
Abstract
Our current knowledge about the Na+/K+ pump is mainly of the structure and function of the pump itself. The knowledge of the relationship between the function of the pump and other cellular functions is inadequate. This is because we are unaware of what the pumping process is and why the pump takes 3 Na+ to 2 K+ as the pumping ratio. In recent years, I proposed a dipole-charging model to simulate the pumping process of the Na+/K+ pump and showed by this model that this pumping ratio of the Na+/K+ pump is determined by the ratio of the ionic mobilities of K+ to Na+ ions. In this article, I further propose an osmotic balance condition that the potassium ion concentration inside the cell and the sodium ion concentration outside the cell are equal. With this condition, the dipole-charging model is further extended to compare different pumping ratios and it shows that only the pumping ratio 3 Na+ to 2 K+ can be selected during evolution. This result also reveals the relationships between the 3 Na+ to 2 K+ pumping ratio, the membrane resting potential, and the osmotic balance condition of the cell. Furthermore, I discuss the osmotic balance condition, the stability of the dipole-charging model and the dipole-charging process, the electrogenic contribution by the Na+/K+ pump to the resting membrane potential, and the effectiveness and the prospect of the dipole-charging model. The dipole-charging model is capable of explaining all the phenomena related to the pumping process of the Na+/K+ pump and is especially suitable for study in situations under pathologic conditions, such as ischemia or hypoxia. Finally, I present analyses of the free energy change of the coupled overall reaction for the Na+/K+ pump and of some reports on different pumping ratios of the Na+/K+ pump. The exception of the pumping ratio in the brine shrimp Artemia is also discussed.
Introduction
There are many ion transporters in animal cells, such as the Na+/K+ pump, the sarco(endo)plasmic reticulum Ca2+ pump, and the plasma membrane Ca2+ pump, which are primary active transporters; and the Na+–glucose cotransporter and the Na+/Ca2+ exchanger as a countertransporter, which are secondary active transporters. 1 All these transporters take part in different physiological functions. 2 For example, for each adenosine triphosphate (ATP) hydrolyzed in one pumping cycle, the Na+/K+ pump transports 3 Na+ ions out of the cell in exchange for 2 K+ ions transported into the cell and maintains large-concentration gradients of Na+ and K+ ions. 3 These concentration gradients are critical for maintaining resting membrane potential, the secondary active transport, and the osmotic balance of the cell. 4
However, unlike other transporters, the Na+/K+ pump does not work at the equilibrium state. 5 That is, the ATP provides more energy than required for pumping 3 Na+ and 2 K+ ions. So the pumping ratio of the Na+/K+ pump remains a question. 6
In recent years, I proposed a dipole-charging model to simulate the pumping process of the Na+/K+ pump and showed that this pumping ratio of the Na+/K+ pump is determined by the ratio of the ionic mobilities of K+ to Na+ ions. 6 In this model, the pumping of the Na+/K+ pump is considered to be a dipole-charging process, in which the pumping of 3 Na+ ions out and 2 K+ ions in creates two electric dipoles, which subsequently charge the cell membrane. We know that, for neurons, the resting potential is −70 mV and the cell membrane has an electric capacitance of 1 μF/cm2. The amount of charge required for producing this membrane potential is about 4350 monovalent ions/μm2. 6 This means that 4350 monovalent ions/μm2 corresponds to −70 mV, provided that the membrane capacitance is 1 μF/cm2, regardless of cell type. However, the pumping rate of the Na+/K+ pump can be 100 times per second, 7 and the density of the Na+/K+ pump can be several thousand pumps/μm2 on the membrane fragments isolated from kidney outer medulla.8–10 So any accumulation of charges on the membrane caused by charging of the two dipoles at this pumping rate and this pump density could significantly change the membrane potential if those charges were not discharged instantly and in time. For this reason, the pumping process of the Na+/K+ pump should be precisely regulated, including the intracellular and the extracellular concentrations of sodium and potassium ions, because these concentrations contribute to the electrolytic conductivities of the intracellular and the extracellular solutions. This is why we should develop a dipole-charging model to simulate the pumping process of the Na+/K+ pump.
By this dipole-charging model, we already showed that when the pumping ratio is taken as 3 Na+ to 2 K+, the concentration ratios CK,i/CNa,o ≈ 1.0 and CK,o/CNa,i ≈ 0.30 can be obtained, 6 where CNa,i and CK,i are the sodium and potassium ion concentrations inside the cell, and CNa,o and CK,o are the sodium and potassium ion concentrations outside the cell. The observation values of CK,i/CNa,o and CK,o/CNa,i for different cell types are shown to be consistent with the two predicted values (see Table 1 in Ref. 6 ).
Calculated CK,o/CNa,i and CK,i/CNa,o According to the Pumping Ratio nna/nk
nna, nk are assumed to take 1 to 3.
In this study, we take a further view of this question. First, we assume that the pumping ratio of the Na+/K+ pump can take different values and the corresponding concentration ratios are calculated according to the dipole-charging model. Then a suitable concentration ratio is selected according to the osmotic balance condition of the cell. With this concentration ratio, the pumping ratio of 3 Na+ to 2 K+ can be uniquely determined by the ratio of the ionic mobilities of K+ to Na+ ions. Finally, I discuss the fact that the pumping ratio of the Na+/K+ pump is not due to the energy requirement, and I also discuss some reports on different pumping ratios of the Na+/K+ pump.
Methods
The dipole-charging model
Detailed analysis of the dipole-charging model can be found in Ref. 6 In this study, I outline the main features of the model.
The pumping process of the Na+/K+ pump involves two parts. In the first or the sodium part, 3 Na+ ions inside the cell are transported to the outside while leaving three negative charges inside. This will create an electric dipole that has charge +3e outside the cell membrane and charge −3e inside the cell membrane, where e is the elementary charge. This electric dipole will subsequently charge the cell membrane, which has a certain capacitance Cm. In the second or the potassium part, 2 K+ ions outside the cell are transported to the inside while leaving two negative charges outside. This will create an electric dipole that has charge +2e inside the cell membrane and charge −2e outside the cell membrane. This electric dipole will subsequently charge the cell membrane with the same capacitance Cm.
The dipole charging process can be simulated by an electric circuit, which I call the dipole-charging model. A group of equations that describe the circuit are as follows (see Figure 1):
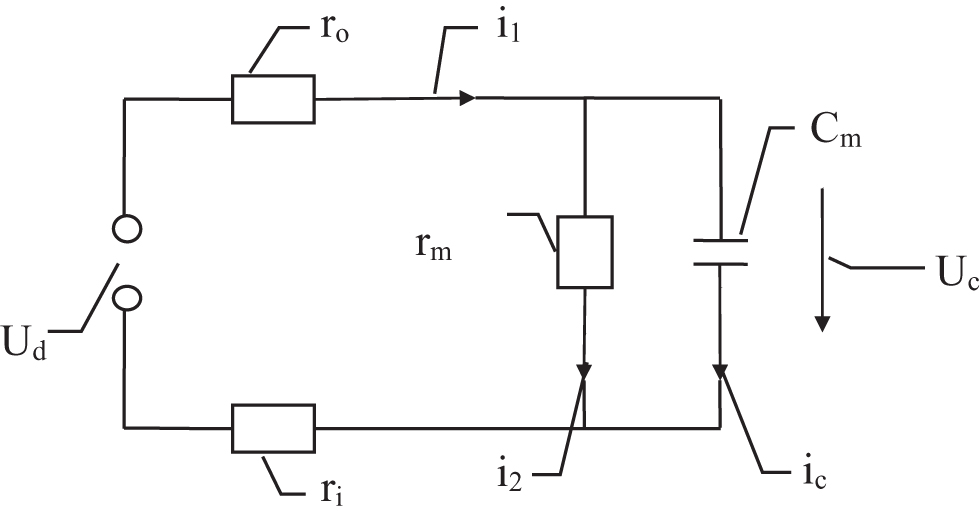
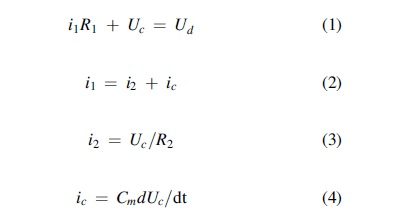
where Ud is the electric potential difference produced by the pump dipole alone and has the unit of V;
Cm is the electric capacitance per unit area of the cell membrane and has the units of F/m2;
Uc is the electric potential difference across the capacitance Cm and has the unit of V;
R1 = ri + ro, where ri is the electric resistance from the inner end of the Na+/K+ pump to the inner surface of the cell membrane and has the unit of Ω; ro is the electric resistance from the outer end of the Na+/K+ pump to the outer surface of the cell membrane and has the unit of Ω. ro and ri are a measure of the resistances of the extracellular and intracellular fluids, respectively;
R2 = rm is the electric resistance of the cell membrane and has the unit of Ω. rm is a measure of the resistance by ion channels of the cell membrane;
i1 is the current through ri and ro and has the unit of A;
i2 is the current through rm and has the unit of A;
ic is the current through the capacitance Cm and has the unit of A.
Solving this group of equations, we have the following:
6
where R// = R1R2/(R1 + R2), τ = R//Cm is the capacitive time constant of the circuit.
The time–average value Ic of ic over 3τ is as follows:
Ic or ic is the part of the charging current flow that directly charges the membrane capacitor.
The membrane charging by the pump dipole is asymmetric,
6
that is, in the sodium part, the electric resistance ri can be taken as zero, so R1 = ro; in the potassium part, the electric resistance ro would be zero, and R1 = ri. According to Equation (6),
So in the sodium part,
In the potassium part,
where Ic,1 is the first or the sodium part of the charging current flow that directly charges the membrane, and Ic,2 is the second or the potassium part of the charging current flow that directly charges the membrane.
The other difference between the two parts is that the potential difference Ud is proportional to the transported charge, or the dipole charge. We assume that the pumping ratio is nna Na+ to nk K+, meaning that in each pumping cycle, nna Na+ ions are pumped out of the cell and nk K+ ions are pumped into the cell, respectively. Then, in the sodium part, Ud = nna ψ, and in the potassium part, Ud = nk ψ, where ψ is defined as the proportional coefficient.
So, in the sodium part,
In the potassium part,
The main contributions to ri and ro are made by the sodium and potassium ions,
6
so we have the following:
where rNa,o and rK,o are the sodium and potassium resistances outside the cell, respectively; and rNa,i and rK,i are the sodium and potassium resistances inside the cell, respectively.
In case of the Na+/K+ pump, the ion flow is produced by a single ion. The corresponding resistance can be calculated as follows:
6
where κ s is the electrolytic conductivity of the solution.
Combining Equations (8), (8′), (9), (9′), and (10), we have
where κNa,o and κK,o are the electrolytic conductivities contributed by the sodium and potassium ions in the extracellular fluid, respectively; and κNa,i and κK,i are the electrolytic conductivities contributed by the sodium and potassium ions in the intracellular fluid, respectively.
The electrolytic conductivity of a solution contributed by a kind of ion is as follows:
11
where κ(i) is the electrolytic conductivity contributed by the ion i in the solution and has the units of S/m;
U(i) is the ionic mobility of the ion i and has the units of m2/(s·V);
C(i) is the concentration of the ion i and has the units of mol/m3;
F = 96,500 J/(V·mol) is the Faraday's constant;
Z(i) is the valence charge of the ion i.
So, according to Equation (12),
where UNa and UK are the ionic mobilities of sodium and potassium ions, respectively, and ZNa = ZK = 1. Substituting Equation (12′) into (11), (11′), we have
We know that Ic, Ic,1, and Ic,2 all indicate the part of the charging current flow that goes on to the membrane. The sodium component of Ic,1 and the potassium component of Ic,2 will affect the membrane electric double layers for the resting potential, so they should be equal to each other, that is,
The potassium component of Ic,1 and the sodium component of Ic,2 can be used by the Na+/K+ pump, so they should be compatible with the pumping ratio, that is,
Equations (14) and (15) relate the pumping ratio, the ionic mobilities of Na+ and K+ ions, and the intracellular and the extracellular concentrations of sodium and potassium ions. By using these two equations, we can calculate CK,o/CNa,i and CK,i/CNa,o according to different pumping ratios of nna/nk.
Results
Table 1 shows the calculated CK,o/CNa,i and CK,i/CNa,o according to the pumping ratio nna/nk, where nna and nk are assumed to take 1 to 3.
From Table 1 we can see that when nna/nk = 3/2, CK,i/CNa,o ≈ 1. We know that sodium ions are the most abundant extracellular cations and potassium ions are the most abundant intracellular cations, and they are the major sources of the extracellular and the intracellular osmotic pressures, respectively 12 (also see Table 2 in Discussion section). So the condition
Osmolar Substances in Extracellular and Intracellular Fluids
Adapted partly from Table 33_2 in Ref. 14
can actually ensure that the cell would maintain osmotic pressure balance. I call this relationship osmotic balance condition. This condition can be used for selection of nna/nk from Equation (14). Substituting Equation (16) into Equation (14), together with UK = 7.62 × 10−8 m2/(s·V) and UNa = 5.19 × 10−8 m2/(s·V), we have
Equation (17) shows that the pumping ratio of 3 Na+ to 2 K+ is determined by the ratio of the ionic mobilities of K+ to Na+ ions. This relation results from the dipole-charging model together with the osmotic balance condition.
Discussion
The dipole-charging model and the osmotic balance condition
Table 1 is calculated from Equations (14) and (15). We can see from Table 1 that CK,i/CNa,o = 1 only when nna/nk = 3/2. We know that the body fluid contains cations and anions (or negatively charged molecules), and they each contribute to about one half of the total osmolarity, respectively (Table 2). As stated above, sodium ions outside the cell and potassium ions inside the cell are the most abundant cations. From Table 2 we can see that, for interstitial fluid, sodium ions contribute to 137/302.2 ≈ 45.33% of the total osmolarity; for intracellular fluid, potassium ions contribute to 141/302.2 ≈ 46.66% of the total osmolarity. The interstitial fluid also contains a large number of chloride ions, and the intracellular fluid contains a high concentration of negatively charged molecules,13,14 but all their contributions to osmolarity belong to the anion (or negatively charged molecules) part (Table 2). So the concentration equivalence CK,i/CNa,o = 1 is enough to ensure the osmotic pressure balance between the interstitial and the intracellular fluids. In this way, only the pumping ratio nna/nk = 3/2 has been selected during evolution.
Stability of the dipole-charging model
For other pumping ratios, the corresponding values of CK,i/CNa,o in Table 1 are far away from 1, so they cannot ensure the osmotic balance condition (16). This is attributed to the stability of the dipole-charging model. We can see that Equations (5) and (6) as well as Equations (14) and (15) are outputs of the group of Equations (1) to (4). We consider Equation (14) here, the inputs of which are nna, nk and CK,i, CNa,o. We can see that Equation (14) is more sensitive to nna and nk than to CK,i and CNa,o. This is because, for each step, nna or nk changes discretely, while CK,i or CNa,o changes continuously. Therefore, the left-hand side of Equation (14) changes significantly with change of nk or nna, but the right-hand side of Equation (14) changes gradually with change of CK,i or CNa,o. This results in two consequences. First, nna/nk can only be 3/2. If nna/nk takes other values, according to Equation (14), CK,i/CNa,o will be far away from 1 and cannot satisfy the osmotic balance condition (16). Second, in the case of nna/nk = 3/2, Equation (14) can allow small variations of CK,i and CNa,o without changing nna/nk to other ratios. These variations are necessary for different cells or animals where CK,i and CNa,o can be adjusted to a small extent in accordance with different concentration combinations of other osmolar substances (Table 2). So here we can see the robustness of the dipole-charging model, which ensures both the pumping ratio nna/nk = 3/2 and the osmotic balance of the cell.
Stability of the dipole-charging process and mechanism for the electrogenic contribution of the Na+/K+ pump
We know that, at rest, there is a higher concentration of sodium ions outside the cell than inside and a higher concentration of potassium ions inside the cell than outside, and there is an electric potential difference ϕ across the cell membrane being negative inside and positive outside. This electric potential difference ϕ is called the membrane resting potential. Therefore, sodium ions tend to enter the cell, which are driven by their concentration gradient and ϕ; potassium ions tend to leave the cell, which are driven by their concentration gradient but are prevented by ϕ. The membrane potential that can balance the concentration gradient of an ion is called the equilibrium potential of the ion. The equilibrium potential can be calculated by the Nernst equation as follows:
where Co and Ci are concentrations of the ion outside and inside the cell, respectively; E is the ion's equilibrium potential; R = 8.31 J/(mol·K) is the gas constant; T is the absolute temperature (K), T = 300K; F = 96,500 J/(V·mol) or C/mol is the Faraday's constant; and Z is the valence charge of the ion.
Let iNa denote the sodium current driven by its concentration gradient and ϕ, and let iK denote the potassium current driven by its concentration gradient and prevented by ϕ. So we have the following:
where gNa is the electric conductance of the membrane ion channels to sodium ions and has the unit of /Ω or S;
gK is the electric conductance of the membrane ion channels to potassium ions and has the unit of /Ω or S;
ϕ is the membrane resting potential defined as ϕ = ϕ(in) − ϕ(out) and has the unit of V;
EK is the potassium equilibrium potential and has the unit of V;
ENa is the sodium equilibrium potential and has the unit of V.
The function of the Na+/K+ pump is to continuously pump sodium ions toward outside the cell and pump potassium ions toward inside the cell to compensate for the consumption of ions used by iNa and iK. Let jNa denote the pump current of sodium ions, and let jK denote the pump current of potassium ions. So it should be that
For the pumping ratio nna/nk = 3/2, jNa = −(3/2)jK.
So iNa = −(3/2)iK, or as in Ref.: 15
gNa(ϕ − ENa) = −(3/2) gK(ϕ − EK).
This equation governs the membrane potential ϕ.
However, as we noted above, the pumping rate of the Na+/K+ pump can be 100 times/s, the density of the Na+/K+ pump can be several thousand pumps/μm2, and the amount of charge required for producing a membrane potential of −70 mV such as in neurons is only about 4350 monovalent ions/μm2. So any accumulation of charges on the membrane by the dipole charging process may cause significant change in the membrane potential because the density and the pumping rate of the Na+/K+ pump are high.
For example, if we assume that the pumping rate is 100 times/s and the density of the Na+/K+ pump is 2000 pumps/μm2, then because the pumping ratio is 3 Na+ to 2 K+, for every second there will be (3 − 2) × 100 × 2000 = 2 × 105 net monovalent ions/μm2 being transported. This will create (2 × 105/4350) × (−70 mV) = −3.2 × 103 mV/s, which is much larger (in absolute value) than −70 mV. So to avoid accumulation of such a large number of charges on the membrane, there should be some mechanisms to regulate the pumping process of the Na+/K+ pump. This includes two aspects.
The pumping process of the Na+/K+ pump can be regarded as two charging processes by two dipoles: the sodium dipole and the potassium dipole. Because the two dipoles have opposite directions, the sodium charging current and the potassium charging current will cancel each other out, but that process should be steady so as to avoid any variation in membrane potential. This is ensured by both Equations (14) and (15). In other words, the sodium component of the sodium charging current and the potassium component of the potassium charging current are equal to each other so that they can cancel each other out steadily, and the potassium component of the sodium charging current and the sodium component of the potassium charging current are compatible with the pumping ratio nk/nna so that they can be used by the Na+/K+ pump again.
After the two dipole charging currents cancel each other out, there will still be some charges left on the membrane. This is because the pumping ratio may not be equal (such as 3 Na+ to 2K+), and not all pumped ions will charge the membrane but some of them will go through ion channels [as i2 in Equation (3)]. So these remaining charges will create a change in the membrane resting potential, and this is known as the electrogenic contribution of the Na+/K+ pump. This electrogenic contribution can be shown by a small depolarization of the membrane potential occurring immediately after the Na+/K+ pump is blocked. 16 It is reported that this electrogenic contribution of the Na+/K+ pump is less than 11 mV, 15 or about 2 to 16 mV, depending on cell type. 16 We can do some calculations to explain this phenomenon.
Let D denote the density of the Na+/K+ pump, and let H denote the pumping rate of the Na+/K+ pump. So the pump current of sodium ions jNa = 3DHF/L, where F = 96,500 C/mol is the Faraday's constant, L = 6.02 × 1023/mol is the Avogadro's number, and 3 indicates three sodium ions being pumped out per pumping cycle. According to Equation (20), iNa = −jNa = −3DHF/L. Substituting iNa into Equation (19), we get rNa = gNa−1 = (ϕ − ENa)/iNa = −(ϕ − ENa)L/(3DHF), where rNa is the electric resistance of the membrane ion channels to sodium ions. The accumulated charges on the membrane caused by the electrogenic contribution of the Na+/K+ pump will discharge through the membrane ion channels. The capacitive time constant for the discharging process of sodium ions can be calculated as follows:
where Cm is the electric capacitance per unit area of the cell membrane as defined in Equation (4).
Similarly, the pump current of potassium ions jK = −2DHF/L, where 2 indicates two potassium ions being pumped in per pumping cycle, and the minus sign means the direction of jK is opposite to that of jNa. According to Equation (20), iK = −jK = 2DHF/L. Substituting iK into Equation (19′), we get rK = gK−1 = (ϕ − EK)/iK = (ϕ − EK)L/(2DHF), where rK is the electric resistance of the membrane ion channels to potassium ions. The capacitive time constant for the discharging process of potassium ions can be calculated as follows:
According to Equation (18), the sodium equilibrium potential is as follows:
and the potassium equilibrium potential is as follows:
Substituting ENa, EK into Equations (21) and (22), we have the following:
We know that for humans, 17 CNa,o = 142 mM, CNa,i = 12 mM, CK,o = 4.3 mM, and CK,i = 139 mM. Assume that the pump density D = 2000 pumps/μm2 = 2.0 × 1015 pumps/m2, the pumping rate H = 100 times/s, ϕ = −70 mV = −7.0 × 10−2 V, and the electric capacitance Cm = 1 μF/cm2 = 1.0 × 10−2 F/m2. Substituting these quantities with R = 8.31 J/(mol·K), T = 300K, F = 96,500 J/(V·mol), L = 6.02 × 1023/mol into Equations (23) and (24), we get τNa = 0.0139 s ≈ 14 ms, and τK = 0.0031 s ≈ 3.1 ms.
We can see that both τNa = 14 ms and τK = 3.1 ms are comparable with the pumping cycle 10 ms (corresponding to pumping rate 100 times/s). Considering here that this is a rough estimation, in a real process it can be regarded that the accumulated charges would be able to discharge within one pumping cycle. Thus the averaged accumulated charges staying on the membrane over one pumping cycle can be estimated as half of the overall accumulated charges in one cycle, which is (1/2) × [(3Na+ − 2K+) × 1 cycle × 2000 pumps/μm2] = 1000 net monovalent ions/μm2. As we have noted, 4350 monovalent ions/μm2 corresponds to −70 mV, so this will create (1000/4350) × (−70 mV) ≈ −16 mV. This value is just what we would expect for the electrogenic contribution of the Na+/K+ pump. 16
We can also see that, in Equations (21) and (22), τNa and τK are functions of variables ENa, EK, ϕ (and therefore rNa, rK), D, H, and Cm; in Equations (23) and (24), τNa and τK are even functions of variables CNa,o, CNa,i, CK,o, CK,i, ϕ, D, H, and Cm. The cell can adjust all these variables so that the accumulated charges would discharge in time and the direct electrogenic contribution of the Na+/K+ pump to the membrane resting potential is small.
Effectiveness and prospect of the dipole-charging model
Our current knowledge about the Na+/K+ pump remains at the stage where much attention is paid to the structural and functional study of the Na+/K+ pump itself, but less attention is paid to the relationship between the function of the Na+/K+ pump and other cellular functions. The main reason for this is that we are unaware of what the pumping process is and why the Na+/K+ pump takes 3 Na+ to 2 K+ as the pumping ratio.
The proposal of the dipole-charging model, which takes the pumping process as a dipole-charging process and makes a complete description accordingly, solves these questions. It not only shows that the 3 Na+ to 2 K+ pumping ratio is due to the ratio of the ionic mobilities of K+ to Na+ ions, but also links the pumping ratio, the pumping rate, the pump density, and other cellular functional parameters, such as the intracellular and extracellular concentrations of Na+ and K+ ions and the membrane capacitance, therefore revealing the relationship between the function of the Na+/K+ pump and other cellular functions, such as the membrane potential and the osmotic balance condition. The effectiveness of the dipole-charging model can be shown by the predictions it makes in the following three aspects:
On the condition that the pumping ratio nna/nk = 3/2, the dipole-charging model predicts for the first time that the concentration ratios CK,i/CNa,o ≈ 1.0 and CK,o/CNa,i ≈ 0.30.
6
The observation values of CK,i/CNa,o and CK,o/CNa,i for different cell types are shown to be consistent with these two predicted values (see Table 1 in Ref.
6
). In this article, I further propose that the concentration equivalence CK,i/CNa,o = 1 can ensure the osmotic pressure balance of the cell and can be regarded as osmotic balance condition [Equation (16)]. The dipole-charging model gives the result that CK,i/CNa,o = 1 when and only when nna/nk = UK/UNa ≈ 3/2 [Equation (17)]. For other pumping ratios, the corresponding values of CK,i/CNa,o are far away from 1 (Table 1), and they cannot ensure the osmotic balance condition. So the pumping ratio of 3 Na+ to 2 K+ is shown to be determined by the ratio of the ionic mobilities of K+ to Na+ ions. According to the dipole-charging model, the pumping process of the Na+/K+ pump can be regarded as two charging processes by the sodium dipole and the potassium dipole. After the cancellation of the two dipole charging currents, there will still be some charges left on the membrane. This will create a small change in the membrane resting potential, which is known as the electrogenic contribution of the Na+/K+ pump. These accumulated charges on the membrane should discharge through the membrane ion channels in time to ensure that the change created by these charges in the membrane potential is small. This is the mechanism I propose in this article to explain the electrogenic contribution of the Na+/K+ pump. The dipole-charging model can be used to explain this mechanism. As we have calculated, when the pump density D = 2000 pumps/μm2 and the pumping rate H = 100 times/s, the capacitive time constant for the discharging process of sodium ions is τNa = 14 ms and the capacitive time constant for the discharging process of potassium ions is τK = 3.1 ms. Both of them are comparable with the pumping cycle 10 ms (corresponding to the pumping rate 100 times/s). So the accumulated charges on the membrane should be able to discharge through the membrane ion channels in the time of one pumping cycle to ensure that this direct electrogenic contribution of the Na+/K+ pump is small. So this mechanism is supported by the dipole-charging model.
The dipole-charging model provides a complete description of the pumping process of the Na+/K+ pump. Any phenomenon related to the pumping process, such as the electrogenic contribution of the Na+/K+ pump, can be explained by this model. So the dipole-charging model would be applicable in medicinal practice. For example, under pathological conditions such as transient ischemia or hypoxia, the cell will undergo depolarization, and the electrogenic contribution of the Na+/K+ pump may affect this process.16,17 This electrogenic contribution may also affect automaticity of the nodal cells of the heart, as well as of other cells that exhibit automaticity. 16 In these cases, we may need to know what factors would influence the electrogenic contribution of the Na+/K+ pump, and what kind of influences these factors may have. Equations (21) to (24) provide information for us.
As already noted, τNa and τK are functions of variables ENa, EK, or CNa,o, CNa,i, CK,o, CK,i, and ϕ (and therefore rNa, rK), D, H, and Cm. For example, the membrane capacitance Cm is due to the lipid bilayer matrix. Under pathological conditions, the structural and chemical composition of the cell membrane may be changed, and the capacitance Cm will change accordingly. This has influence on τNa and therefore on the electrogenic contribution of the Na+/K+ pump. In addition, some drugs and toxins exert primary or secondary effects on the electrical properties of the cell membrane. For an understanding of the mode of action of these therapeutic drugs, toxic agents, neurotransmitters, hormones, and plasma electrolytes, it is necessary to understand the electrical properties and behavior of the cell membrane. 16 The dipole-charging model would be useful in these areas. So the dipole-charging model has a broad prospect in medicinal and biological applications.
The pumping ratio of the Na+/K+ pump is not due to the energy requirement
There is an opinion that the pumping ratio of the Na+/K+ pump is due to the energy requirement. In Ref.,
6
I have discussed the free energy change of the coupled overall reaction for the Na+/K+ pump [Reaction (26) in Ref.
6
]:
where “in” indicates intracellular and “out” indicates extracellular, and ATP, adenosine diphosphate (ADP), and Pi are in the intracellular solution.
The free energy change of Reaction (25) is [Equation (39) in Ref.
6
]:
where KATP is the equilibrium constant for ATP hydrolysis, ϕ is the membrane potential as defined before, and CATP, CADP, CPi denote the concentrations of ATP, ADP, and Pi, respectively.
The free energy used for pumping 1 mol Na+ ions, ΔGNa, and for pumping 1 mol K+ ions, ΔGK [Equation (42) in Ref.
6
], can be obviously obtained from Equation (26):
where ENa = (RT/F)ln[(CNa,o)/(CNa,i)] >0 and EK = (RT/F)ln[(CK,o)/(CK,i)] <0 are equilibrium potentials for Na+ and K+ ions, respectively; ϕ = ϕ(in) − ϕ(out) <0, and ZNa = ZK = 1. Two cases have been calculated in Ref., 6 for mammalian skeletal muscle and for the squid axon. For mammalian skeletal muscle, CNa,i = 12 mM, CNa,o = 145 mM, CK,i = 155 mM, CK,o = 4 mM, and ϕ = −80 mV, and we have ΔGNa = 13.9 kJ/mol and ΔGK = 1.40 kJ/mol. For the squid axon, CNa,i = 50 mM, CNa,o = 440 mM, CK,i = 400 mM, CK,o = 20 mM, and ϕ = −70 mV, and we have ΔGNa = 12.2 kJ/mol and ΔGK = 0.713 kJ/mol.
These results show that the free energy used for pumping 1 mol Na+ ions is about 10-fold larger than that for pumping 1 mol K+ ions. This is because the K+ equilibrium potential EK is close to the resting potential ϕ, and the resting potential favors the pumping of K+ ions from outside to inside the cell [Equation (26′)]. So if only energy requirement is considered, the number of K+ ions pumped in each cycle by the Na+/K+ pump can actually be arbitrary.
In this study, I make a further discussion on the use of the free energy in Equation (26). Let
ΔGion is the free energy used for pumping 3 mol Na+ ions from inside to outside the cell and 2 mol K+ ions from outside to inside the cell, corresponding to the pumping ratio of 3 Na+ to 2 K+. Apparently, ΔGion depends not only on the pumping ratio but also on the concentrations of Na+ and K+ ions in the intracellular and the extracellular solutions. For example, using the data above for mammalian skeletal muscle and for the squid axon, we can calculate that the ΔGion for the mammalian skeletal muscle is 44.6 kJ/mol and the ΔGion for the squid axon is 38.0 kJ/mol. The difference between these two values of ΔGion is 6.6 kJ/mol, which is caused mainly by the difference in the intracellular and extracellular concentrations of Na+ and K+ ions between the mammalian skeletal muscle and the squid axon (the difference in the membrane potential ϕ makes a little contribution). This 6.6 kJ/mol difference is apparently large enough to allow more K+ ions to be pumped in each cycle by the Na+/K+ pump in case of the squid axon (ΔGK = 0.713 kJ/mol as above), but actually it does not cause any change in the pumping ratio in the squid axon. This shows that the pumping ratio of the Na+/K+ pump is not due to the energy requirement.
Moreover, as we can see from Tables 4–6 in Ref., 6 the extracellular ion concentrations decrease dramatically from marine animals to freshwater and terrestrial animals, with those of marine animals being similar to those of seawater. As discussed above, a change in ion concentrations would cause a change in ΔGion. So if the pumping ratio were dependent on the energy requirement, such a change in ion concentrations or in ΔGion should have given rise to mutation of pumping ratio during evolution. In another way we can say that the ion concentrations of the body fluid could adjust themselves to any pumping ratio within certain energy available from the hydrolysis of 1 mole ATP. In such cases, the number of Na+ ions pumped in each cycle by the Na+/K+ pump could also be varied, just like that the number of K+ ions could be varied in the above cases of the mammalian skeletal muscle and the squid axon. However, as we know, the Na+/K+ pump only takes 3 Na+ to 2 K+ as the pumping ratio. So we can say that the energy requirement is not the factor that determines the pumping ratio of the Na+/K+ pump during evolution.
Some reports on different pumping ratios of the Na+/K+ pump
There have been some reports on different pumping ratios of the Na+/K+ pump. For example, Ref. 18 reported a conversion of the normal 3 Na+/2 K+/1 ATP stoichiometry of the Na+/K+-ATPase to electroneutral 2 Na+/2 K+/1 ATP stoichiometry. By sequence comparing of the electrogenic Na+/K+-ATPase and the nonelectrogenic H+/K+-ATPase, the researchers found that a cysteine conserved in the α-subunit transmembrane helix M8 of all Na+/K+-ATPases was replaced by arginine in H+/K+-ATPases at the corresponding position. Replacement of this cysteine C932 (rat α1 Na+/K+-ATPase numbering) in the transmembrane helix M8 of the Na+/K+-ATPase with arginine converted the 3 Na+/2 K+/1 ATP stoichiometry of the Na+, K+-ATPase to 2 Na+/2 K+/1 ATP stoichiometry, similar to the electroneutral transport mode of the H+/K+-ATPase. The mechanism is that the guanidium group of the M8 arginine, which works as an internal cation, prevents Na+ binding at the third Na+-specific site, while allowing Na+ to bind at the two other sites and become transported. For our purposes, this work provides the structural basis for 2 Na+ to 2 K+ pumping ratio of the Na+/K+ pump, which means that during evolution, the occurrence of a 3 Na+ to 2 K+ pumping ratio, rather than a 2 Na+ to 2 K+ pumping ratio, is not due to the structure restriction of the Na+/K+ pump.
Furthermore, Ref. 18 found that the substitution of this M8 cysteine of the Na+/K+-ATPase with phenylalanine led to spectacular reductions of apparent Na+ affinity, which could explain the function of its presence in a patient with alternating hemiplegia of childhood. 19 These facts indicate that the mutation of this M8 cysteine of the Na+/K+-ATPase to arginine may also have appeared in the course of evolution but has not been conserved because it might be a more severe mutation than that of the M8 cysteine to phenylalanine.
Another exception is found in the brine shrimp Artemia, which can thrive at extreme salt concentrations in hypersaline lakes. 20 The shrimp drinks seawater, excreting salt from salt glands and maintaining low salt concentrations (155–185 mM) in the extracellular hemolymph fluid.21,22 There are two isoforms of the Na+/K+-ATPase in Artemia franciscana, the α1-NN-subunit 23 and the lysine-substituted α2-KK-subunit. 24 In the α2-KK-subunit, two lysines are substituted for two asparagines in the transmembrane segments M4 and M5, introducing two positively charged ɛ-amino groups of lysine near the center of the binding cavities for cations, 25 which interfere with the binding of Na+ and K+ ions. 20 The lysine substitutions of α2-KK-Na+/K+-ATPase act as fixed internal cations to reduce the number of ions transported per ATP hydrolyzed and enable the Na+/K+ pump to extrude Na+ ions against steeper gradients.
The α2-KK-subunit is almost exclusively located in the salt gland of Artemia larvae, while the α1-NN-subunit is ubiquitous. 26 An increase of the α2 mRNA levels was noticed when salinity increased, whereas expression of α1 remained unchanged. 20 Refs.27,28 further determined the pumping ratio of the Artemia α2-KK-Na+/K+-ATPase to be 2 Na+ to 1 K+ by studying the functional effects of the equivalent asparagine-to-lysine substitutions in the Xenopus. However, the α2-KK-Na+/K+-ATPase with this pumping ratio only exists in the salt gland of Artemia, and its function is to extrude Na+ ions against steeper gradients in the salt gland, which contributes to the salt adaptation of Artemia. In most cases in Artemia, the α1-NN-Na+/K+-ATPase with the normal 3 Na+ to 2 K+ pumping ratio plays a central role in cellular function.
Conclusion
In this article, the dipole-charging model is extended to obtain a universal group of Equations (14) and (15) that relate the pumping ratio of the Na+/K+ pump, the ionic mobilities of K+ and Na+ ions, and the intracellular and the extracellular concentrations of sodium and potassium ions. By combining with the osmotic balance condition CK,i/CNa,o = 1, the pumping ratio of 3 Na+ to 2 K+ can be determined as the ratio of the ionic mobilities of K+ to Na+ ions. The dipole-charging model is shown to be useful in medicinal and biological applications. Further analysis of the free energy change of the coupled overall reaction for the Na+/K+ pump shows that the pumping ratio of the Na+/K+ pump is not due to the energy requirement. Some reports on different pumping ratios of the Na+/K+ pump are also explained.
Footnotes
Author's Contribution
N.X. designed the research, performed the research, and wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.