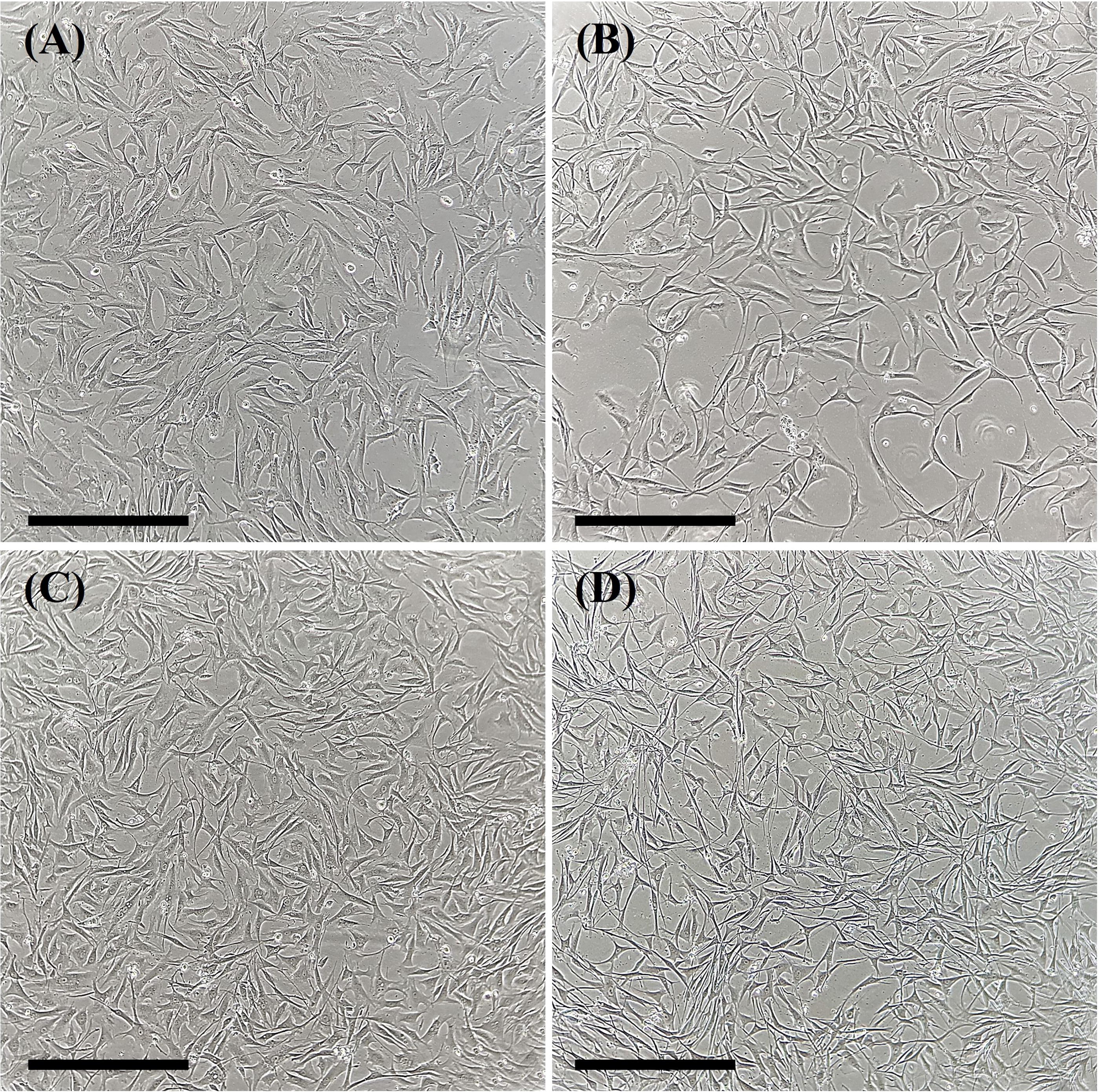
Representative images of groups of fibroblasts isolated from Galea spixii ear skin before and after slow freezing.

Viability analysis (immediately after cryopreservation and after in vitro culture) and physiological characteristics of G. spixii skin fibroblasts before and after slow freezing.
After cryopreservation, it was initially evident that cell metabolism was >90% in all experimental groups (Fig. 2B; p > 0.05), demonstrating the cell physiological functionality per group. Similar results were obtained when comparing the experimental groups after PDT analysis, in which the proliferation rate varied between 17 and 20 hours (Fig. 2C; p > 0.05). Although the metabolism and proliferation rate between experimental groups did not differ, after 24 hours of cell growth curve analysis (Fig. 2D), only the groups containing 10% Me2SO, 10% FBS, and 5 mM L-proline (22.3 ± 1.3 cells/mL) and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline (25.3 ± 1.6 cells/mL) had a higher concentration of cells than the non-cryopreserved group (13.7 ± 1.4 cells/mL). Furthermore, after 168 hours of evaluation, only the group containing 10% Me2SO, 10% FBS, and 5 mM L-proline (41.1 ± 3.5 cells/mL) and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline (50.0 ± 2.7 cells/mL) were similar to the non-cryopreserved (63.6 ± 11.8 cells/mL) group, while the group containing 10% Me2SO, 10% FBS, and 0.2 M sucrose (36.0 ± 3.0 cells/mL) had a lower cell concentration (Fig. 2D).
After apoptotic analysis of the cryopreserved cells (Fig. 3A–D), only the groups with 10% Me2SO, 10% FBS, and 5 mM L-proline (78.4% ± 3.4) and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline (78.8% ± 3.1) were similar to the non-cryopreserved group (88.2% ± 1.1, Fig. 3E). Furthermore, the 10% Me2SO, 10% FBS, and 0.2 M sucrose group (74.6% ± 4.1) showed a lower number of viable cells than the non-cryopreserved group (88.2% ± 1.1, Fig. 3E). An identical result was observed after evaluating the levels of ROS and ΔΨm (Fig. 4), where only the group containing 10% Me2SO, 10% FBS, and 0.2 M sucrose showed an increase in ROS levels (2.31 ± 0.9) and ΔΨm (1.27 ± 0.5) compared to the non-cryopreserved group (1.00 ± 0.5; 1.00 ± 0.4), whereas the groups containing 10% Me2SO, 10% FBS, and 5 mM L-proline (1.03 ± 0.5 and 0.99 ± 0.42) and Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline (1.07 ± 0.5 and 1.22 ± 0.73) had similar results.

Assessment of the percentage of apoptotic levels after in vitro culture of G. spixii skin fibroblast lines.
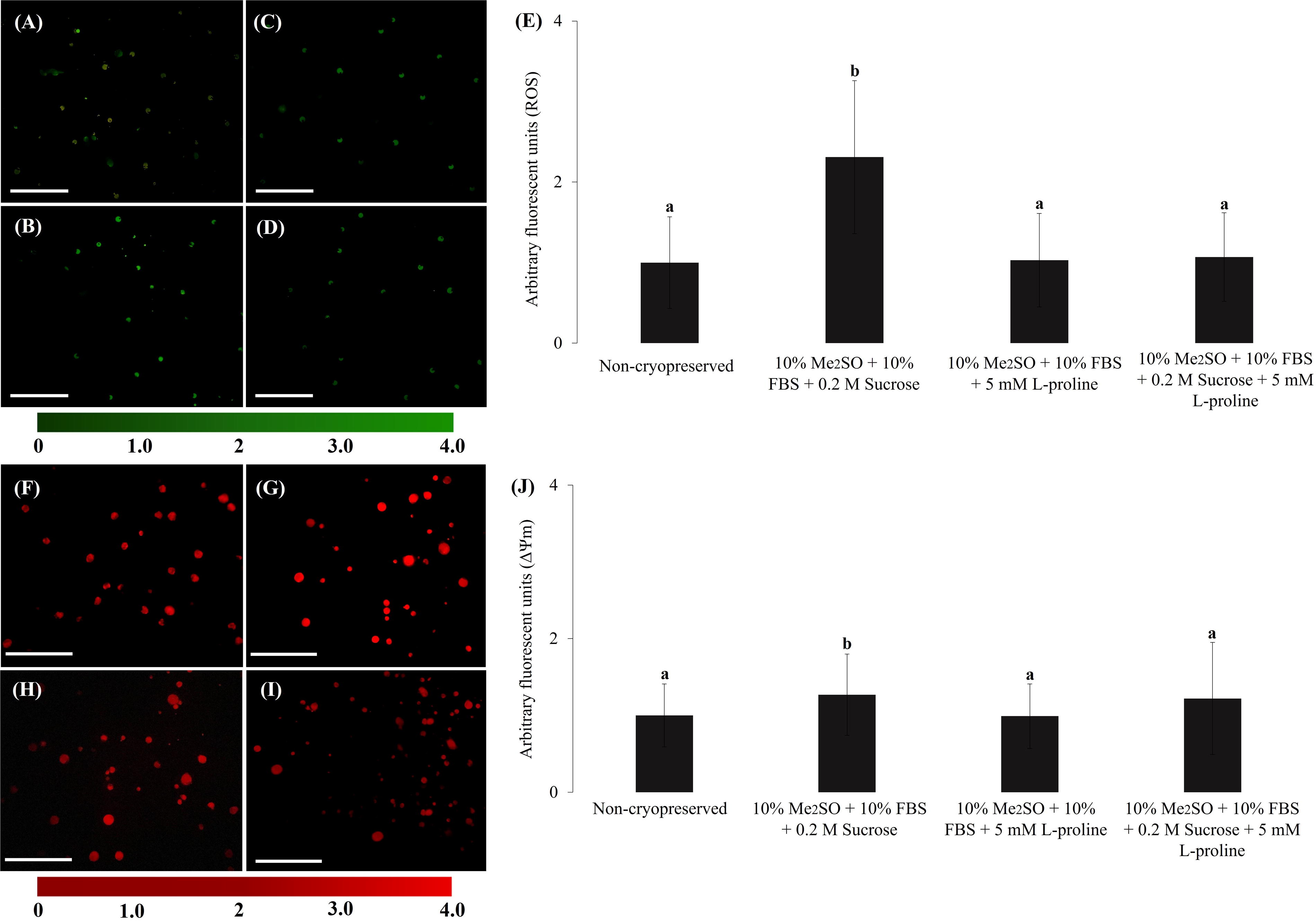
Assessment of intracellular levels of reactive oxygen species (ROS) and mitochondrial membrane potential (ΔΨm) of G. spixii fibroblasts before and after slow freezing.
Discussion
The formation of cell biobanks for wild rodents through cryopreservation significantly contributes to studies aimed at conservation, 3 as it maintains genetic resources. The samples can be used for basic and applied studies, to gain information regarding the physiological, reproductive, biological, and molecular nature of G. spixii. 30 For G. spixii, cryopreservation of somatic cells occurred for the first time using fibroblastic lines isolated from the skin by slow freezing. 8 Although this is a promising technique, cryopreservation by slow freezing results in morphological, structural, and physiological damage to the fibroblasts. In addition, cryopreservation causes an imbalance in the cellular redox system, altering ROS levels and ΔΨm. In other laboratory and wild rodents, such as Cavia porcellus, 18 Mesocricetus auratus, 26 and Dasyprocta leporina, 27 characteristics similar to those of G. spixii have been verified, leading to a reduction in the quality, viability, and stability of cells stored in cellular biobanks.
Due to the damage generated in the fibroblastic lines derived from G. spixii, we searched for alternatives to improve the quality of this cell type after cryopreservation. As the first parameter evaluated, the selection of the cryoprotectant solution components was an essential factor to be improved during slow freezing 31 as this guarantees greater cell viability and stability during in vitro culture and cryopreservation. We used a protocol usually applied to the cryopreservation of wild mammalian cells, 3 involving the use of 10% Me2SO, which is as an intracellular cryoprotectant that promotes multiple effects and actions on specific and non-specific targets. 21 Furthermore, Me2SO acts through the osmotic dehydration of cells, reducing ice crystal formation during nucleation and thawing events, which may lead to cell death. 32
In this same cryopreservation protocol, 0.2 M sucrose was used to stabilize the lipid membrane. Sucrose interacts directly with the cellular components present in the lipid bilayer, avoiding intracellular ice aggregation and providing cytoprotection. 21 Although cryoprotectants are widely used in the cryopreservation of wild rodents cells, 33 many studies have demonstrated that these agents are not always efficient. For instance, when Me2SO reaches different cellular compartments, it can cause negative effects that are required for the physiological activity of cells, causing changes in the enzymatic reactions of the mitochondria, modifications in transport mechanisms, and ionic exchange. 33 Therefore, an increase in the release of electrons increases ROS synthesis, leading to oxidative stress, as observed in some experimental groups in our study.
To optimize slow freezing during G. spixii fibroblast cryopreservation, with the aim of minimizing the deleterious effects caused by these cryoprotective solutions, L-proline was used either alone or in combination with sucrose as an alternative to synthetic congeners and their limitations, and its effects in terms of toxicity and cytoprotective function on cryopreserved G. spixii fibroblastic lines were studied. L-proline is a natural amino acid synthesized by microorganisms in response to cold exposure. It aids in eliminating oxidizing molecules 34 and involves certain biological functions, such as protein synthesis, 35 metabolism, 36 cellular structure maintenance, 25 and intracellular redox state regulation. 37
Using this approach, we initially observed no phenotypic difference between the fibroblasts of the experimental groups after morphological evaluation; this result was consistent with those of other studies involving somatic cells from laboratory and wild rodents.3,18 However, after analyzing the viability of these cells using the membrane exclusion test, we observed that, immediately after cryopreservation, cells in the group cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose had lower viability than those in the non-cryopreserved group and group cryopreserved with 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline. A similar result was observed in the study carried out by Jesus et al., 38 where superior viability was observed for HaCaT cells cryopreserved with L-proline compared to cells cryopreserved with only Me2SO. According to Bojic et al., 31 this may have occurred because of the close interactions between the cells and cryopreservation medium, causing the cells to be markedly more susceptible to damage caused by changes during the freezing and thawing phases.
In M. auratus, 26 a reduction in cell viability was observed immediately after cryopreservation due to the association between cell osmotic limits and the cryoprotective solutions used in the study (10% Me2SO and 90% FBS). In our study, L-proline guaranteed a greater interaction between the molecules of cell cytosol and extracellular environment to which this natural amino acid was subjected. Pi et al. corroborated this response, 39 by combining and observing the synergistic effect of L-proline during T-cell cryopreservation.
The results previously obtained are important because in our study, we also observed that, after cryopreservation and in vitro culturing of G. spixii fibroblastic lines, the groups cryopreserved in 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline had superior cell growth curves (cells/mL) than non-cryopreserved cells in the first 24 hours of evaluation. However, after 168 hours, the group cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose showed a lower number of cells (cells/mL) than the non-cryopreserved group. This may have occurred because after reculturing, the structural integrity of G. spixii fibroblasts was not positively supported when they were cryopreserved in a solution containing 10% Me2SO, 10% FBS, and 0.2 M sucrose only; however, the cells that were cryopreserved with 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline were less affected by slow freezing.
Therefore, cryopreserved cells containing L-proline recover more rapidly from injuries caused by ice crystal formation during freezing and thawing. In contrast, G. spixii fibroblasts cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose lost their biochemical activity, although they maintained intact cell membranes. These results corroborated those of a study on endothelial cells. 22 When compared with the control group (Me2SO) and cells treated with the other osmolytes evaluated, cells cryopreserved in a medium containing L-proline were more adherent to the in vitro culture dishes.
Although 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline promoted an increase in cell viability and proliferation rate, after analyzing the cell growth curve in comparison with cells cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose, we did not observe differences in PDT or metabolism. These results suggested that the cryoprotective agents tested, at the appropriate concentrations, are not toxic to fibroblastic lines isolated from the skin of G. spixii. Furthermore, cryoprotective solutions act positively in reducing and controlling ice crystal formation, and in processes related to recrystallization after cell thawing, especially in the presence of L-proline, to allow greater cell viability and proliferation after cryopreservation. 21
In addition, the apoptotic index of G. spixii fibroblasts was analyzed to predict cell death, which has been associated with osmotic stress events during the slow freezing of cells. 40 This assay also allowed cell quality and stability to be observed during in vitro culturing. 28 The number of viable cells was lower in the group cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose than in the non-cryopreserved group. However, the number of viable cryopreserved fibroblasts in the groups cryopreserved with 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline was identical. This may be because the group containing only sucrose may have initiated biochemical pathways related to programmed cell death, such as caspase-3 and caspase-7 pathways, which are highly active during cell death.41,42
Moreover, unlike L-proline, sucrose is a disaccharide with a mechanism of action similar to that of trehalose, allowing lipid membrane stabilization only because of direct interactions between the polar components of the cell bilayer. 17 L-proline exerts different biological functions, such as inducing the control of the p53 gene and enzyme synthesis, for example, proline oxidase (POX/PRODH), which is located in the inner mitochondrial membrane and catalyzes the transfer of electrons as well as regulates intracellular redox reactions, 37 thereby regulating the in vitro culture of cells.
This justification was verified in our study, as cells cryopreserved with 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline maintained ROS and ΔΨm levels similar to their levels in non-cryopreserved cells, whereas their levels were higher in cells cryopreserved with 10% Me2SO, 10% FBS, and 0.2 M sucrose. This may have occurred because free and polypeptide-bound proline can react with hydrogen peroxide (H2O2) and hydroxyl groups to form stable free radicals. 25 For instance, a study of the antioxidant action of 5 mM L-proline 43 found that, after 30 minutes, the H2O2 levels were reduced by >90% when compared to those of the control.
In addition, L-proline was added to a culture of human keratinocytes, and it was found to protect cells against induced apoptosis. 44 Therefore, this molecule reduces oxidative stress, ROS levels, and ΔΨm levels in the experimental groups containing 10% Me2SO, 10% FBS, and 5 mM L-proline and 10% Me2SO, 10% FBS, 0.2 M sucrose, and 5 mM L-proline, ensuring greater osmotic control as well as balance of deplasmolysis pressure and cell turgidity.20,44
Conclusions
This is the first study to optimize a technique for the cryopreservation of fibroblasts derived from G. spixii using L-proline. In our study, G. spixii cells stored at low temperatures, after thawing, L-proline alone or with sucrose remained viable and functional. Moreover, the cells cryopreserved with L-proline showed better maintenance of viability and proliferative rate, as well as control of ROS and ΔΨm levels. Therefore, L-proline, either alone or with sucrose, is an alternative for the cryopreservation of G. spixii-derived fibroblasts. Thus, it is possible to develop an adequate cellular biobank for G. spixii, in which the stored cells could be used to obtain biological, genetic, and reproductive knowledge, by somatic cell nuclear transfer cloning and inducing pluripotency.
Footnotes
Acknowledgments
The authors thank the Center for Wild Animals Multiplication (UFERSA) for providing biological material.
Authors’ Contributions
L.V.C.A.: Conceptualization, methodology, writing, original draft, and investigation. L.L.V.R.: Methodology and investigation. S.L.O.: Methodology and investigation. Y.L.F.S.: Methodology and investigation. L.R.M.O.: Methodology and investigation. Y.B.F.M.: Methodology and investigation. A.F.P.: Conceptualization, methodology, writing, original draft, supervision, project administration, and funding acquisition.
Author Disclosure Statement
The authors report no potential conflicts of interest.
Funding Information
This study was partially funded by the Coordination of Improvement of Higher Education Personnel, Brazil (CAPES, Financial Code