
Abstract
Hypothesis:
If double stranded DNA (dsDNA) is a charged biomolecule that moves in Earth's magnetic field at a Brownian velocity, then dsDNA may emit bioelectromagnetic waves at energies that reflect discrete genetic states.
Methods:
This work leverages the Planck-Einstein-de Broglie relationship and applies this concept to Brownian velocity of dsDNA within a cell, to describe the relationship between dsDNA mass, the average Brownian velocity of dsDNA within a cell, and the theoretical wavelengths at which DNA may emit bioelectromagnetic waves.
Results:
Theoretical emission wavelengths of dsDNA, derived from first principles, were found to correlate closely with experimentally observed emission wavelengths from spectroscopic measurements across various cellular systems in the literature.
Conclusion:
This work provides a conceptual basis for the potential for unification of bioelectromagnetism with Brownian motion, to elucidate how electromagnetic information can be generated at a subcellular level in biological systems. The implications of how finite mass changes in dsDNA can result in discrete emission wavelengths on electromagnetic timescales is discussed through the lens of genomics. Future refinements of this fundamental methodology may provide a conceptual basis to address previously unexplained multilevel phenomena in the field of biology and is general enough to be extended to other charged biomolecules at a subcellular level. Further exploration in this area could lead to new biological tool development that may augment current genomics methods.
Introduction
Much is known about the chemical conditions for life, although much less is known about the electromagnetic requirements for life. Biological life evolved on Earth in the presence of abiotic forces such as electromagnetic fields and a wide array of sensitivities are conserved across biological space. While there has been much focus on the negative effects of biochemical perturbations associated with the energetic deposition from external electromagnetic radiation at many levels of granularity, 1 such research has not led to a complete model of how electromagnetic forces directly modulate biological responses.
The most common example of how electromagnetic depositions modulate biological responses is when cells absorb external ionizing radiation from the environment. In general, it is a commonly accepted theory that ionizing radiation modulates genomic stability and subsequently alters phenotype within an organism. However, a conceptual boundary exists in the fact that we cannot currently predict all relevant biomolecular change involved in mutagenic phenomena associated with external electromagnetic fields that is based upon assumed reconstruction of properties within individual genes. 2 There is a large body of experimental methodology that exposes healthy cells to finite regions of the electromagnetic spectrum and then analyzes biomolecular changes. However, such reductionist experimental methods often rely on chemical principles alone and exclude physical properties of subcellular components that may also contribute to biological effects.
External electromagnetic forces are often explained as a stressor to cells and various chemical theories have been proposed to develop mechanistic understanding for such “stress.” For example, the primary damaging effect of ionizing electromagnetic deposition to cells is often attributed to the radiolysis of water. 3 Radiation-induced radical oxygen species (ROS) are known to alter the chemical makeup of biomolecules (i.e., lipids, proteins, nucleic acids), although it is not theoretically or mechanistically clear how finite concentrations of ROS modify biomolecules or modulate eustress at a systems level. For example, despite a widely accepted chemical theory of how increased ROS concentrations contribute to increased oxidative stress in biological systems, experimental evidence suggests that radical scavenging therapies contain little if any clinical efficacy for treatment of disease states associated with oxidative stress.4–6 It is not in debate that radical species serve functional roles in transferring bioelectronic information through electron transfer reactions within cells, 7 but new theory is required to develop a more complete understanding of how electromagnetic waves may modulate biological systems beyond biochemical concepts of oxidative stress.
Conceptually, chemical theories often imply that ROS are sufficient to explain bioelectronic states of organisms through electron transfer reactions between biomolecules. It is often overlooked that electromagnetic fields can shift chemical enzyme kinetics and can induce altered bioelectronic states that involve electron transfer reactions between subcellular components.8,9 Small-molecule electron transfer reactions in biological systems have been widely utilized to obtain redox information that is related to stress and/or signaling events. However, there is a current lack in theory to delineate how electron transfer processes contribute to oxidative stress and/or signaling events that arise in cells due to external electromagnetic exposures. This may be due to both the lack of theory on and experimental quantification in support of how cells directly utilize electromagnetic fields to transmit complex bioelectronic information and direct chemical reactivity.
The theory that cells have evolved to utilize electromagnetic fields to catalyze biological processes has been previously proposed. 10 Further theoretical analyses propose that biological systems utilize discrete emission energies of cellular components to translate nonchemical, noncontact information. 11 Experimental evidence suggests that biological structures interact through quantized bursts of photons in finite regions of the electromagnetic spectrum at low-energy density (∼10−20 J), yet these claims have not been broadly studied or well characterized. 12
While there are current limitations in instrument sensitivity that prevent reproducible detection of bioelectronic emissions produced by subcellular components in complex environmental matrices, evidence suggests that bioelectronic emissions at the cellular level occur in finite regions of the electromagnetic spectrum. 13 When cells are exposed to discrete wavelengths within the electromagnetic spectrum, cellular development and differentiation is known to be affected.10,14,15 While it is not known which wavelengths in the electromagnetic spectrum produce discrete cellular changes, such evidence suggests that discrete wavelengths are absorbed and emitted by cells to produce change unrelated to cellular stress. Therefore, biophysical theory of how discrete wavelengths are absorbed and emitted by cellular components must be more well defined to support experimental advances.
As many biomolecules of appreciable mass carry net electric charges (e.g., DNA, RNA, proteins), it is expected that their oscillation within Earth's magnetic field can generate local electromagnetic fields. Long-standing principles of electromagnetism, originally proposed by Lorentz, posit that a magnetic force perpendicular to a charged particle is dependent upon the velocity of that particle. At the nanoscale, charged biomolecules contain a Brownian velocity that oscillate within Earth's magnetic field. While charged biomolecules colliding with molecules that comprise the surrounding media in a cell are expected to be random, more recent advances in Brownian motion of biomolecules suggest that such phenomena may be governed by nonrandom dynamics that are detectable by their relative motions. 16 If such Brownian velocities of biomolecules are treated as nonrandom, then it is plausible that a charged biomolecule moving within a cell in a magnetic field may undergo periodic electromagnetic emissions that are observable.
Understanding how electromagnetic emissions within cells may be a necessary step toward developing a more complete model of bioelectricity. Current theory has yet to conceptualize how charged subcellular components, with a Brownian velocity within Earth's magnetic field, contribute to electromagnetic emissions at discrete wavelengths. As a method to gain more fundamental understanding of electromagnetism in biological systems, this work explores how the Brownian velocity of double-stranded DNA (dsDNA) moving in Earth's magnetic field can generate local electromagnetic emissions. The overarching goal of this work is to fill the theoretical gap of how dsDNA may emit finite wavelengths and further conceptualize how these forces may contribute to discrete biological change on electromagnetic timescales. Practical applications of this theoretical work may enable novel spectroscopic methods to measure multilevel biological phenomena and augment current genomic tools.
Derivation of Average Brownian Velocity for dsDNA
The theory of Brownian movement has been experimentally validated, yet has not been applied to problems involving the generation of electromagnetic radiation by singular components at the subcellular level. 17 Recent measurements have experimentally verified the Maxwell–Boltzmann distribution of velocities for individual Brownian particles and the equipartition theorem of energy for Brownian motion.18,19 For calculating the average Brownian velocity of dsDNA in a cellular environment, it is known that the inertia of the liquid medium surrounding a biomolecule of interest is important. 20 Inertia of the liquid medium inside the cell has been included for calculating the root mean squared instantaneous Brownian velocity of dsDNA and is given by Equation (1):
where
where mp is the individual mass of dsDNA within a cell and mf is the mass of the fluid that comprises that cell and is approximated here as total cell mass. Substitution of Equation (2) into Equation (1) gives the explicit formula in Equation (3) for the root mean squared particle velocity within a cell and is depicted in Figure 1.
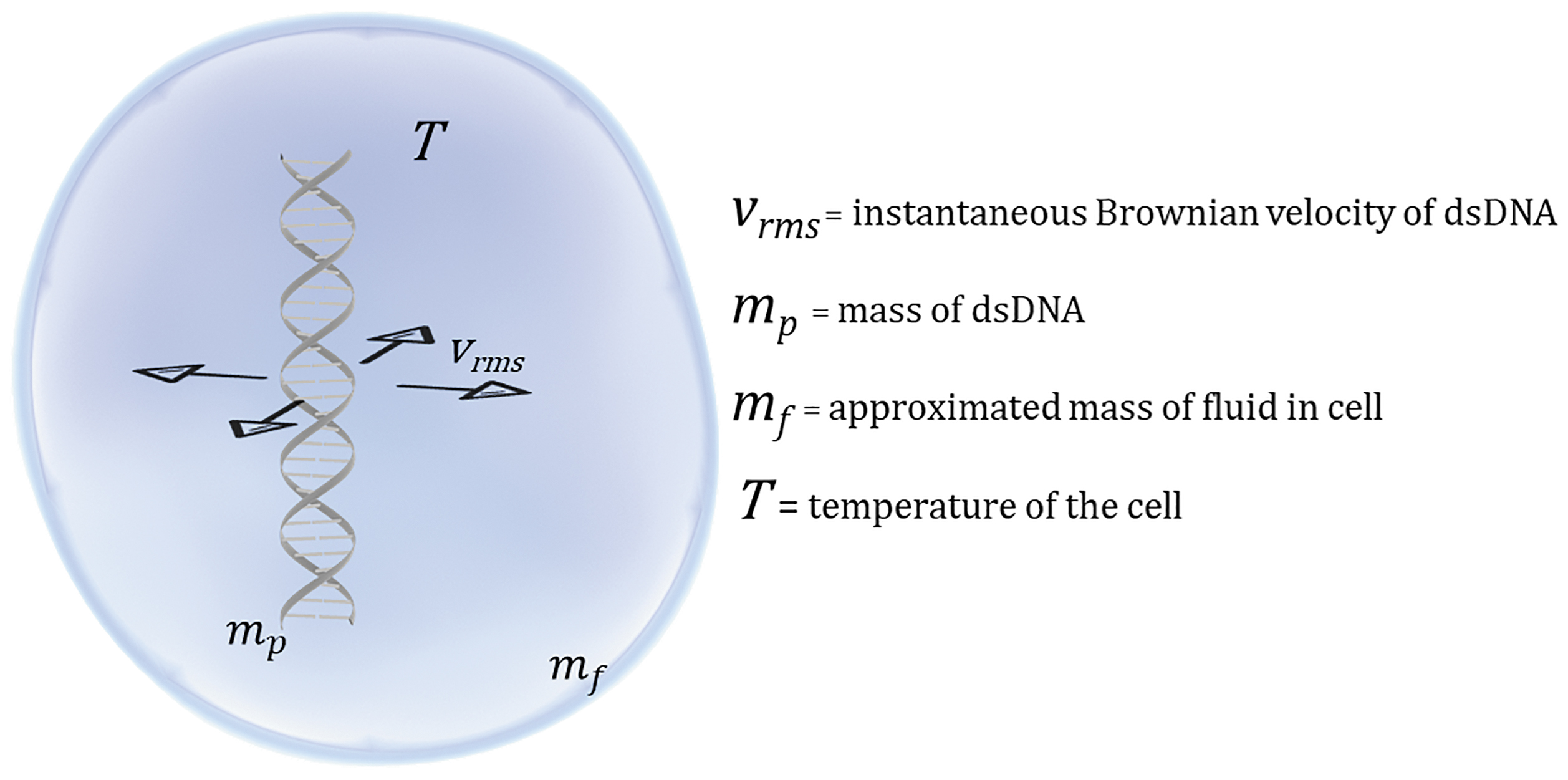
Depiction of variables involved in Equations (2) and (3). dsDNA, double-stranded DNA.
Equation (3) will be used to calculate average instantaneous Brownian velocity of dsDNA, to interrogate individual velocities that contribute to discrete energies associated with electromagnetic emissions from dsDNA within cells.
Calculation of Individual dsDNA and Cell Masses
While this theory is general and may be extended to many types of charged biomolecules within a cell, dsDNA was selected based on the wealth of sequencing data available for many species and the ease of conversion to mass. Both experimentally determined and theoretically derived masses of cells and dsDNA were used and are described here further. Advanced optical imaging measurements have estimated the mass of human red blood cell masses at 27 pg and will be used as an approximate value for human cells across various genome sizes.21,22 Yeast cell mass will be taken to 79 pg, based on previous estimates. 23 Escherichia coli bacterium cell mass will be taken to be 0.67 pg, based on experimental measurements with nanoelectromechanical systems. 24
It is known that cellular density, volume, and mass varies dependent on phase in its life cycle. 25 However, static estimates used in this study were chosen as a proof-of-concept method for more precise measurements in the future. The masses of dsDNA were calculated with known genome sizes in base pairs (bp).26,27 The number of base pairs were then multiplied by the average molecular weight of individual base pairs (660 g/mol) and converted to mass in grams with Avogadro's number (bp/mol). 28
Derivation of dsDNA Emission Wavelengths
As the velocity of various macromolecules within a cell can be deduced through known equations of Brownian motion, then the energy associated with that motion can thus be calculated for a known mass. The energy E associated with the Brownian velocity of a subcellular component is given in Equation (4):
Substitution of the vrms of Equation (3) into Equation (4) gives Equation (5) and yields the energy associated with the mass of a subcellular component in its condensed form [Eq. (6)].
Substitution of Equation (6) into the Planck–Einstein relationship [Eq. (7)] allows for the energy of photons emitted from a given subcellular mass within a cell to be written as a frequency in Equation (8):
where f is the frequency (s−1) and h is Planck's constant (6.626 × 10−34 J-s). For convenience in spectroscopy, the frequency of Brownian velocity of a charged mass in a cell can be converted to an emission wavelength by substitution of Equation (8) into Equation (9), where c is the speed of light, to yield the condensed form of Equation (10).
Electromagnetic waves emitted from a point source to the surrounding medium are not known to change in frequency but are known to change wavelength, which is dependent upon the refractive index of the medium through which they propagate. If an electromagnetic wave is emitted from a charged biomolecule within a cell into the air, then the wave must move from a denser medium in a cell λcell to a less dense medium of air λair. The corresponding change in wavelength measured in the less dense medium will be proportional to the refractive index n and is given by Equation (11).
Values for the refractive indices of cell membranes have been measured between 1.46 and 1.60 (Ref. 29 ). However, a value of 1.46 will be used as an estimate for all cell types in this calculation as a first approximation. Substitution of Equation (10) into Equation (11) gives the predicted wavelength in air that is emitted by charged subcellular components [Eq. (12)]. A simplified schematic is shown in Figure 2.
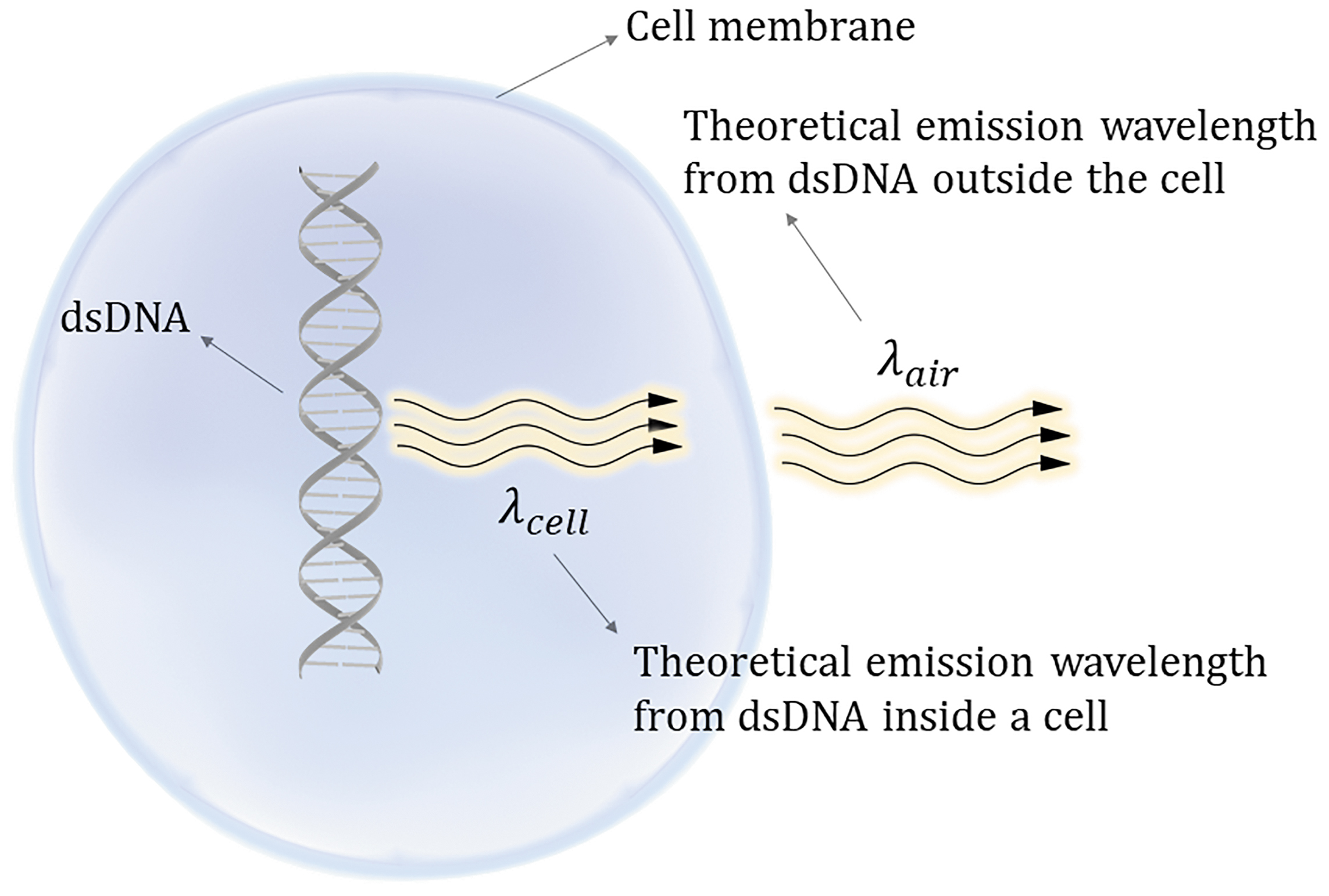
Simplified schematic of emission wavelengths from dsDNA.
Results and Discussion
Emission energies for masses of dsDNA that comprise subcellular components were calculated according to Equation (6) and frequencies were calculated according to Equation (8). The results of calculations are compiled in Table 1, where the frequencies are listed along with their associated region within the electromagnetic spectrum. In accordance with spectroscopic frames of reference, theoretical dsDNA emission wavelengths within a cell and in air were calculated in nanometers using Equations (10) and (12), respectively.
Results of Calculations for Predicted Wavelengths Emitted by Double-Stranded DNA in Various Cellular Systems
Predicted wavelengths were calculated in meters.
EM, electromagnetic.
The human genome emission wavelength outside the cell corresponds well with experimentally observed values obtained from spectral pattern analyses of photon emissions from the human body. Emission spectra measured from human skin covers a range from 450 to 750 nm, with maximum intensities ranging from 570 to 670 nm. 30 Theoretical emission wavelength of the human genome is reported in this study as 660 nm and is well within the measured range of the maximum intensity of bioelectronic photon emission from human skin. Predicted emission wavelength for mitochondrial dsDNA is in the microwave range. While no current measurements have directly observed a specific mitochondrial emission frequency of 4.3 GHz, there is evidence to suggest that exogenous electromagnetic perturbations of mitochondria within the range of microwave frequencies around 4.0 GHz affect their underlying function. 31 A frequency of 1.1 THz for Saccharomyces cerevisiae DNA is predicted and closely corresponds with measurements in the frequency band between 0.75 and 1.1 THz that have been used to monitor changes in S. cerevisiae. 32 Emission wavelengths associated with dsDNA in two strains of E. coli (one benign to humans the other pathogenic) fall within with the infrared range. Laser tweezer Raman spectroscopy has been used to measure single cells of E. coli and spectra correspond to peak intensities from 1000 to 2000 cm−1 (Ref. 33 ) Theoretical predictions using Brownian velocity of dsDNA within Escherichia coli K-12 and Escherichia coli O157:H7 are well within this range, corresponding to 1126 and 1300 cm−1, respectively.
Several factors may influence predicted outcomes of dsDNA emission wavelengths by this method. Mass differences from average molecular weights of dsDNA base pairs may result in variation of predicted subcellular emission frequencies. If an exact DNA sequence of interest is known, then more accurate molecular weights can be achieved to greatly increase mass accuracy in these calculations. More refined experimental methods, such as using high-fidelity DNA sequencing information, can be applied to obtain exact masses in systems of interest. Multiple biophysical factors may affect the predicted refractive index of endogenously produced dsDNA emission wavelengths, and therefore may alter the observed emission wavelengths measured outside the cell [Eq. (12)]. Two such factors are the molecular composition and mobility found within cellular membranes. Previous calculations have shown that the lateral diffusion of phospholipids and proteins in cell membranes is around 2 μm/s. 34 As phospholipid structure across cell membranes is not static, the refractive index must be more accurately measured or normalized based on the species of observation.
The agreement of spectroscopic observations in the literature with predicted bioelectromagnetic emission wavelengths of dsDNA is encouraging, for a first approximation of applying this Planck–Einstein–de Broglie relationship in a biological frame of reference. However, this approximation is based on a singular moment in time when dsDNA mass is static. The mass of dsDNA is a dynamic process where changes in mass are reflective of the biological needs of the cell at any given time. There are numerous examples of dsDNA processes that alter the overall mass of dsDNA that subsequently impact overall cell dynamics. Transposition elements promote DNA double-strand cleavage and cause temporary changes in overall dsDNA mass through insertions, deletions, and translocations. Relevant to epigenetics, methylation of cytosine adds mass to dsDNA (15 g/mol per methylation per base pair). Relevant to dsDNA damage/repair processes, oxidation of nucleic acids also adds mass (16 g/mol per oxidation per base pair). Relatively small contributions to the overall dsDNA mass that arise from mass additions to individual base pairs (e.g., oxidation, methylation) and/or changes in genome size (e.g., transposable element insertions/deletions) may seem negligible. However, when taken in aggregate across an entire genome, this may lead to substantial changes in mass differences between discrete genetic states. This theory implies that such mass differences in dsDNA can result in a unique corresponding emission wavelength that may be reflective of a discrete genetic state on electromagnetic timescales.
Another implication of this theory relates to the ubiquitous presence of noncoding regions of dsDNA, the so-called “dark matter” of the genome that vastly outnumbers coding regions of DNA in many organisms. 35 In genetics, noncoding regions are often viewed as inactive and this inactivity is defined by the lack of detectable genetic products (e.g., messenger RNA or proteins) that are produced through known transcription/translation processes within a cell. While the structure/function relationships of noncoding regions of dsDNA are not completely understood, noncoding regions show exceptionally high levels of similarity across species and are highly conserved genetic elements in biological systems. 36 As noncoding regions dsDNA represent the majority of the mass of a genome, these highly conserved masses of dsDNA may contribute to the bioelectromagnetic emissions from dsDNA and may be critical to maintaining a specific electromagnetic microenvironment within a cell. Noncoding DNA is thought to play a significant role in gene regulation and experimental data support that gene regulation may occur through specific electromagnetic response elements. 37 While genomic sequencing has identified patterns of noncoding regions of dsDNA, specific genomic masses and their corresponding energetic signatures have not been previously considered as a method to quantify multilevel bioelectromagnetic phenomena associated with noncoding dsDNA.
Conclusion
This theoretical work leverages the well-substantiated Planck–Einstein–de Broglie relationship and applies this concept to Brownian velocity within a cell to describe the relationship between masses of dsDNA, the average Brownian velocity of dsDNA movement within a cell, and the theoretical wavelengths at which dsDNA may emit bioelectromagnetic waves. The utility of unifying concepts from theories of Brownian motion in a biological context with electromagnetism is to yield information on how changes in dsDNA mass relate to a particular genetic state and can be measured by new spectroscopic methods. Further development in measuring such bioelectronic states may become important quantitative tools for biology, where measuring bioelectronic states through nondestructive spectroscopic methods may increase the accuracy of tests that monitor cellular processes. New spectroscopic interpretations of bioelectronic states are also needed to provide for orthogonal methods of biological measurement without using invasive and/or destructive biochemical techniques. While this theoretical study focuses on dsDNA, this methodology may be generalizable to charged biomolecules of an appreciable mass and may be extended to other subcellular components that affect cell dynamics (e.g., RNA or proteins). With improvements in instrumentation to measure theoretical bioelectromagnetic emissions, refinements to this theory may lead to new spectroscopic tools that can quantify multilevel phenomena associated with dsDNA in cells on electromagnetic timescales.
Footnotes
Acknowledgments
The author would like to thank Heather Meeks of the Defense Threat Reduction Agency's Research and Development Directorate for the fascinating intellectual discussions surrounding progressive ideas in radiobiology, which led to the generation of this work.
Author Contributions
The author is the sole contributor to this work and is responsible for this article. Furthermore, the author confirms this material or similar material has not been and will not be submitted to or published in any other publication before its appearance in Bioelectricity.
Author Disclosure Statements
There are no conflicts of interest to report related to this work and no competing financial interests exist.
Funding Information
No funding was received for this article.