
Abstract
Aim:
Ethylene glycol (EG) has been employed as a cryoprotectant for many years in mammalian semen cryopreservation but not assessed for birds except for its recently illustrated beneficial effects on commercial chicken lines. The Indian red jungle fowl is facing trouble in its native range due to human encroachment. Therefore, the present study was designed to elucidate the cryoprotective effect of different EG concentrations (5%, 10%, 15%, and 20%) on frozen Indian red jungle fowl semen.
Materials and Methods:
Semen was collected from 20 cocks, and qualifying ejaculates (>70% motility) were pooled and diluted (15) with red fowl extender. EG was added to the four samples and 20% glycerol in control at 4°C. Samples were equilibrated and cryopreserved in LN2. Semen quality and biochemical activity were assessed at various stages of cryopreservation.
Results:
Sperm motility, viability, plasma membrane and acrosomal integrity, chromatin integrity, and mitochondrial activity were recorded highest (p < 0.05) with 20% EG at the post-equilibration and post-thaw stages. Lipid peroxidation was recorded lowest (p < 0.05) with 20% EG compared with other concentrations and control at the post-equilibration and post-thaw stages.
Conclusions:
It is concluded that 20% EG exhibits cryoprotective properties in terms of regulating morphological and biochemical traits of frozen Indian red jungle fowl sperm.
Introduction
Ethylene glycol (EG) (C2H6O2) is an organic compound with a molecular weight of 62.07 g/mol that makes it highly permeable and less toxic compared with other compounds utilized as cryoprotectants in freezing practices with respect to germ cells and embryo preservation. 1 It is recently known to replace the most widely used cryoprotectant, glycerol, during semen cryopreservation in some mammalian2–4 and a few avian species.5–7 Glycerol has a higher molecular weight (92.09 g/mol) and viscosity, which causes detrimental changes in frozen spermatozoa 3 by altering the organization and viscosity of cytoplasm and disturbing the structure of phospholipids and proteins in the sperm plasma membrane, ultimately affecting its permeability and durability. 8 Besides harming the sperm structure, glycerol affects the cryopreserved sperm physiology via displaying the contraceptive properties even at concentrations suitable for freeze protection. It could potentially impair sperm transport in the oviduct by changing either the genital tract osmolality, properties of the luminal fluid, or pattern of the cilia motion, eventually limiting access to the ovum perivitelline layer. 9 Hence, all of this requires glycerol removal before utilization in artificial insemination, or it will reduce sperm fertilization capacity. 10
As a result of detrimental changes inflicted by the cryopreservation on sperm physiology, cytoplasm, genome, and sometimes cytoskeleton, employment of an effective cryoprotectant is imperative. 3 During freezing, EG permeates the plasma membrane much faster with higher affinity compared with other cryoprotectants, leading to reduced hydraulic conductivity and osmotic stress in the frozen sperm. This phenomenon maximizes the survival rate of spermatozoa during freeze-thaw. When EG enters the sperm, it causes efflux of water from the cytoplasm to achieve an intracellular stability against low temperatures. 2 However, the effectiveness of cryoprotective compounds depends upon the concentrations that were utilized and the species under study. Different levels of cryoprotectants used during freezing are reported to display wide response variations even among genetically relevant species.10–12
Indian red jungle fowl (IRJF; Gallus gallus murghi), a wild gallus subspecies of South-East Asia, is facing trouble in its native range due to habitat destruction, egg poaching, and predation. 10 Additionally, fowl species are exploited as a food resource globally, 13 which exerts stress on the wild populations, as domestication favors only certain traits, hence disturbing the gene pool. Semen cryopreservation is considered an appropriate procedure for the conservation of such species due to its lower comparative costs when paralleled with in situ management. 14 However, glycerol, most often used for freezing, has been tested in IRJF, and substantial loss in semen quality and fertility was noted.10,15 Furthermore, during the freezing of chicken semen, EG maintained the highest postthaw motility compared with other cryoprotectants, but due to possible differences between breeds/strains, it was suggested to assess for different chicken lines/breeds. 6 Hence, the present study was conducted to evaluate different EG concentrations for quality and biochemical activity of the frozen IRJF semen.
Materials and Methods
All chemicals in this study were purchased from Sigma-Aldrich, Co., St Louis, USA.
Ethical statement
The study was approved by the Ethics Committee of Pir Mehr Ali Shah Arid Agriculture University Rawalpindi, Pakistan.
Experimental birds
Twenty mature IRJF males (age: 3 years) were maintained separately in pens of 106.68 cm × 121.92 cm at an avian research center located in Pir Mehr Ali Shah Arid Agriculture University Rawalpindi, Pakistan. Each bird was offered commercial poultry cock breeder feed (approximately 100 g) daily. Fresh water was offered to the birds ad libitum during the experimental period.
Semen processing
Semen collection was done by the abdominal massage technique. 16 Semen samples with motility of >70% were subjected to further processing. Initial motility of each ejaculate was assessed by mixing 5 µL of semen in 250 µL of phosphate buffer saline (PBS). 17 A drop of sample was placed on a pre-warmed glass slide and observed under a phase contrast microscope at 400× (Olympus, Japan). The sperm concentration of an ejaculate was determined on a hemocytometer. Semen volume was measured on a digital weighing balance by weighing a sample tube before and after the collection. Semen volume of individual cocks ranged from 25 to 30 µL at each collection, and the pooled volume was made up to range 400–500 µL.
Experimental design
Pooled samples were processed for the post-dilution and post-cooling stages of cryopreservation. At equilibration, samples were divided into seven aliquots. The EG was added at the level of 5%, 10%, 15%, 20%, 25%, and 30% to the six freezing samples along with 20% glycerol in the seventh aliquot taken as control. However, post-thaw preliminary results of the samples supplemented with 25% EG and 30% EG showed no motility; hence, these two treatment groups were not used to continue with other sperm quality and biochemical assessments. Five replicates were performed in total for the current study. The sequence of experimental steps has been presented in Figure 1.
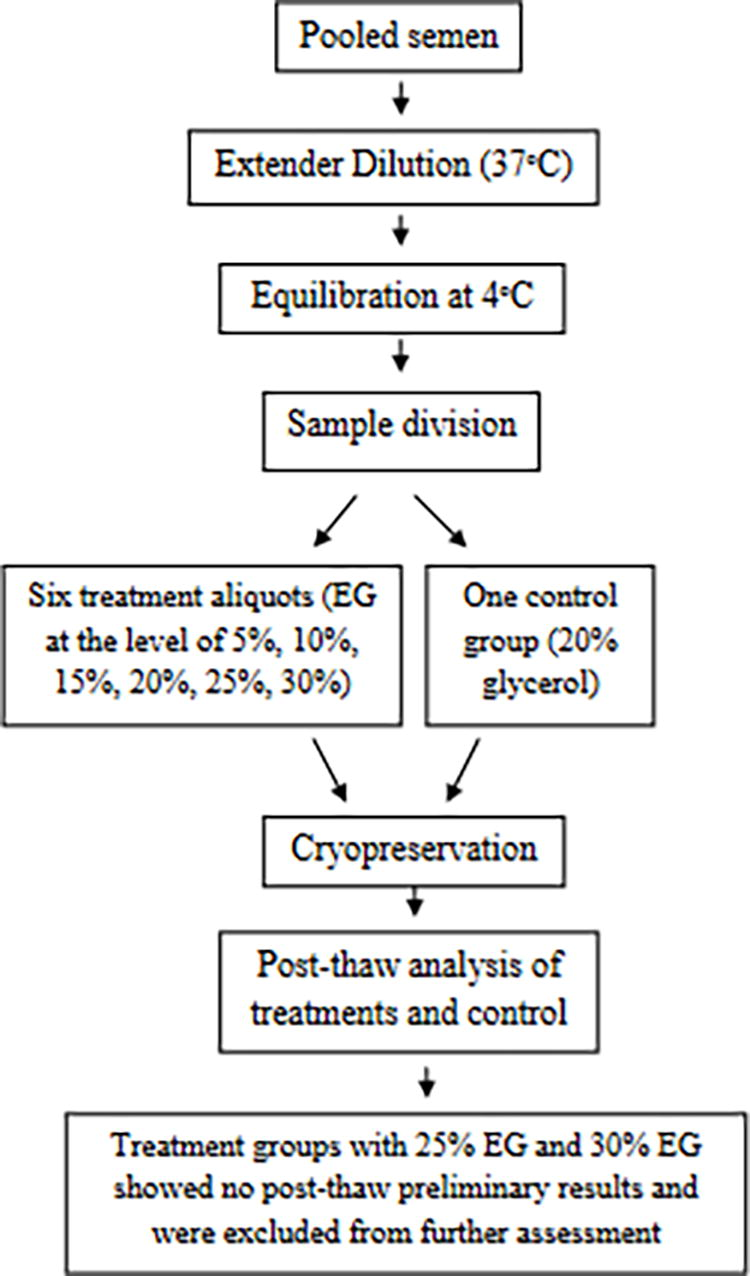
The sequence of experimental steps for Indian red jungle fowl semen cryopreservation with ethylene glycol as a cryoprotectant.
Dilution and freezing process
The red fowl extender (RFE) 18 containing potassium acetate (0.5 g), sodium glutamate (2.1 g), d-fructose (1.15 g), Polyniylpyrrolidone (PVP) (0.6 g), and glycine (0.2 g) in 100 mL distilled water was used as a diluent [pH 7.0; 380 milliosmoles per kilogram (mOsm/kg)]. The pooled sample in each replicate was diluted with RFE (1:5) in a single step at 37°C and processed further. These diluted pooled samples were placed in a cold cabinet till the temperature dropped to 4°C in 2 hours (−0.275°C/min). At the equilibration stage, samples were divided into treatment groups and control as mentioned in the experimental design. All samples were kept again in a cold cabinet for 10 minutes for equilibration. Semen was transferred to 0.5 mL French straws (IMV, France) and placed 5 cm above the liquid nitrogen vapors for 10 minutes. Frozen straws were plunged into liquid nitrogen (−196°C) for storage. After 48 hours, straws were thawed for 30 seconds in a water bath and subjected to post-thaw analysis.
Assessment of EG as a cryoprotectant for IRJF semen
Semen quality
Sperm progressive motility was assessed by placing a drop of semen sample, previously diluted to 1:5 (v/v) in the RFE, on a prewarmed (37°C) glass slide under a phase contrast microscope (×400, Olympus, Japan). The percentage of motile spermatozoa was subjectively evaluated on a scale of 0%–100%. 15
The intactness of the membrane was investigated by hypo-osmotic swelling test (HOST). The Hypo-Osmotic Swelling (HOS) solution was prepared (1 g sodium citrate to 100 mL of distilled water; 100 mOsm/kg); semen samples (1 µL) mixed with HOS solution (20 µL) and a drop of eosin stain were kept for 25 minutes at room temperature. A drop of solution was placed on a prewarmed slide, fixed in buffered 2% glutaraldehyde, covered with a cover slip, and examined under a phase contrast microscope at 1000× (oil immersion, Olympus, Japan). Morphologically normal spermatozoa responded with coiled heads and tails, while dead and abnormal sperm remained straight. A total of 200 spermatozoa was counted. 19
The sperm viability was investigated by Lake’s glutamate solution, which was composed of sodium glutamate (0.01735 g), potassium citrate (0.00128 g), sodium acetate (0.0085 g), and magnesium chloride (0.000676) in 100 mL distilled water. Nigrosin (5 g) and Eosin-bluish (1 g) were added to the Lake’s glutamate solution. The semen sample and stain were mixed, spread on the slide, air dried, and observed under a phase contrast microscope (1000×, oil immersion, Olympus, Japan). The white sperm were considered as normal and live, while pink-stained sperm were taken as dead, with a total of 200 spermatozoa counted. 20
Acrosomal integrity of spermatozoa was evaluated using Giemsa stain. Stain was made by adding Giemsa (3 g) and Phosphate buffered saline (PBS) at pH 7.0 (2 mL) into 35 mL of water. Semen samples were placed on a glass slide and dried. The slide was channeled to neutral formal saline for 30 minutes for fixation and shifted to Giemsa stain for 1.5 hours. Spermatozoa were studied under a phase contrast microscope (1000×, oil immersion). If stained blue, spermatozoa were considered to have intact acrosomes and acrosomal caps, while partially stained or unstained sperm were considered to have abnormal acrosomes. 19
The sperm chromatin damage was assessed by aniline blue stain. The staining solution was prepared by adding 5 g of aniline blue to 100 mL of PBS, filtering the stain, and adjusting its pH (3.5) with the help of 2% glacial acetic acid. A drop of semen was spread on a glass slide and air dried. The slide was stained in aniline blue for 5 minutes, rinsed, and air-dried again. A total of 200 spermatozoa were studied under a phase contrast microscope (1000×, oil immersion). The spermatozoa that were fully or partially stained dark blue were recorded to have chromatin damage, while unstained spermatozoa were recorded for intact chromatin. 19
Biochemical parameters
The 2,2-diphenyl-1-picrylhyrazyl (DPPH) radical is used to evaluate the scavenging activity of samples at an absorption of 517 nm. A working solution was prepared by mixing 3.4 mg of DPPH in 100 mL of 80% methanol (BDH, England). Then, 100 µL of semen sample was added to 1400 µL of DPPH working solution. The mixture was shaken vigorously and incubated at room temperature for 1 hour in the dark to obtain stable absorbance values via spectrophotometer. 21
Detection of lipid peroxidation (LPO) in seminal plasma and spermatozoa is based on the reaction of one Malondialdehyde (MDA) molecule with two NMPI molecules to produce a stable carbocyanine dye showing maximum absorption at 586 nm. For this purpose, 100 µL of sample was mixed with 5 µL of probucol and 320 µL of NMPI (Total Lipid Liquid, Fardiag, Italy) and vortexed. After vortexing, 75 µL of HCl was added, mixed, and incubated at 45°C for 1 hour. Clear seminal plasma and sperm pellet were taken separately by centrifugation (10,000 ×g, 10 minutes). Sperm suspension was set at 50 million sperm for every assessment. The MDA concentration in the seminal plasma was expressed as µM/mL and in the spermatozoa as µM/109 sperm. 22
In the 3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenylytetrazolium bromide (MTT) reduction assay, the Nicotinamide adenine dinucleotide phosphate (NADPH)-dependent cellular oxireductase enzyme depicts the number of viable cells in the sample by reducing tetrazolium dye MTT to insoluble blue formazan. For this purpose, MTT stock solution was prepared by adding 5 mg of MTT in per mL of PBS. Stock solution (5 µL) was added to 50 µL of semen sample. The absorbance was recorded immediately at 550 nm on a spectrophotometer and samples were incubated for 1 hour at 37°C. After incubation, absorbance was recorded again. The rate of MTT reduction was determined by calculating the difference between initial and final absorbance readings. 19
Statistical analysis
Results are presented as means ± Standard Error of Mean (SEM). The data were normalized and the effect of different EG concentrations at the stages of cryopreservation was analyzed by two-factor factorial analysis of variance by using MSTATC® (Version 10.3 Release 3.2.1, J. B. Orris, Butler University, McGraw-Hill Higher Education). Post-hoc comparison was done between means through Tukey’s simultaneous comparison test (when p < 0.05).
Results
Effect of EG on semen quality
The data on the effect of EG in extender on percent sperm motility, viability, plasma membrane and acrosome integrity, and chromatin damage at different stages of cryopreservation are given in Figures 2–6. Overall sperm motility and viability declined (p < 0.05) from the post-dilution to the post-thawing stage of cryopreservation. However, following the addition of EG at the post-equilibration stage, motility and viability were found (p < 0.05) to be higher in 20% EG (68.4 ± 1.6 and 69.6 ± 0.4) as compared with 5% EG (62.0 ± 1.2 and 63.0 ± 0.4), 10% EG (63.2 ± 1.1 and 64.0 ± 0.5), 15% EG (65.6 ± 1.2 and 65.6 ± 0.2), and control, that is, 20% glycerol (65.4 ± 1.1 and 66.0 ± 0.7). A similar trend was observed for motility and viability at the post-thaw stage with the highest motility and viability at 20% EG (45.8 ± 1.9 and 49.2 ± 0.7) compared with other concentrations (5% EG: 40.0 ± 2.7 and 41.0 ± 1.8; 10% EG: 42.6 ± 2.5 and 46.4 ± 0.7; 15% EG: 45.8 ± 1.9 and 49.2 ± 0.7) and control (45.0 ± 1.2 and 50.4 ± 0.3). An overall reduction (p < 0.05) in sperm membrane integrity and acrosomal integrity was recorded from post-dilution to post-thaw stage. At postequilibration, plasma membrane and acrosome integrity was found to be higher (p < 0.05) with 20% EG (67.0 ± 0.7 and 69.4 ± 0.9) compared with 5% EG (60.5 ± 0.9 and 58.6 ± 0.7), 10% EG (62.6 ± 1.0 and 61.0 ± 0.7), 15% EG (64.6 ± 0.7 and 65.8 ± 0.8), and control (64.5 ± 0.9 and 65.6 ± 0.8). Furthermore, the least damage (p < 0.05) to plasma membrane and acrosome was recorded at post-thaw stage for spermatozoa having 20% EG (55.4 ± 0.2 and 54.6 ± 0.4) compared with other concentrations (5% EG: 41.6 ± 1.8 and 41.8 ± 0.9; 10% EG: 44.4 ± 2.2 and 44.0 ± 0.7; 15% EG: 49.4 ± 1.4 and 49.8 ± 0.6) and control (50.4 ± 0.2 and 50.8 ± 0.6). A significant increase (p < 0.05) in the sperm chromatin damage was recorded from post-dilution to post-thaw. Following the addition of EG at post-equilibration, the least chromatin damage (p < 0.05) was recorded in spermatozoa containing 20% EG (12.3 ± 1.0) compared with 5% EG (17.2 ± 0.4), 10% EG (15.2 ± 0.4), 15% EG (14.2 ± 0.5), and control (13.8 ± 0.6). Similarly, at post-thaw the least (p < 0.05) chromatin damage was recorded in spermatozoa with 20% EG (18.3 ± 0.5) compared with other concentrations (5% EG: 25.8 ± 0.4; 10% EG: 22.4 ± 0.2; 15% EG: 19.2 ± 0.4) and control (19.8 ± 0.4).
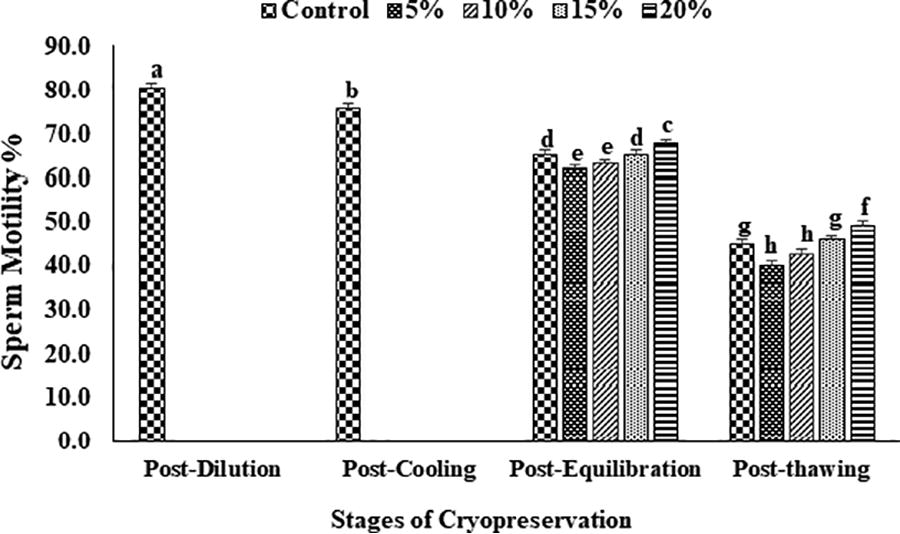
Effect of different concentrations of ethylene glycol (0%, 5%, 10%, 15%, and 20%) in extender on sperm motility (%) of Indian red jungle fowl at post-equilibration and postthaw stages of cryopreservation. Different superscripts indicate statistically significant differences (p < 0.05).

Effect of different concentrations of ethylene glycol (0%, 5%, 10%, 15%, and 20%) in extender on sperm chromatin damage (%) of Indian red jungle fowl at the post-equilibration and post-thaw stages of cryopreservation. Different superscripts indicate statistically significant differences (p < 0.05).
Effect of EG on biochemical activity of semen
The data on the effect of EG in extender on semen scavenging activity (µg/mL), sperm mitochondrial status (nmol/min/109 sperm), and LPO in seminal plasma and sperm at the stages of cryopreservation are given in Table 1, and lower values indicate higher scavenging activity rates. An overall decline (p < 0.05) in the semen scavenging activity and sperm mitochondrial status was recorded at successive stages of cryopreservation. Following EG addition at post-equilibration, the lowest (p < 0.05) scavenging activity was recorded with 5% EG (0.25 ± 0.001), while 20% EG (0.20 ± 0.001), 15% EG (0.22 ± 0.001), 10% EG (0.23 ± 0.001), and control (0.21 ± 0.001) did not differ from each other. However, MTT reduction was recorded higher (p < 0.05) with 20% EG (1.30 ± 0.001) at post-equilibration compared with control (1.18 ± 0.001), 15% EG (1.16 ± 0.001), 10% EG (1.10 ± 0.001), and 5% EG (0.99 ± 0.001). At post-thaw, higher scavenging activity and mitochondrial status (p < 0.05) were recorded in the semen having 20% EG (0.39 ± 0.001 and 1.06 ± 0.001), followed by control (0.43 ± 0.001 and 0.82 ± 0.012), 15% EG (0.44 ± 0.001 and 0.82 ± 0.008), 10% EG (0.51 ± 0.001 and 0.70 ± 0.001), and 5% EG (0.58 ± 0.001 and 0.62 ± 0.000). At post-equilibration, the lowest MDA levels (p < 0.05) were recorded in the spermatozoa and seminal plasma containing 20% EG (1.37 ± 0.000 and 1.20 ± 0.001) compared with control (1.46 ± 0.001 and 1.24 ± 0.001) and other concentrations of EG (5% EG: 1.75 ± 0.000 and 1.34 ± 0.001; 10% EG: 1.58 ± 0.000 and 1.24 ± 0.001; 15% EG: 1.48 ± 0.000 and 1.22 ± 0.001). A similar trend was recorded for spermatozoa and seminal plasma at post-thaw with the lowest MDA levels at 20% EG (1.63 ± 0.002 and 1.43 ± 0.001) compared with other EG concentrations (5% EG: 2.19 ± 0.001 and 1.58 ± 0.001; 10% EG: 1.91 ± 0.001 and 1.53 ± 0.001; 15% EG: 1.80 ± 0.001 and 1.50 ± 0.001) and control (1.79 ± 0.001 and 1.53 ± 0.000).
Effect of Different Concentrations of Ethylene Glycol (0%, 5%, 10%, 15%, and 20%) in Extender on Biochemical Attributes of Indian Red Jungle Fowl Semen at Postequilibration and Postthaw Stages of Cryopreservation
The values with different superscript a to h for a given parameter in rows differ significantly (p < 0.05).
LPO, lipid peroxidation.
Discussion
Assisted reproductive techniques (ARTs) mostly employ sperm cryopreservation and become a robust tool in the conservation plans of wild avian species. However, the lack of comprehensive studies on wild avian reproductive physiology makes freezing a bigger challenge for them compared with domesticated lines. 23 Efficient cryoprotectants are reported to modify water permeability, volumetric and osmotic stress during sperm frosting, and enter the cells quickly with no dependence on the range of temperatures defined for cryopreservation.3,15 EG, having a higher affinity for membranes can act as a practically useful cryoprotectant compared with other routinely used cryoprotectants5–7 and needs further evaluation for wild birds. It is reported to preserve cellular ultrastructures 24 via reducing the ice crystallization levels due to its vitrification properties, which is not a characteristic of most other cryoprotectants. 1 Interestingly, EG freezes at much lower temperatures when mixed with water molecules compared with its pure form, which probably reduces the capacity of intracellular water to transform into crystals and guard the cells from cryo-injuries. 25 Hence, here different EG concentrations were evaluated for the frozen IRJF semen.
In the current study, the addition of 20% EG during cryopreservation maintained higher (p < 0.05) sperm motility, viability, and mitochondrial status compared with 15% EG, 10% EG, 5% EG, and control at post-equilibration and post-thaw. Previously reported cryopreservation of Venda chicken semen with different cryoprotectants revealed EG as effective to improve sperm motility and velocity parameters post-thaw. 5 Another study on cryoprotectants for chicken semen showed beneficial effects of EG on sperm progressive motility and cellular apoptosis at post-thaw. 6 This protective nature of EG primarily relies on its capacity to reduce the likelihood of severe osmotic fluctuations 8 and prevent cells from suffering volumetric changes. As normal mitochondrial functions and ATP production are known to be altered with osmotic shifts19,26 the higher sperm MTT reduction rates, viability, and motility in the current study at post-thaw with EG in the extender compared with the control confirm its ability to aid sperm mitochondrial dehydrogenases in reducing MTT and supplying ATP for the recovery of viable and motile sperm.
Extensively unsaturated lipid content in the avian sperm plasma membrane compared with mammalian sperm increases its vulnerability toward LPO.17,19 Toxic aldehydes and peroxides are byproducts of LPO and generate detrimental changes in the sperm. Peroxidative activity in the plasma membrane results in loss of its own integrity as well as acrosomal integrity with the production of morphological defects, ultimately reducing the sperm’s fertility. 27 In the present study, sperm membrane and acrosome intactness and scavenging capacity were found to be higher (p < 0.05) with 20% EG at post-thaw compared with other concentrations and control. Additionally, 20% EG showed the lowest LPO levels in IRJF sperm and seminal plasma during cryopreservation. Similar findings for EG have been reported in Ghagus chicken semen to achieve acceptable fertilization rates post-thaw. 7 These results indicate the cryoprotective role of EG in maintaining the morphological and physiological stability of frozen semen, including some of its part in balancing scavenging capacity. The lower molecular weight of EG compared with other cryoprotectants allows for immediate permeability within cells and causes the reduction of osmotic lesions.8,28 This faster penetration and exiting of EG minimizes the lethal effects of dehydration and rehydration. 8 With lesser osmotic lesions, less ice crystal formation is observed, and in turn there will be lesser cell damage. This may prevent the malfunction of antioxidants to at least some extent, which is quite evident from the positive effect of 20% EG on the postthaw Reactive Oxygen Species (ROS) scavenging activity. Although the 20% EG did not differ from the control at post-equilibration but differed at post-thaw, it indicates that EG may exhibit an effect on sperm scavenging activity that is amplified at the later stage. Previously, there have been reports of some compounds and substances serving both as cryoprotectants and antioxidants. 29 However, this probable bifunctional role of EG is yet to be defined practically, and future investigations on this aspect could provide novel insights to improve freezing procedures.
Intact chromatin of the sperm exhibits great significance for successful fertilization and preventing deleterious gene mutations. 20 In the current study, the highest chromatin damage (p < 0.05) was recorded in spermatozoa having 5% EG and the lowest (p < 0.05) with 20% EG during freezing. Free radicals are a byproduct of sperm cryostorage, which can produce strand breaks and base modification in the DNA. 30 The levels of ROS production depend upon the extent of disturbance developed in the normal cellular processes.20,26 However, the magnitude of this disturbance could be limited with EG addition as a permeable cryoprotectant that enters into the cells coupled with the efflux of water molecules to prevent ice formation and any possible damage to the nucleus, its DNA, and enzymatic proteins. 31 There are few known studies on mechanisms involved in cryopreservation-related DNA damage, and further studies are needed to elaborate on its importance, which in turn can improve outcomes of ARTs. 30
Sperm volumetric changes are normally regulated via potassium channels against osmotic shock, and EG might affect the working or activation of potassium channels along with fluxes of ions and osmolytes. 2 Nonetheless, each species has its own tolerance range for the supplemented cryoprotectant based on the sperm sensitivity and extent of resistance that semen could exert against freezing to keep redox activity normal.10,18 In the current study, 20% EG showed optimal sperm quality and biochemical activity, while lower EG concentrations showed declined (p < 0.05) seminal attributes, with 5% EG having the worst semen quality. Surprisingly, an increase in the EG concentrations, that is, >20%, did not yield any post-thaw results instead of a gradual decline (mentioned in the experimental design). This may be due to the EG’s strong positive effect on regulating sperm functions, which has also been stated previously. 31 The current data displayed this property in a manner that 20% EG had the highest sperm quality but a decline or increase in its concentrations did not make it better than the control. Moreover, concentrations above 20% EG probably damaged the sperm to the extent where the normal redox mechanism is totally destroyed. This may be due to EG’s likely strong toxicity, which, instead of letting the semen quality fall gradually with the increasing EG concentrations, showed no live sperm at all just after a certain concentration (20% EG). There are several different reasons for different EG concentrations yielding variable results in cryostored sperm of different species to date3,5,6,32,33 with the main reasons being different densities of substances used in diluents, cock age, in vitro storage conditions, number of spermatozoa within each straw, and varying cryopreservation protocols.32,34,35 All these particulars could be worth consideration when interpreting outcomes of various studies performed to explore the appropriate cryoprotectant and to achieve long-term storage of desired genetic resources for future employment in assisted reproduction.
Conclusions
It is concluded that 20% EG addition in RFE extender exhibits cryoprotective properties in terms of regulating motility, viability, plasma membrane and acrosome integrity, DNA integrity, mitochondrial status, and ROS production in frozen IRJF sperm. Furthermore, to some extent EG addition showed improvement in the antioxidative scavenging activity at post-thaw, which, if further investigated, may provide new insights into its bifunctional role, if it exists. Although the extent of EG’s positive effects varies among species/breeds and needs a separate investigation, it came out to be superior in maintaining IRJF semen quality.
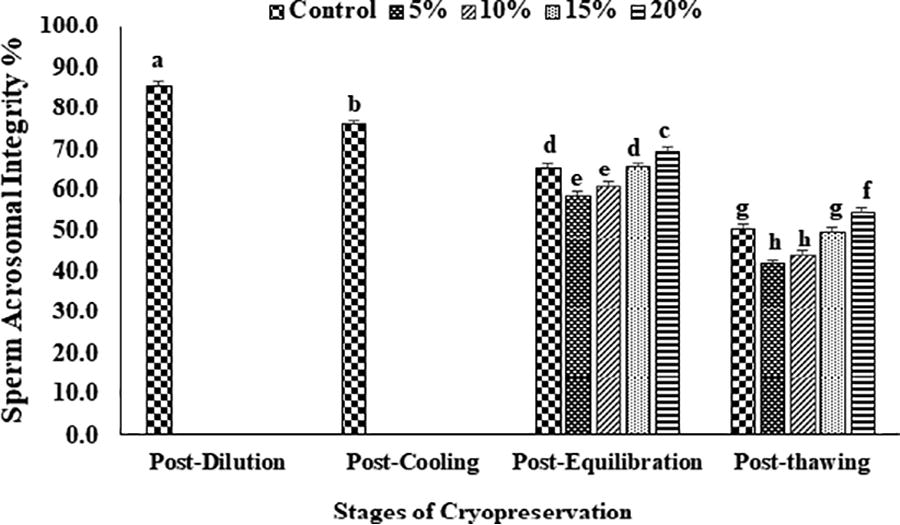
Effect of different concentrations of ethylene glycol (0%, 5%, 10%, 15%, and 20%) in extender on sperm acrosomal integrity (%) of Indian red jungle fowl at post-equilibration and post-thaw stages of cryopreservation. Different superscripts indicate statistically significant differences (p < 0.05).
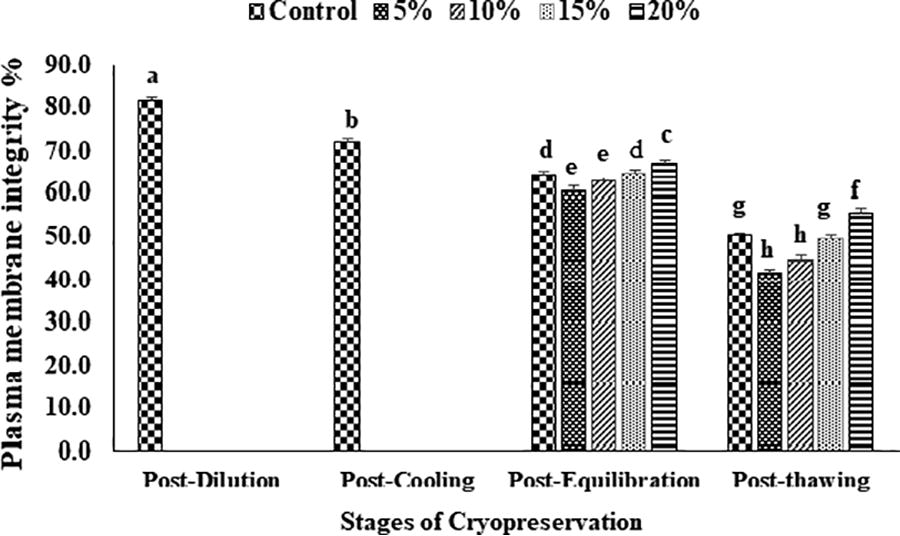
Effect of different concentrations of ethylene glycol (0%, 5%, 10%, 15%, and 20%) in extender on sperm plasma membrane integrity (%) of Indian red jungle fowl at post-equilibration and post-thaw stages of cryopreservation. Different superscripts indicate statistically significant differences (p < 0.05).
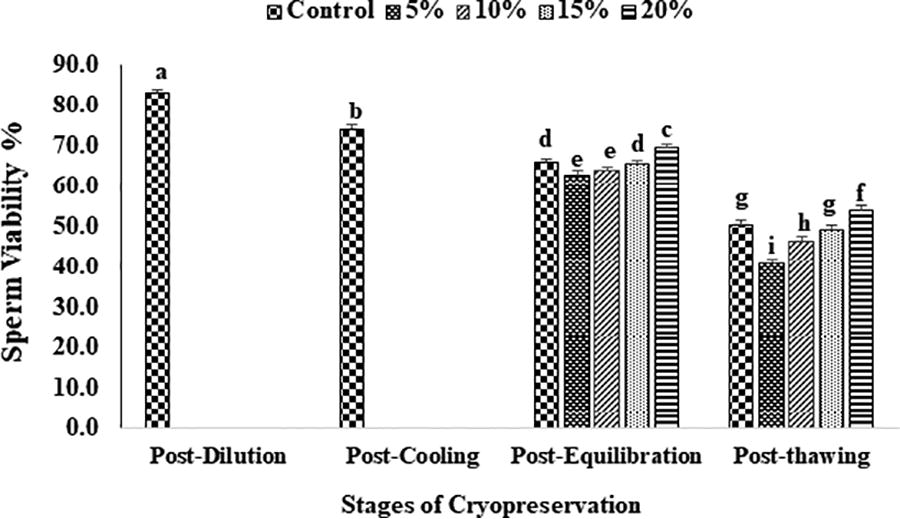
Effect of different concentrations of ethylene glycol (0%, 5%, 10%, 15%, and 20%) in extender on sperm viability (%) of Indian red jungle fowl at the post-equilibration and post-thaw stages of cryopreservation. Different superscripts indicate statistically significant differences (p < 0.05).
Footnotes
Authors’ Contributions
F.K.: Management of study animals (lead), investigation (lead), methodology (lead), data acquisition (lead), formal analysis (lead), and validation (supporting). B.A.R.: Conceptualization (lead), project administration (lead), methodology (lead), supervision (lead), writing—review and editing (equal), and validation (lead). S.Z.: Management of study animals (lead), data acquisition (lead), formal analysis (lead), writing—original draft (lead), writing—review and editing (lead), and validation (supporting). M.S.A.: Data curation (lead), validation (supporting), formal analysis (lead), and writing—review and editing (equal). S.A.: Conceptualization (lead), resources (lead), visualization (lead), validation (lead), and writing—review and editing (equal).
Author Disclosure Statement
There are no disclosures.
Funding Information
No funding was provided for this research.