
Abstract
Background:
Voltage-gated sodium channels (VGSCs) are upregulated during cancer progression and promote metastatic cell behaviors. In contrast, voltage-gated potassium channels (VGPCs) that control proliferation, among other roles, are downregulated. In this study, we tested the effects of blocking VGPCs on the Matrigel invasiveness specifically of strongly metastatic cancer cells.
Materials and Methods:
The rat prostate cancer Mat-LyLu cell line and the human breast cancer MDA-MB-231 cell line, both strongly metastatic and expressing functional VGSCs, were adopted as models. Matrigel invasion assays and patch-clamp electrophysiology were used to monitor pharmacological effects of VGPC blockage.
Results:
Margatoxin (MGTX) significantly suppressed the outward currents in Mat-LyLu cells, consistent with Kv1.3 being the predominant VGPC in these cells. For the MDA-MB-231 cells, 4-aminopyridine (4-AP) was used as a general blocker of VGPCs. Treatment with the respective blocker significantly increased the cellular invasiveness under conditions wherein proliferative activity was not affected. As expected, invasiveness was reduced by treating the cells with the VGSC blocker tetrodotoxin (TTX). Importantly, the proinvasive effect of MGTX was suppressed completely by cotreating the MDA-MB-231 cells with TTX.
Conclusions:
(1) VGPC blockage promotes invasiveness in strongly metastatic carcinoma cell lines and this is associated with functional VGSC expression well known to occur in such cells. (2) The available data taken together are supportive of the Celex Hypothesis, proposing that the electrical excitability of cancer cells (resulting from functional VGSC upregulation and concomitant VGPC downregulation) promotes metastatic invasiveness.
Introduction
A variety of ion channels are known to contribute to different components of the cancer process, from initiation to metastasis. 1 In particular, voltage-gated sodium channels (VGSCs) promote metastatic cell behaviors such as motility and invasiveness in vitro and metastasis in vivo.2–4 In contrast, voltage-gated potassium channels (VGPCs) are pivotal for cell proliferation, among other roles. 5 This is due, in part, to the role of VGPCs in generating and modulating the membrane potential (Vm). 6 The role of VGPCs in cancer cell behavior appears complex, as well as proliferation, effects on adhesion, apoptosis, invasion, angiogenesis, etc. have also been attributed to VGPC expression/activity in vitro and/or in vivo. 7 There are at least two main reasons for this complexity. First, conceptual, changes in Vm can have profound secondary effects on a range of cellular activities. Second, technical, in key in vitro functional assays, such as the Boyden chamber, proliferation is integral to Matrigel invasion and this can complicate interpretation of the data.
Electrophysiological (whole-cell patch-clamp) recordings of voltage-activated membrane currents in cell lines from a variety of carcinomas revealed that compared with nonmetastatic cells, metastatic cancer cells show (1) functional VGSCs and (2) much reduced outward currents due mainly to VGPC activity.3,4,8,9 These observations led to the development of the “Celex Hypothesis,” which proposes (1) that the combination of functional VGSCs coupled with reduced VGPC activity would make the membranes of metastatic cells electrically excitable and (2) that the excitability, in turn, would make the cells hyperactive and enable them to behave aggressively, with disruption/invasion of their surroundings, as would be expected to occur during metastasis. 10 Indeed, the strongly metastatic human breast cancer MDA-MB-231 cells were shown recently to spontaneously generate VGSC-dependent action potentials (APs). 11
In this study we have investigated the effect of VGPC blockage on invasiveness of metastatic cell lines from rat prostate cancer and human breast cancer. First, for the former, we used the Mat-LyLu cells that are strongly metastatic and express a functional VGSC (Nav1.7).4,12 Blocking VGSC activity using tetrodotoxin (TTX) or ranolazine suppressed their invasiveness in vitro and metastatic ability in vivo.4,13,14 The MDA-MB-231 cell line, a classic model of triple-negative breast cancer, also expresses a functional VGSC, the neonatal splice variant of Nav1.5 (nNav1.5). 3 Silencing nNav1.5 (using siRNA), or treating the cells with TTX or ranolazine, suppressed their invasiveness in vitro and metastatic ability in vivo.3,15–17 Both cell lines have significantly reduced VGPC currents in comparison with their weakly/nonmetastatic counterparts.3,8
Our specific aim here was to study the consequences of blocking VGPC activity in the Mat-LyLu and MDA-MB-231 cell lines on their Matrigel invasiveness as an in vitro indicator of their metastasis capability. Importantly, the experiments were carried out under conditions wherein the cells' proliferative activities were not affected.
Materials and Methods
Cell culture and chemicals
Rat prostate cancer Mat-LyLu cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 1% fetal calf serum, 2 mM
Electrophysiology
Whole-cell patch-clamp recordings were performed on cells under superfusion with mammalian physiological saline containing (in mM) 144 NaCl, 5.4 KCl, 1 MgCl2, 2.5 CaCl2, 5 HEPES, and 5.6
Cell viability, proliferation, and Matrigel invasion
These assays were described previously in detail for both cell lines: Mat-LyLu 4 and MDA-MB-231.3,15 In brief, cell viability was determined by the trypan blue dye exclusion assay. Upon completion of the treatment period (48 h), the medium was aspirated and replaced for 10 min with 0.2 mL of 0.4% trypan blue (Sigma-Aldrich) and 0.8 mL medium. The cultures were returned to 1 mL fresh medium and the cells (stained and nonstained) were counted under an inverted microscope (ID 03, Zeiss) at 200 × magnification. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to assess number of cells (which was assumed to represent proliferative activity in the absence of toxicity). Cells (2 × 104 cells/well) were seeded in 24-well plates (Becton Dickinson, Oxford, UK). Treatment lasted 48 h with cells incubated in 500 μL of the drug solution(s) in each well. The drug solutions were then removed and 400 μL medium and 100 μL MTT solution were added into each well. The plates were incubated for 3 h at 37°C. The bathing MTT solution was aspirated and replaced with 500 μL dimethyl sulfoxide and 75 μL of Sorenson's glycine buffer. The plates were rocked at 150 cycles/min for 5 min and the absorbance was measured at 570 nm on a multiplate reader (ELX800 Universal Microplate Reader, Bio-Tek Instruments, UK). Absorbance was converted to number of cells using a linear standard curve. Cellular invasiveness was measured using modified Boyden chambers, again as described before.3,4,15 Chemotactic gradients of 0.1%–1% and 1%–5% FBS were used to facilitate movement, and assays lasted 48 and 12 h for Mat-LyLu and MDA-MB-231 cells, respectively.3,4,15 Twelve randomly chosen fields of view were counted for each insert. Seven inserts were used for the MGTX (and control) experiments; 10 inserts for 4-AP and control; 3 inserts for TTX experiments on Mat-LyLu cells; and 8 inserts for TTX and 4-AP+TTX experiments. The number of cells in each field of view was then normalized to the largest cell count in the respective control inserts.
Data analysis
Parametric data were analyzed as means ± standard errors of the mean, whereas nonparametric data are presented as median, interquartile range, 5% and 95% confidence intervals, and outliers (GraphPad Prism 5; GraphPad Sowftware, Inc., San Diego, CA). Parametric data were statistically analyzed with Student's t-test whereas nonparametric data were analyzed by Mann–Whitney U-test. (***) indicates p < 0.001.
Results
We have shown previously that the predominantly expressed VGPC in Mat-LyLu cells is Kv1.3, of which MGTX is a selective blocker. 8 First, we performed two control experiments. As expected, acute application of 10 nM MGTX to the Mat-LyLu cells resulted in significant reduction in the outward current by 43.5% ± 7.5% (p < 0.001; n = 4; Fig. 1A). Also, as expected, 1 μM TTX significantly decreased the number of invading cells by 37% ± 3% (p < 0.001; Fig. 2). The 10 nM MGTX did not affect the cell viability or proliferation for 48 h (not shown). Importantly, however, treatment with 10 nM MGTX significantly increased the Matrigel invasiveness by 57% ± 4% (p < 0.001; Fig. 2). We also questioned whether long-term (24 h) treatment with MGTX would affect the VGSC characteristics. Compared with control recordings, there was no effect on the VGSC peak current density (15.9 ± 2.9 vs. 14.5 ± 2.1 pA/pF; n = 10 and 12, respectively), the proportion of cells with VGSC, the current–voltage relationship (Fig. 1B), steady-state inactivation (Fig. 1C), or recovery from inactivation (Fig. 1D). It was concluded that the steady-state VGSC expression did not change after long-term treatment with MGTX.

Electrophysiology and pharmacology of VGPC (Kv1.3) and VGSC expression in Mat-LyLu cells.
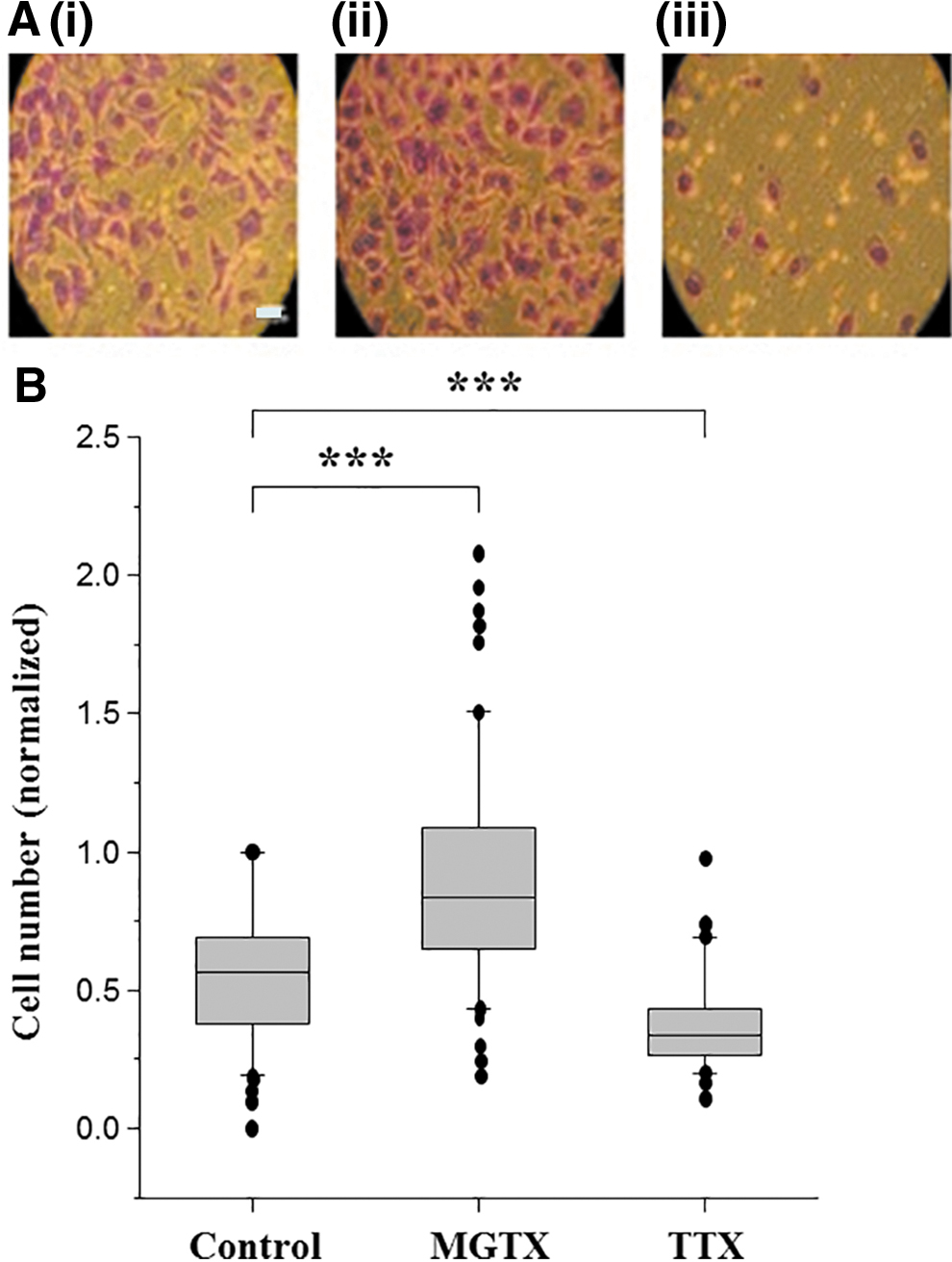
Effects of MGTX and TTX on Matrigel invasiveness of Mat-LyLu cells.
We then studied the effect of VGPC block on invasion for the MDA-MB-231 cell line (Fig. 3). Since the molecular identity of the predominant VGPC(s) in the MDA-MB-231 cells is still uncertain, we used 4-AP as a general blocker of VGPCs. At 1.6 mM, 4-AP had no effect on cell viability or proliferation for 24 h (data not shown). Matrigel invasiveness was studied for 12 h under four conditions in parallel: 4-AP, TTX, 4-AP+TTX, and compared with the control. Treatment with 4-AP significantly increased the invasiveness by 39% ± 6% for 12 h (p < 0.001; Fig. 3). As expected, 10 μM TTX significantly reduced the invasiveness of the MDA-MB-231 cells by 56% ± 4% (Fig. 3; p < 0.001). Importantly, combined application of 1.6 mM 4-AP and 10 μM TTX reduced invasion to the same extent as when TTX was applied alone, that is, with the VGSC activity blocked, the proinvasive effect of 4-AP was completely suppressed (Fig. 3).
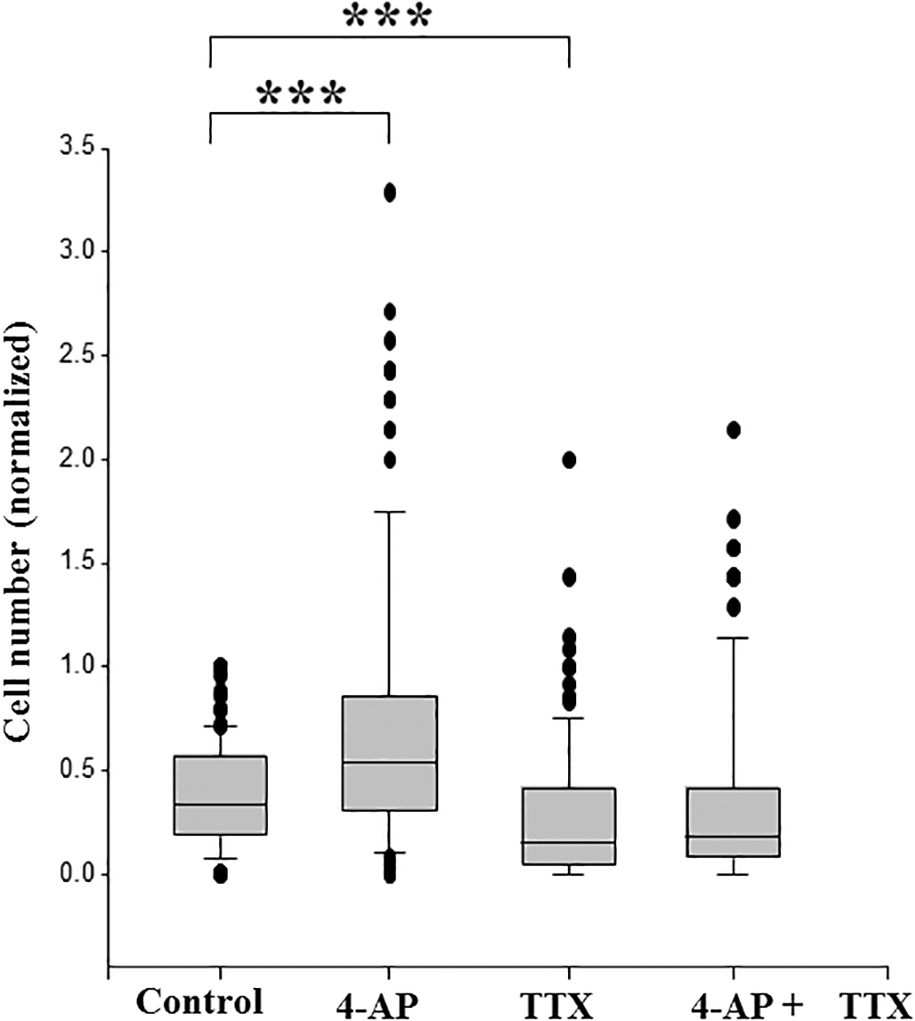
Effects of MGTX and TTX on Matrigel invasiveness of MDA-MB-231 cells. Cells were treated with 4-AP (1.6 mM), TTX (10 μM), or 4-AP+TTX, and compared with untreated controls. Data are presented as box plots showing medians, interquartile range, 25% and 75% confidence intervals, and outliers. Twelve fields of view were measured for each of 10 inserts (for control and 4-AP) or 8 inserts (for TTX, and 4-AP+TTX). ***p < 0.001. 4-AP, 4-aminopyridine.
Discussion
Prostate and breast cancer share several similarities, including hormone sensitivity, metastasis to bone, and expression of VGSC (although different subtypes), and tend to co-occur in families.3,4,23,24 Our main conclusion here is that suppressing VGPC activity in both carcinoma cell lines of strong metastatic potential promoted their Matrigel invasiveness significantly by 40%–50%. Also, as shown many times already, invasiveness was suppressed by TTX.3,4 These effects, which occurred under conditions wherein cells' proliferative activities were not affected, are consistent with the Celex Hypothesis. 10 Furthermore, the complete suppression by TTX of the proinvasive effect of VGPC blockage in the MDA-MB-231 cells would suggest that VGSC activity played a significant intermediary role in the effect.
In this study, a concentration of MGTX (10 nM) was chosen that blocked a significant portion of the outward current in Mat-LyLu cells without affecting their proliferation. The latter is inherent to the Matrigel invasion assays lasting for 24 h or longer. Although MGTX-sensitive Kv1.3 is the predominant VGPC in Mat-LyLu cells, low levels of Kv1.4 and Kv1.6 are also expressed. 8 However, it is not known whether these channels also contribute to the cells' invasiveness (and/or proliferation). As regards the MDA-MB-231 cells, it is known from electrophysiological data that these cells possess only low-level functional VGPCs, with outward currents of ∼100 pA (at +60 mV). 3 Nevertheless, MDA-MB-231 cells have been reported to express a range of VGPCs, including Kv1.1, 1.3, 25 Kv10.1, 26 and Kv2.1, 3.2, 4.2, 12.2. 27 However, since their relative contributions (if any) are not known, we chose to use 4-AP as a general blocker of VGPCs. Again, a concentration (1.6 mM) that did not affect proliferation was used (although it was unlikely that this would change over the 12 h invasion time for the MDA-MB-231 cells). Importantly, blocking the VGPC activities overall with 4-AP resulted in increased invasiveness, and this effect was blocked completely by cotreatment with TTX. Interestingly, somewhat different results were reported earlier. 25 It was found, also using MDA-MB-231 cells, that treatment with “Kv1 blocker” toxins from scorpion venom suppressed “migration.” The latter was a different assay to ours, however, since it involved (1) plating of the cells on small quantities of fibronectin (rather than Matrigel), just to promote adhesion, and, correspondingly, (2) the assay was run for 5 h (cf. 24 h in our experiments). It has also been observed that the inhibitory effect of TTX in Boyden chamber assays (incorporating 8 μm transwell filters) is seen only when the filters are coated with Matrigel (SP Fraser, unpublished results). 28 It would appear, therefore, the VGSC–VGPC interaction as proposed by the Celex Hypothesis is strictly a phenomenon of invasiveness, which is a major component of metastasis. Consistent with this notion, first, the VGPC-induced increase in invasiveness is seen in functional assays involving degradation of Matrigel, which, in turn, involves proteolysis driven by VGSC-dependent acidification of pericellular pH. 29 Second, VGPC inhibitors were much less effective in suppressing the proliferation of the strongly metastatic VGSC-expressing Mat-LyLu cells compared with the isogenic but weakly/nonmetastatic AT-2 cells. 30 Finally, also in support of the Celex Hypothesis, it has been shown conversely that promoting Kv11.1 activity pharmacologically in MDA-MB-231 cells in vivo suppressed metastasis, in part, by reprogramming epithelial–mesenchymal transition (EMT). 31 The latter is an early event in invasion and largely responsible for conferring malignancy-associated phenotypes. 32 Importantly, EMT is controlled by VGSC activity. 33
Finally, we can place the results in the context of the Celex Hypothesis. Central to the argument is the plasma membrane potential (Vm).34,35 Basically, blocking the “background” VGPC(s) would be expected to depolarize the Vm that will promote “membrane excitability” (VGSC activity) including generation of TTX-sensitive APs. The latter were recently demonstrated independently to occur under resting conditions in the MDA-MB-231 cells. 11 Accordingly, enhancing invasiveness by MGTX or 4-AP will be opposed by the VGSC blocker, TTX, as observed. In addition, but somewhat less importantly, VGPC blockage will prolong the APs that could also promote invasiveness by allowing more Na+ to enter the cells.
In conclusion, the evidence presented here supports the Celex Hypothesis. Overall, the functional role of VGPC expression in the cancer process appears to be complex. In particular, the role would depend on the stage of the cancer and, hence, the metastatic character of the cell under investigation.
Footnotes
Author Disclosure Statement
M.B.A.D. is involved in a small biotech company aiming to develop ion channel blockers as cancer drugs.
Funding Information
A.T. and B.B. were supported by the Erasmus Program of the European Union.