
Abstract
Reactive oxygen species (ROS) during cryopreservation causes mechanical, biochemical, and structural damage to the sperm, which leads to reduced sperm motility and fertility. N-acetyl cysteine is a cysteine-derived amino acid antioxidant that functions as a scavenger of ROS and regulates mitochondrial activity. Mitochondrial uncoupling protein 2 (UCP2) plays a leading role in this process and is one of the major regulators of human spermatozoa motility and metabolism. The purpose of the study was to examine the changes in UCP2 in frozen-thawed human sperm when exposed to N-acetyl cysteine, an effective antioxidant commonly used in human semen freezing. Semen samples were collected from 20 normozoospermia men and were divided into four experimental groups: fresh, frozen control, frozen N-Acetylcysteine (NAC, 100 μM), and frozen negative control with Genipin (25 μM). Subsequently, post-thaw sperm quality parameters, as well as UCP2 relative quantity, ROS, mitochondrial membrane potential (MMP), and malondialdehyde, were assessed. Semen treated with NAC exhibited significantly higher total and progressive motility, as well as viability, when compared to the control and genipin groups (p < 0.05). Moreover, UCP2 relative quantity was significantly lower in all frozen groups compared to the fresh group (p < 0.0001). The UCP2 relative quantity was not significantly different between NAC and control groups (p ≥ 0.05). Also, there were no significant differences in MMP, ROS, and malondialdehyde levels among the frozen groups (p ≥ 0.05). It can be concluded that UCP2 undergoes a modification during cryopreservation, and it could be an explanation of the reduction in post-thaw motility of sperm. Additionally, NAC supplementation in freezing media enhances post-thaw sperm motility and viability.
Introduction
Sperm cryopreservation is an essential method for preserving male fertility. However, this approach encounters the rise of oxidative stress in freezing-thawing sperm alongside the unavoidable rise of mitochondrial destruction as cryo-injury in the freezing process. Mitochondria are the primary source of reactive oxygen species (ROS), and the damage to mitochondria following cryopreservation leads to oxidation of the plasma membrane, DNA fragmentation, and impaired motility. Mitochondria play a pivotal role in regulating sperm physiology during the freeze-thaw process, which can be mediated through a variety of mitochondrial proteins. 1
The uncoupling protein 2 (UCP2) belongs to the mitochondrial anion transporter superfamily that uncouples oxidative phosphorylation and regulates ATP synthesis.2,3 UCP2 plays a crucial physiological role by regulating mitochondrial biogenesis, maintaining energy balance, keeping calcium homeostasis, 4 eliminating ROS, and regulating cellular autophagy.5,6 Thus, it provides cellular protection and possibly has anti-aging properties. 7
To date, limited studies have investigated UCP in sperm or the male reproductive system. Initial studies found that UCP2 is expressed in the spermatozoa of zebrafish and yellow catfish. There is also evidence of a relationship between the quantitative change of UCP2 and ROS levels, as well as post-thaw viability. 8
UCP2 has been reported to be present in human spermatozoa by Wang et al. In another study using mRNA microarray screening, the relative quantity of the newly identified UCP5 and UCP6 in the testis was also predicted. Carrageta et al. screened the mRNA relative quantity of UCP1-6 homologs in human spermatozoa and, for the first time, identified the transcripts of all six UCP homologs—UCP1, UCP2, UCP3, UCP4, UCP5, and UCP6. 9 Wang et al. suggested that the level of UCP2 relative quantity in normospermic men is significantly higher than in asthenozoospermic men and has a relationship with motility.
A recent study used genipin to inhibit UCP, finding it selectively inhibited UCP2 and dose-dependently UCP1 and UCP3 without reducing protein expression.
N-acetyl cysteine is an amino acid derivative of cysteine, developed in the 1950s. It functions as an antioxidant by enhancing the production of glutathione. 10 As a potent antioxidant, it can effectively address infertility in both men and women. The cysteine in glutathione can be reversibly reduced and oxidized, giving it antioxidant properties.11–13 N-Acetylcysteine (NAC) compensates by boosting glutathione production and directly neutralizes free radicals and ROS through its sulfur group. Moreover, this powerful antioxidant facilitates the production of nitric oxide by dilating the arteries, which also have many therapeutic applications with this function. Numerous studies have shown that NAC enhances both sperm quality and quantity by reducing ROS, maintaining membrane integrity, and preventing lipid peroxidation, which is crucial for sperm morphology. 10 NAC increases testosterone levels, promoting sperm health and protecting cell division. It also reduces apoptosis, prevents DNA fragmentation, and boosts sperm viability.
Several studies have examined NAC’s antioxidant effects on mitigating sperm damage from cryopreservation in animals. Partyka et al. (2013) and Michael et al. (2007) found that 5 mM and 1.5 mM NAC increased sperm motility in chickens and dogs, respectively. Ashoori et al. observed improved quality of human sperm post-cryopreservation with NAC treatment. 13 Wang et al. used NAC to reduce mitochondrial ROS levels, showing a link between UCP2 and ROS reduction. 14 Additionally, Sedaghat et al. conducted research in which stress preconditioning of bull sperm was applied by xanthine oxidase before cryopreservation to enhance the functions of thawed sperm. This effect may be mediated through an increase in UCP2 relative quantity. 15 These studies demonstrated that UCP2 relative quantity can be altered by increasing or decreasing oxidative stress and ROS levels. In the present study, we reduced ROS levels by including NAC before freezing sperm to investigate how these changes affect UCP2 relative quantity, as well as the motility and viability of thawed sperm.
Materials and Methods
Sperm collection and analysis
This study was approved by the Ethics Committee of Royan Institute, Tehran, Iran (IR.ACECR.ROYAN.REC.1398.227). Semen samples were collected from 20 men (age between 20 and 40 years) who had been referred to Royan Institute with normal parameters according to WHO 2010 (World Health Organization). 16 All patients provided informed consent prior to their participation in this study, in compliance with institutional and ethical guidelines for research. Semen samples were collected by masturbation into sterile containers after 3–4 days of abstinence. After liquefaction, a computer-assisted sperm analyzing system (CASA) assessed sperm concentration and motility. Papanicolaou staining evaluated normal sperm morphology, with at least 200 spermatozoa examined for abnormalities under a light microscope using immersion oil.
After analyzing sperm parameters with CASA, samples were washed twice with HTF medium (Sodium Chloride 97.8 mM, Potassium Chloride 4.69 mM, Magnesium Sulfate 0.20 mM, Potassium Phosphate, Monobasic 0.37 mM, Calcium Chloride 2.04 mM, Sodium Bicarbonate 25.0 mM, Glucose 2.7 mM, Sodium Pyruvate 0.33 mM, Sodium Lactate 21.4 mM, Gentamicin 10 μg/mL, Phenol Red 5 mg/L) and diluted with 3 mL HTF. Then, samples were divided into four groups: fresh, frozen control, frozen NAC 100 µm, and frozen negative control with 25 µM Genipin (all from Sigma-Aldrich, USA).
Freezing and thawing process
For the freezing procedure, the mentioned concentrations of NAC and genipin above were added separately to sperm suspended in HTF and incubated for 10 minutes at room temperature (RT). Afterward, the commercial freezing medium (Sperm Freeze™, Life Global, Belgium) was added slowly and dropwise in equal volume to the sample and incubated at RT for 10 minutes. Then, the cryovials were placed at 6 cm from liquid nitrogen (−80°C) for 15 minutes and then stored in liquid nitrogen at −196°C. The samples were thawed in a water bath (37°C for 10 minutes) and washed (300 g, 5 minutes) with an equal volume of HTF/10% human serum albumin (HSA, Vitrolife, Sweden) to remove the cryoprotectant. Finally, sperm pellets were resuspended in HTF containing 10% HSA and incubated at 37°C in 5% CO2 for 20 minutes, followed by sperm analysis.17,18
Sperm motility parameters
CASA evaluated kinematic parameters of sperm, including total motility (TM%), progressive motility (PM%), average path velocity (VAP µm/s), straight-line velocity (VSL µm/s), curvilinear velocity (VC µm/s), linearity (LIN%), and straightness (STR%) in five random fields per slide before and after freezing.
Sperm viability
Sperm viability was assessed using eosin-nigrosin staining (0.2 g eosin + 2 g nigrosine in phosphate buffer saline). 10 µL of the stain was mixed with a 10 µL semen sample, left at room temperature for 5 minutes, and smeared on a slide. Under light microscopy at ×100 magnification, 200 sperm per sample were examined across five regions. Nonviable sperm heads appeared red, while viable ones remained colorless.
Western blot analysis
Sperm were washed twice with PBS (Sigma-Aldrich, USA) before extracting total protein using a lysis buffer containing 7 M urea, 2 M thiourea, 4% CHAPS (w/v), 75 mM DTT, 1% ampholyte (w/v), and 40 mM Tris-HCl. Protein concentration was determined using the competitive Bradford method (Thermo Scientific, Rockford, USA). A 20 µg protein sample was loaded onto a 10% SDS‐polyacrylamide gel, electrophoresed, and then transferred to a PVDF membrane (Bio‐Rad, Hercules, USA). The membrane was blocked with 3% bovine serum albumin (Sigma‐Aldrich, USA) for 1 hour at room temperature. It was then incubated overnight at 4°C with primary antibodies against UCP2 (monoclonal anti-UCP2, 1:500, Santa Cruz Biotechnology, Europe) and tubulin (1:1000, monoclonal, Proteintech, USA). This was followed by a 2-hour incubation at room temperature with horseradish peroxidase‐conjugated secondary antibodies. Antirabbit IgG (1:30,000) (Sigma, USA) and antimouse IgG (1:20,000) (Sigma, USA) immunoglobulins were used to detect β‐tubulin and UCP2, respectively. Protein-bound signals were detected using the enhanced chemiluminescence (ECL) detection system (Thermo Scientific, USA). The chemiluminescence signal was captured using gel doc (UVITEC-Cambridge) and results were analyzed using ImageJ software version 1.50i (US National Institutes of Health, Bethesda). Finally, the UCP2 level was normalized to tubulin as a housekeeping protein and then calculated in proportion to the fresh sperm group.19,20
Measurement of mitochondrial membrane potential
JC-1 (Sigma-Aldrich Chemical Co., Germany) was used for the detection of mitochondrial membrane potential (MMP). The spermatozoa samples, at a concentration of 1∼2 × 106 ml, were washed in PBS and incubated with 1 µL JC-1 at 37°C for 15 minutes in the dark. Afterward, sperm cells were washed twice with PBS and analyzed by flow cytometry (BD FACS Calibur; Becton-Dickinson, San Jose, CA, USA). A minimum of 10,000 spermatozoa were examined for each sample at a flow rate of <100 cells/s. In healthy spermatozoa with high mitochondrial potential, JC-1 formed complexes, showing intense red fluorescence (peak emission at 590 nm), which was measured in the FL2 channel (585 nm). For JC-1-stained cells or sperm with low MMP, JC-1 remained in the monomer form, which showed only green fluorescence (peak emission at 527 nm) that was measured in the FL1 channel (530 nm). 21
Measurement of intracellular superoxide anion
Dihydroethidium (DHE; Sigma, USA) is freely permeable to spermatozoa and can directly be oxidized to ethidium bromide by a superoxide anion produced by the sperm cell. DHE was used to assess the levels of intracellular superoxide anion. The spermatozoa samples in concentration 1∼2 × 106/mL were washed in PBS and incubated with DHE (final concentration: 1.25 µM) for 20 minutes in the dark at room temperature. Flow cytometric analysis was conducted using FACS Caliber (BD Immunocytometry Systems, San Jose, CA, USA). The red fluorescence detected was then evaluated between 590 and 700 nm (excitation wavelength 488 nm, emission wavelength 530 nm in the FL-2 channel). All flowcytometric data were analyzed by Flowjo software (v10.5.3, USA). 21
Quantification of lipid peroxidation
Malondialdehyde (MDA), an indicator of lipid peroxidation, was measured via the thiobarbituric acid (TBA) reaction. Thawed straws were incubated with FeSo4 (25 mmol/L) and ascorbic acid (125 mmol/L) at 37°C for 1 hour using 5 × 106/mL sperm. After adding 250 μL of 40% trichloroacetic acid and cooling on ice for 20 minutes, the supernatant was collected, followed by centrifugation (800 × g, 20 minutes). It was then mixed with 500 μL of 2% TBA in 0.2 N NaOH, boiled for 10 minutes, and cooled. TBA-reactive species were detected by spectrophotometry at 532 nm. 22
Statistical analysis
The data distribution was examined using the Kolmogorov-Smirnov test and the Univariate instruction. Then, the data that initially had a normal distribution or obtained a normal distribution following logarithmic transformation were analyzed using the GLM instruction. Data that still did not have a normal distribution despite logarithmic transformation were analyzed using the Kruskal–Wallis test and the NPAR1WAY instruction. The LSMEANS procedure was used for multiple comparisons. All analyses were performed using SAS statistical software, version 9.4. Data were presented as mean ± standard error, and differences at the level of p < 0.05 were considered significant.
Results
Sperm motility and viability
All motility parameters (total motility, progressive motility, VCL, VSL, VAP, LIN, STR, and WOB) (p < 0.01) and viability (p < 0.0001) were decreased after thawing in all frozen groups compared to the fresh control group. However, the frozen NAC group had higher total and progressive motility than the frozen control and frozen genipin group (p < 0.05). In addition, VCL, VSL, VAP, WOB, and LIN in the frozen genipin group were significantly lower than in the frozen NAC group (p < 0.05). Viability was significantly increased in the frozen NAC group compared to the frozen control and frozen genipin group but was significantly lower than in the fresh group (p < 0.05). The percentage of total motility, progressive motility, and viability was significantly higher in the frozen NAC group compared to the frozen control group. (Table 1).
Sperm Motility and Viability: Effect of Adding Antioxidant (NAC) and Inhibitor of UCP2 (Genipin) in Freezing Medium on the Motility and Viability Parameters (Mean ± SEM)
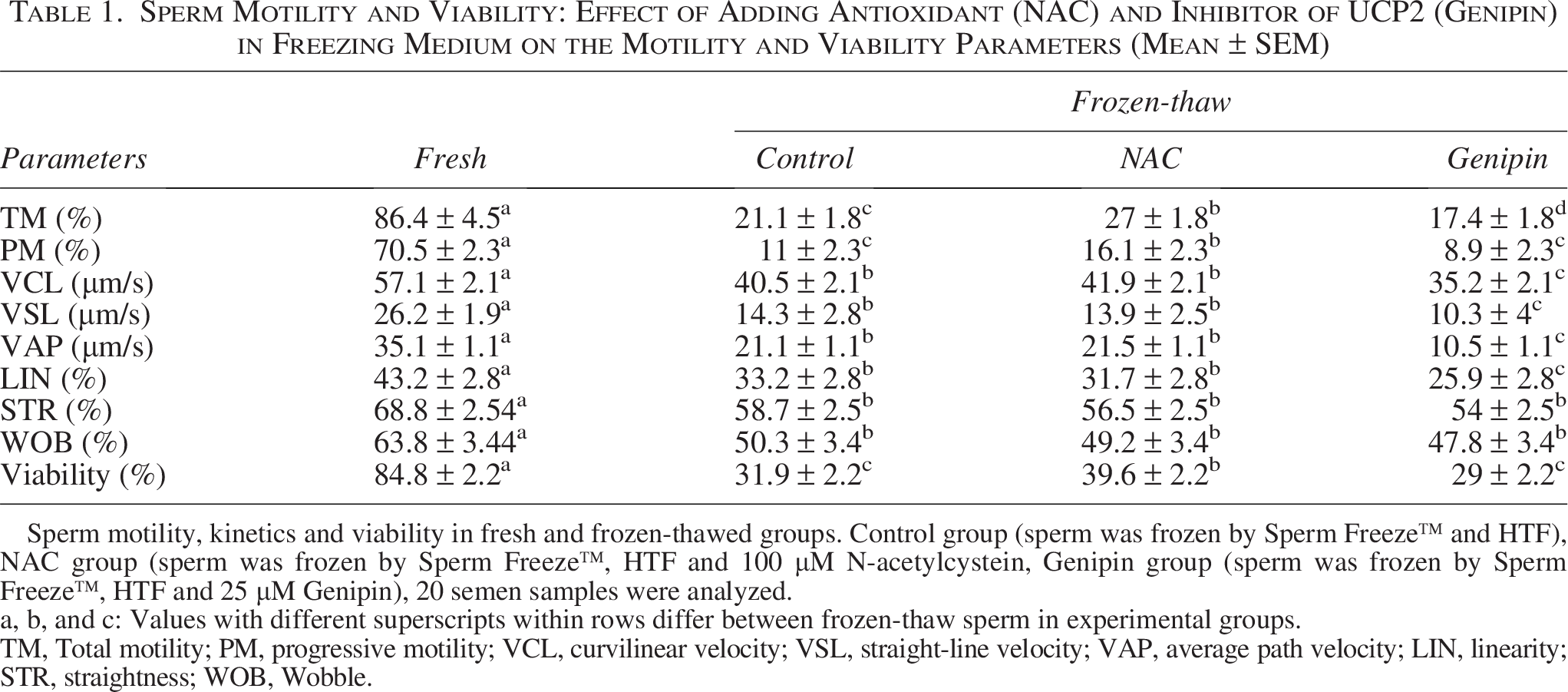
Sperm motility, kinetics and viability in fresh and frozen-thawed groups. Control group (sperm was frozen by Sperm Freeze™ and HTF), NAC group (sperm was frozen by Sperm Freeze™, HTF and 100 µM N-acetylcystein, Genipin group (sperm was frozen by Sperm Freeze™, HTF and 25 µM Genipin), 20 semen samples were analyzed.
a, b, and c: Values with different superscripts within rows differ between frozen-thaw sperm in experimental groups.
TM, Total motility; PM, progressive motility; VCL, curvilinear velocity; VSL, straight-line velocity; VAP, average path velocity; LIN, linearity; STR, straightness; WOB, Wobble.
Malondialdehyde, ROS production, and mitochondrial membrane potential
MDA and superoxide anion did not differ among various groups (p > 0.05). Similarly, the percentage of low and high mitochondrial MMP did not differ among various frozen groups (p > 0.05) (Table 2).
Effect of Adding Antioxidant (NAC) and Inhibitor of UCP2 (Genipin) in Freezing Medium on the MDA, Reactive Oxygen Species Level and Mitochondrial Membrane Potential of Sperms (Mean ± SEM)

Control group (sperm was frozen by Sperm Freeze™ and HTF), NAC group (sperm was frozen by Sperm Freeze™, HTF and 100 µM N-acetylcystein and Genipin group (sperm was frozen by Sperm Freeze™, HTF and 25 µM Genipin). 20 human semen samples were analyzed.
UCP2 relative quantity in protein levels
The level of UCP2 dramatically decreased following the freezing-thawing process (p < 0.0001). Interestingly, the level of protein was remarkably lower in the frozen genipin group (0.08) than in the frozen control (0.29) and NAC (0.32) groups (p < 0.05; Fig. 1 and Fig. 2).

Western blot of UCP2 protein in fresh and post-thaw semen. Tubulin was used as a loading control. Western blot bands showing UCP2 and tubulin protein expression in fresh and post-thawed samples. Control group (sperm was frozen by Sperm Freeze™ and HTF), NAC group (sperm was frozen by Sperm Freeze™, HTF and 100 µM N-acetylcystein) and genipin group (sperm was frozen by Sperm Freeze™, HTF and 25 µM Genipin). Number of samples: 6.

Expression of UCP in fresh and post-thaw semen. Control group (sperm was frozen by Sperm Freeze™ and HTF), NAC group (sperm was frozen by Sperm Freeze™, HTF and 100 µM N-acetylcystein) and genipin group (sperm was frozen by Sperm Freeze™, HTF and 25 µM genipin). Number of samples: 6.
Discussion
We observed a decrease in UCP2 protein relative quantity during sperm freezing and thawing, affecting motility post-freezing. Several studies have evaluated the influence of cryopreservation on the whole sperm proteome in different species such as humans, 14 fish,23,24 boar, 25 and roosters. 26 Cryopreservation affects proteins crucial to sperm metabolism, membrane permeability, flagella structure and movement, apoptosis, signaling, capacitation, and fertilization. Bogle et al. investigated changes in human sperm proteins during the freezing process and observed significant alterations in the sperm proteome at each stage of cryopreservation, which may ultimately impair sperm fertilizing capability. 27 Wang et al. reported that sperm proteins in infertile men decrease after cryopreservation, likely due to degradation from freezing-thawing stress. It is also possible that in the resumption of cell activity, some genes are not expressed due to lack or deficiency of transcription and translation factors. UCP2 plays an essential role in controlling ROS and maintaining energy homeostasis, and cryo-injuries may reduce its levels during the freezing process. 28
The relative quantity of UCP2 in the midpiece of human spermatozoa was previously reported to be higher in normozoospermic men compared to asthenospermic men. Another study found that by inducing sublethal oxidative stress on thawed bull sperm, the UCP2 relative quantity could be increased, thereby improving the functions of the thawed sperm. 15
This study measured UCP2 protein in fresh and frozen-thawed human sperm. Results showed a 3.4-fold decrease in UCP2 during the freezing-thawing process, indicating significant cryo-injury. This finding underscores the delicate balance of proteins crucial to maintaining sperm functionality. Our data indicated that the addition of NAC did not affect UCP2 relative quantity compared to the control group. However, NAC significantly improved total motility, progressive motility, and viability of post-thawed semen, suggesting that antioxidant therapy can be beneficial for preserving sperm quality during cryopreservation. Paralleling these observations, our study also found that genipin inclusion before cryopreservation led to a significant reduction in UCP2 relative quantity, concurrently decreasing total motility. This reduction aligns with the hypothesis that UCP2 plays a critical role in regulating sperm motility through its influence on metabolism and energy content. Although genipin reduced UCP2 relative quantity, no changes were observed in ROS levels or MDA levels, suggesting that the decrease in motility might not be mediated by oxidative stress but rather by metabolic disruptions. NAC is a powerful antioxidant used to treat various diseases for over 30 years. It was observed that NAC in sperm suspended in HTF does not affect UCP2 relative quantity. Conversely, research on myocardial cells demonstrated that NAC regulates UCP2 relative quantity by reducing ROS and preventing severe ATP decreases induced by UCP2. 29
Studies have indicated that UCP2 levels are influenced by ROS concentration. For instance, research suggests that mildly excessive ROS can increase UCP2 relative quantity, whereas very high, lethal levels of ROS lead to a decrease in UCP2 levels. 14
In men with a normal range of ROS in sperm, adding NAC can lead to the inactivation of UCP2. Ryan et al. (2011) discovered that UCP2 is glutathionylated and remains inactive when mitochondrial ROS levels are kept within tolerable limits and the cell’s redox state is normal. Small increases in ROS result in the deglutathionylation and activation of UCP2. 30 Therefore, in this study, measuring the deglutathionylation form of UCP2 would have shown changes in UCP2 relative quantity with NAC inclusion, indicating its effect on UCP2 activation or inactivation. It was also revealed in a study that NAC treatment before freezing sperm enhanced the quality of sperm parameters, decreased the level of reactive oxygen species, and reduced apoptosis. 13 Although ROS levels may affect UCP2 relative quantity, it was found that reducing ROS with NAC antioxidant did not change UCP2 relative quantity in human spermatozoa. In our study, the addition of genipin before cryopreservation resulted in a significant reduction in the quality of sperm compared to the control group. Total motility, VCL, VSL, LIN, and VIP were decreased with the inclusion of genipin, while no changes were observed in MDA and ROS levels. It was noted that with the inclusion of genipin, a decrease in UCP2 relative quantity accompanied a decrease in total motility. Therefore, similar to the findings of Wang et al. (2018), it can be concluded that total motility and UCP2 relative quantity are directly related, although the underlying mechanism is likely not due to changes in ROS levels in human sperm.
Inhibiting UCP3 in human spermatozoa with genipin resulted in a loss of motility without impacting viability, along with a decrease in lactate production. The proposed hypothesis was that the reduced lactate production is linked to decreased ATP production through glycolysis, leading to reduced spermatozoa motility. This research indicates that UCPs act as metabolic modulators of human spermatozoa. Additionally, this study observed no change in ROS levels with the inhibition of UCPs, which is consistent with other data showing no alterations in ROS levels. 9 It was shown that genipin reduces motility by promoting mitochondrial scavenging rather than ATP production, suggesting a regulatory system involving UCP2, mROS, and motility in human spermatozoa. No change in ROS levels or MDA was found with 25 µM genipin, leading to the hypothesis that the decreased motility is due to metabolic control.
Conclusion
The relative quantity of UCP2 protein in post-thaw human sperm was measured, and a significant reduction in its quantity was observed after freeze-thawing. Supplementing the freezing medium with NAC significantly improved the progressive motility and viability of sperm after thawing but did not result in a significant change in UCP2 relative quantity. It is suggested that future studies consider post-translational modifications of UCP2, and the use of infertile samples with abnormal sperm parameters is recommended. Supplementing the freezing medium with genipin shows a direct relationship between sperm motility and UCP2 relative quantity, likely due to changes in sperm metabolism and energy levels.
Authors’ Contributions
P.D. collected experimental data, sample collection, contributed to writing the paper, prepared figures, reviewed and edited the article, interpreted results. L.R., S.A., and V.E. contributed to experimental data collection, interpreted results, and edit the draft. V.A. consulted on statistical analysis. A.S., A.A., and M.S. conceived of the project and developed experimental design, obtained funding, consulted on data, coordinated sample collection, interpreted results, reviewed, and edited the article. All authors reviewed and approved the final draft.
Footnotes
Acknowledgments
The authors thank all participants who participated in the present study. We also would like to extend our appreciation to the Royan Institute clinic and laboratory staff for their sincere contributions to all procedures.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors appreciate the financial support provided by the Royan Institute, Tehran, Iran (Grant Number. 98000008).
Confirmation Statement
The authors declare that Royan Institute is primarily focused on research and education. Royan Institute is a center of excellence in research and technology, a pioneer in the development of science, technology, and innovation of biological sciences