
Abstract
Introduction:
Human mesenchymal stromal cells (MSCs) are attractive for both medical practice and biomedical research. Nonfreezing short-term storage may provide safe and simple transportation and promote the practical use of MSCs.
Objectives:
We aimed to determine the duration of efficient storage at ambient temperature (22°C) of human dermal MSCs in different three-dimensional organization and to investigate the role of cell metabolic mode in the resistance to the ambient storage damaging factors.
Methods:
MSCs in monolayer, suspension, and encapsulated in alginate microspheres (AMS) were stored in sealed containers at 22°С in culture medium. Viability (fluorescein diacetate /ethidium bromide) and metabolic activity (Alamar Blue assay) were assessed at 0, 3, 7, 10, and 14 days of the storage. Mitochondrial membrane potential (JC-1 test), cell cycle analysis, reactive oxygen species level, and resistance to hydrogen peroxide were analyzed under culture conditions.
Results:
Alginate encapsulation was shown to maintain viability (about 85%), metabolic activity, and adhesion ability during storage for 7 days. The storage of MSCs in both monolayer and suspension was less efficient. Culture of MSCs in AMS decreased basal metabolic activity, mitochondrial activity, and led to reversible cell cycle arrest compared to standard two-dimensional culture. MSCs in AMS have a lower basal level of reactive oxygen species and higher resistance to hydrogen peroxide compared with those in monolayer culture.
Conclusion:
Revealed shift into quiescent metabolic mode is essential for alginate-encapsulated MSCs resistance to storage at ambient temperature.
Introduction
Human mesenchymal stem/stromal cells (MSCs) are a promising cell type for clinical medicine and biomedical research due to their unique properties. MSCs can be isolated from a variety of tissue sources.1,2 Besides the generally accepted osteogenic, adipogenic, and chondrogenic lineages, 3 MSCs have been shown to differentiate into myocytes, cardiomyocytes, 4 hepatocytes,5,6 and neural cells.7,8 High therapeutic potential of MSCs is supported by the secretome with angiogenic,9,10 immunomodulatory, 11 and growth stimulatory action. 12 Practical use of MSCs requires effective preservation technologies. The only established approach for long-term storage is cryopreservation. However, it has limitations such as the need for expensive equipment and liquid nitrogen for storage and transportation, its effects on the cytoskeleton, 13 the differentiation potential, 14 and the gene expression rate. 15 Various approaches to reduce dimethyl sulfoxide concentration have been investigated,16,17 but it is still generally accepted that the negative effects of cryopreservation should be mitigated. The development of nonfreezing storage of MSCs is highly anticipated due to its simplicity and safety. It has been shown that some cell types may be stored under hypothermic conditions, but the data are contradictory. Thus, storage of MSCs in a monolayer at 4°C for 18 hours to 3 days resulted in increased apoptosis and necrosis. 18 The study 19 revealed that induced pluripotent stem cell-derived retinal pigment epithelium cells stored in suspension at 37°C exhibited the lowest viability due to hypoxia; those preserved at 4°C were damaged via microtubule fragility, while cells stored at 16°C were optimally preserved up to 6 hours. It was found 20 that the survival rate of MSCs in suspension stored for 6 hours at room temperature was lower compared to storage at 4°C. Conversely, the study 21 demonstrated that MSCs stored in suspension for 6 hours at 4°C lost their viability. Several studies22–24 have shown alginate encapsulation to be a promising approach for effective storage of MSCs at ambient temperatures. The mechanisms of alginate encapsulation protective effect are still not clear. Mahler and colleagues 25 hypothesized that alginate gel could provoke integrin ligation disrupting the integrin–caspase complex, thereby preventing integrin-mediated apoptosis. It is also widely accepted that alginate entrapment provides protection against mechanical stress. 26 However, the impact of encapsulation into alginate hydrogel on cellular metabolism remains poorly investigated.
We have chosen high-voltage electrospraying 27 for MSCs encapsulation because it operates in a controlled manner. Among the existing types, we focused on human dermal MSCs due to the notably less invasive tissue collection procedure compared to bone marrow and adipose tissue. Dermal MSCs possess similar multilineage differentiation potential and even higher growth and angiogenic capacities compared with bone marrow-derived MSCs. 28 Previously, we demonstrated that human dermal MSCs are positive for CD29, CD73, CD90, and CD105 and negative for CD34 and CD45, 29 and that alginate encapsulation supports their viability and differentiation potential. 30
Herein, we integrated our knowledge of MSC biology with expertise in cryotechnology to devise a freezing-free protocol for short-term MSCs storage. Additionally, we investigated the role of metabolic mode in cell resistance against storage-related damage. The design and idea of the study are shown in Figure 1.

The design and idea of the study.
Materials and Methods
Human skin-derived mesenchymal stromal cell isolation and culture
The MSCs were isolated by the explantation method, 31 following the written informed consent of the donor in accordance with the Helsinki Declaration of the World Medical Association for Biomedical Research and the requirements of the IPC&C Ethics Committee. Cells were cultured in adhesive culture flasks (TPP, Switzerland) at 37°C, 5% CO2, and 95% humidity in minimal essential medium-α modification (α-MEM, Biowest, France) with 10% fetal bovine serum (PAA, Austria), 50 μg/mL penicillin (Biowest, France), 50 μg/mL streptomycin (Biowest, France), and 0.2 mM l-glutamine (Sigma-Aldrich, USA). MSCs at passages 4–8 were used for experiments.
MSCs encapsulation in alginate microspheres
Alginate sodium (Sigma, 71238) 2% solution was mixed with a cell pellet (0.5 mL per 1 × 106 cells). A high-voltage encapsulation device and a 0.4 mm diameter blunt cannula (100 Sterican®, B.Braun Melsungen, Germany) were used to disperse the mixture into a 100 mM solution of CaCl2 via electrospaying. Alginate microspheres (AMS) were allowed to cross-link for 5 minutes, then stepwise washed out with Hank’s balanced salt solution (HBSS, Sigma). During the final washing-out step, the saline solution was replaced with a culture medium.
Preparation of samples with different MSC 3D-organization
To obtain a monolayer, cells were cultured for 5 days, detached by trypsinization, washed, seeded in 96-well plates at 12 × 103 cells/well, and cultured overnight for adhesion. One plate was then analyzed, while the remaining plates were stored or cultured. For the suspension group, cells were cultured for 6 days, detached from the flask, samples were taken for analysis, and the remaining cells were stored.
To prepare AMS, cells were cultured for 5 days on a flask, detached from the flask, encapsulated, and cultured overnight in AMS for adaptation to encapsulated state, then samples were taken for analysis, and the remaining AMS were stored or cultured.
Storage at ambient temperature
MSCs in monolayer, suspension, and encapsulated in AMS were stored in sealed containers at 22°С in culture medium in a cooling thermostat LAUDA (LAUDA, Germany). Viability and functional state parameters were assessed before storage and at days 3, 7, 10, and 14 of storage (n = 3 in triplicate for each group).
Viability assessment
Viability was assessed by double staining with fluorescein diacetate (FDA)/ethidium bromide (EB). 32 Fluorescent dye incorporation into cells was evaluated using LSM 510 META confocal laser scanning microscope (Carl Zeiss, Germany). Confocal images were obtained along the Z-axis with 20 μm intervals at an excitation wavelength of 488 nm for FDA and 543 nm for EB.
Metabolic activity assessment
The tested samples were incubated for 2 hours with culture medium containing 10% Alamar Blue (AB, Serotec Ltd., Bio-Rad, USA). The fluorescence of the reduced form of AB was measured using a Tecan GENios microplate reader (Tecan GENios, Austria) at an excitation wavelength of 550 nm and an emission wavelength of 590 nm and expressed in relative fluorescence units (RFU).
Attachment efficiency
The attachment efficiency was assessed in such a way: cell suspensions were seeded in 24-well plates (for the AMS group, cells were first extracted from alginate hydrogel using citrate solution), after 24 hours of culture the number of unattached cells was counted using a hemocytometer. Attachment efficiency was calculated using the formula:
Haematoxylin–Eosin staining
Samples were stained as described by Fischer et al. 33 and were examined using inverted microscope CETI EpiFluor (CETI, Belgium).
Mitochondrial activity (JC-1 test)
Samples were washed with HBSS and then incubated for 30 minutes with fluorescent cationic dye JC-1 at a final concentration of 2 μM. The fluorescence of J-aggregates was measured at an excitation wavelength of 550 nm and an emission wavelength of 590 nm, and the fluorescence of JC-1 monomer was measured at an excitation wavelength of 514 nm and an emission wavelength of 535 nm using a TECAN GENios microplate reader. The red/green fluorescence ratio was used as an index of mitochondrial activity.
Cell cycle analysis
MSCs in monolayer were cultured using Premo™ FUCCI Cell Cycle Sensor kit reagents for 24 hours. The cells were then collected, one half was seeded on tissue culture plastic, the rest was encapsulated in AMS. Live-cell imaging of cell cycle progression was performed using an Olympus FV10i-LIV confocal laser scanning microscope.
Reactive oxygen species level and resistance to hydrogen peroxide-induced oxidative stress
Reactive oxygen species (ROS) level was measured using Cellular ROS Assay Kit Deep Red (Abcam). Basal ROS level was assessed in MSCs cultured for 24 hours. MSCs cultured for 24 hours in monolayer or in AMS were exposed to 3 mM hydrogen peroxide solution for 2 hours to induce oxidative stress, 34 washed out with HBSS, and induced ROS level was determined. Fluorescence of ROS sensor was detected using confocal laser scanning microscope Olympus FV10i-LIV and Olympus cellSense Software.
Statistics
The obtained results were statistically processed. The normality of the data distribution was determined by the Shapiro–Wilk test. The significance of differences between groups was examined by Student t-test for independent variables. Data were represented as M ± m; differences were considered significant at p < 0.05.
Results
The viability of MSCs before storage was over 98% in all groups (Fig. 2a). In AMS, cell viability remained higher throughout the entire storage period compared to the other groups. Over the course of 7 days, viability exceeded 70%, meeting FDA requirements. 35 In monolayer storage, viability remained sufficiently high (above 70%) for 3 days, but significantly declined with further storage. The lowest viability was observed in suspension storage, dropping below 70% after 3 days of storage.
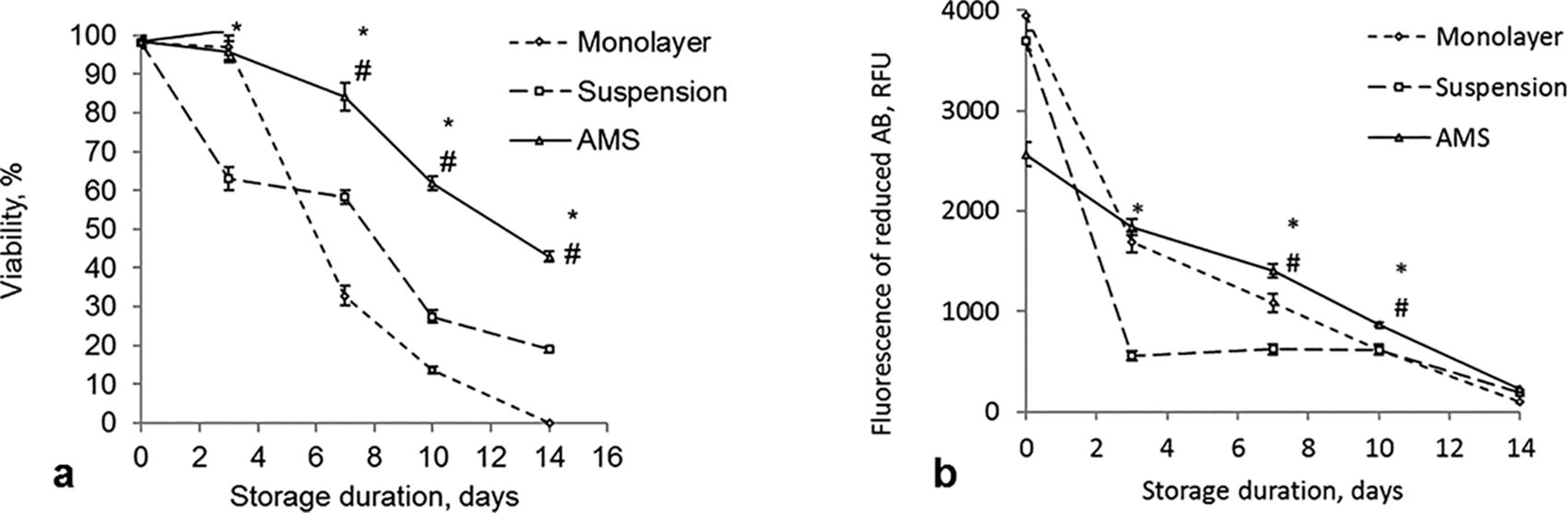
Viability
The metabolic activity of MSCs in AMS was 30% lower after 24 hours of culture before the storage compared to the other two groups (Fig. 2b, 0 day). However, during the storage, metabolic activity in the AMS group was supported in the most efficient manner. When stored in monolayer, metabolic activity of MSCs decreased by 50% by day 3 and continued to decline by 20%–25% every subsequent 3 days. The lowest metabolic activity was observed in cells stored in suspension, decreasing to nearly 15% of the initial level within the first 3 days.
To assess the impact of storage on the self-renewal potential of MSCs, we examined attachment efficiency and proliferation ability before and after 7 days of storage. Prior to storage, all groups exhibited high attachment efficiency (>99%). However, following 7 days of storage, a significant difference emerged. While MSCs stored in AMS maintained a high attachment efficiency (64.0 ± 3.6%) and displayed normal morphology, cells stored in suspension showed significantly reduced attachment (12.0 ± 0.8%) (Fig. 3b, c). Moreover, the lower attachment efficiency of suspension-stored MSCs correlated with impaired proliferation. Monolayer-stored MSCs were almost completely detached after 7 days (Fig. 3a), rendering them unsuitable for further analysis. These findings suggest that encapsulation in AMS efficiently preserves the self-renewal potential of MSCs after 7 days of storage.
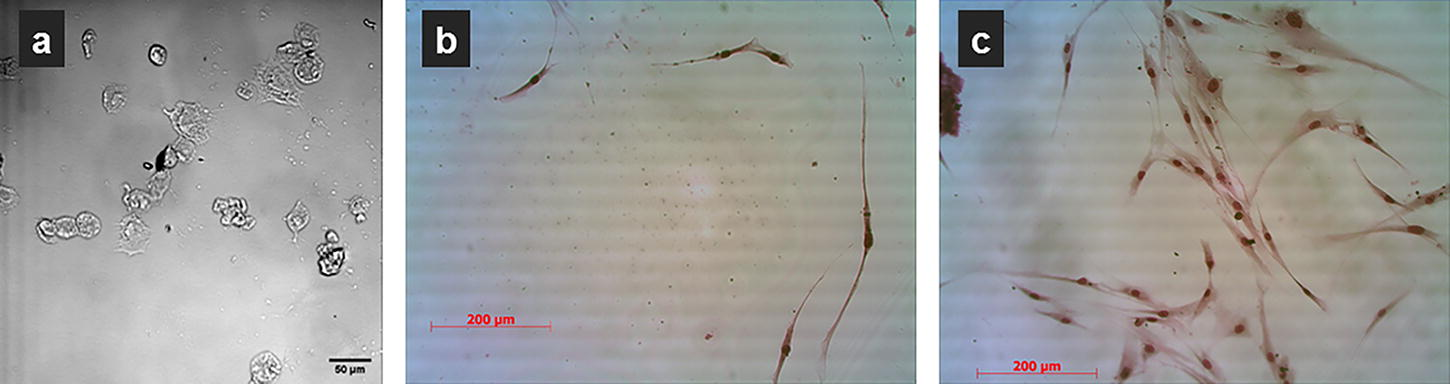
MSCs after 7 days of storage at ambient temperature: detached cells in stored monolayer
Thus, we have revealed that alginate encapsulation supported viability and self-renewal potential of MSCs stored at ambient temperature but decreased their basal metabolic activity in the first 24 hours before storage. Therefore, we decided to further investigate how alginate encapsulation and culture in AMS affect metabolic mode, cell cycle, and ROS metabolism as key factors in MSC resistance to storage-induced damage.
The mitochondrial activity of MSCs after culture in monolayer or in AMS for 3 days was assessed using JC-1 dye, which exhibits green fluorescence in its free form and red fluorescence in the form of J-aggregates. 36 It was revealed that the red/green JC-1 fluorescence ratio was 1.47 for MSCs in monolayer and 0.5 for MSCs cultured in AMS. Considering that a higher mitochondrial potential results in more J-aggregates, it can be concluded that alginate encapsulation decreased mitochondrial activity.
For cell cycle analysis MSCs were transduced with Premo™ FUCCI Cell Cycle Sensor constructs, encoding red (RFP) and green (GFP) fluorescent proteins fused to Cdt1 and geminin. In the G1 phase, only Cdt1-RFP is detectable, rendering cells red. Conversely, during the S, G2, and M phases, only geminin-GFP is expressed, resulting in green cells. The G1/S transition exhibits both Cdt1 and geminin, leading to yellow nuclei. Live imaging was utilized to visualize the cell cycle in MSCs growing in two-dimensional (2D) monolayer and in AMS 3D culture. Cells transduced with different phases of the cell cycle, predominantly in S, G2, and M phases (green cells), were either seeded on tissue culture plastic or encapsulated in AMS (Fig. 4, 1 hour of culture). During subsequent culture, MSCs were mainly in S, G2, and M phases in the monolayer, if sufficient space was available, while those encapsulated in AMS became arrested in the G1 phase 48 hours later (Fig. 4).

Live-cell imaging of cell cycle progression during culture of MSCs transduced with Premo™ FUCCI cell cycle sensor constructs in monolayer and AMS: green—S, G2, and M phases; red—G1 phase; and yellow—G1/S transition.
Basal intracellular ROS levels, assessed as real-time ROS sensor fluorescence, were 32.8 ± 5.2 and 2.32 ± 0.25 RFU/cell in monolayer and in AMS cultured for 24 hours, respectively. Following a 2-hour exposure to 3 mM hydrogen peroxide, ROS sensor fluorescence increased to 154.5 ± 7.8 RFU/cell in monolayer culture and 11.78 ± 0.44 RFU/cell in AMS. Therefore, encapsulation reduced basal ROS levels and enhanced MSC resistance to oxidative stress.
Discussion
The optimal storage method for MSCs is still uncertain. Currently, cryopreservation is the gold standard, though the procedure and cryoprotective agents’ toxicity impact MSCs. 37 Nonfreezing storage is a promising alternative, but an optimal protocol is yet to be developed. Early studies showed hypothermic temperatures (2–10°C) were more favorable than room temperature for short periods (6–24 hours) for MSC suspensions.38,39 Later, hypothermic and ambient temperatures were found to be similarly effective for storage.40,41 To prolong viability for an extended period, two strategies affecting metabolism proved effective: MSC suspension storage in specific solutions21,42,43 or storage in 3D constructs like alginate hydrogels.23,24,30,34,44,45 Given that “low” hypothermic temperatures suppress enzymatic systems responsible for antioxidant defense, genome and cell structure repair, and homeostasis, we opted for storage at ambient temperature. We then compared the effects of different 3D MSC organizations on storage efficacy. Our findings showed that encapsulation maintained high MSC viability for 7 days at 22°C, meeting the cell therapy standards of the US Food and Drug Administration. Encapsulation preserved adhesion, metabolic activity, and proliferation of MSCs stored for 7 days, whereas monolayer or suspension storage resulted in viability loss.
The next step was to identify mechanisms responsible for storage resistance. While previous studies23,24,30,34,44,45 also demonstrated the successful storage of MSCs in alginate hydrogels at ambient temperature, our study provides additional insights into underlying mechanisms of encapsulated cell resistance by highlighting the contribution of metabolic mode into enhanced storage capacity. We analyzed the changes in the metabolic pillars of cell functionality following encapsulation and subsequent culture. We observed a reduction in MSC metabolic activity 24 hours after encapsulation, a finding also reported by other researchers. 45 Using the JC-1 test, we detected decreased mitochondrial activity in MSCs cultured in AMS compared to monolayer culture. These findings align with results 46 showing higher glycolytic activity and lactate production in MSCs within nonadhesive alginate hydrogel compared to modified hydrogels that promote cell adhesion. The unmodified alginate used in our study limited interactions, resulting in round-shaped encapsulated cells with reduced metabolic and mitochondrial activities.
MSC proliferative behavior is influenced by 3D cell organization. We observed cell cycle arrest in the G1 phase 48 hours after encapsulation in AMS, corroborating findings 47 that culture in alginate hydrogel suppresses MSC proliferation. Low metabolic activity and reversible cell cycle arrest are key features of the natural quiescence state, which protects adult stem cells from damage and preserves stemness.48,49 Similarly, resistance to nonfreezing storage may result from the quiescence induced by alginate encapsulation. Slight changes in alginate concentration and cross-linking conditions did not appear to affect the development of resistance to nonfreezing storage in encapsulated cells. Successful ambient storage of MSCs in alginate hydrogels has been demonstrated in both our study and others, using calcium-cross-linked hydrogels with alginate concentrations ranging from 1.2% to 2.2%34,45 as well as strontium-cross-linked alginate hydrogel. 23
The damaging factors in hypothermic preservation include ion imbalance, pH shift, oxidative stress, and endoplasmic reticulum stress,18,50 with oxidative stress becoming most critical under hypoxia. Evidence shows increased ROS generation during hypothermic storage and rewarming.51–53 Given the decreased mitochondrial activity in encapsulated MSCs, we anticipated a lower basal ROS level, as the electron transport chain is the main ROS producer. Analysis using the intracellular ROS sensor deep red fluorescence confirmed a significantly reduced basal ROS level in encapsulated MSCs compared to monolayer. Lower basal ROS levels indicate a more balanced redox state, which can enhance the cell’s capacity to withstand oxidative stress. We actually found increased resistance to oxidative stress, with ROS levels after hydrogen peroxide exposure significantly lower in cells in AMS than in monolayer. Alginate polymers were shown to scavenge-free radicals 54 and suppress oxidative stress-induced apoptosis.55,56 Mechanisms behind enhanced ROS resistance may be attributed to both the protective effects of alginate and metabolic adaptations induced by the microenvironment within the alginate matrix and need to be deeper delved in future studies. Our findings of changes in ROS metabolism in encapsulated MSCs have positive implications for enhanced cellular component protection and oxidative stress response during ambient storage.
Conclusion
In conclusion, we demonstrated the high survival rate of alginate-encapsulated human skin-derived MSCs during storage at 22°C in sealed containers for up to 7 days. Alginate encapsulation resulted in decreased basal metabolic activity, mitochondrial activity, and basal levels of ROS, leading to reversible cell cycle arrest. We also observed increased resistance to hydrogen peroxide-induced oxidative stress in encapsulated MSCs. Therefore, the process of alginate encapsulation initiates a series of metabolic changes that significantly contribute to cell resistance during hypothermic storage at ambient temperature and under hypoxic conditions.
Footnotes
Authors’ Contributions
O.P. and N.T. conceived of the study concept and design. N.T., O.H., Y.K., and O.T. carried out the experiments. Y.K., I.K., and K.K. participated in technical assistance. N.T., Y.K., and O.T. performed the statistical analysis. N.T., O.P., and O.T. took the lead in writing the article. All authors provided analysis of results, critical feedback, and helped shape the research and the article.
Author Disclosure Statement
The authors have no relevant affiliations or financial involvement with any organization with a financial interest in or financial conflict with the subject matter discussed in the article.
Funding Information
This study was supported by the National Research Foundation of Ukraine (project no. 2021.01/0276) and the National Academy of Sciences of Ukraine (project 2.2.6.145/no. 6 23.09.2021§ 24-5).