
Abstract
Background:
Tissues, such as skeletal muscle, have been targeted for the delivery of plasmid DNA (pDNA) encoding vaccines and therapeutics. The application of electric pulses (electroporation or electrotransfer) increases cell membrane permeability to enhance plasmid delivery and expression. However, the molecular effects of DNA electrotransfer on the muscle tissue are poorly characterized.
Materials and Methods:
Four hours after intramuscular plasmid electrotransfer, we evaluated gene expression changes by RNA sequencing. Differentially expressed genes were analyzed by gene ontology (GO) pathway enrichment analysis.
Results:
GO analysis highlighted many enriched molecular functions. The terms regulated by pulse application were related to muscle stress, the cytoskeleton and inflammation. The terms regulated by pDNA injection were related to a DNA-directed response and its control. Several terms regulated by pDNA electrotransfer were similar to those regulated by pulse application. However, the terms related to pDNA injection differed, focusing on entry of the plasmid into the cells and intracellular trafficking.
Conclusion:
Each muscle stimulus resulted in specific regulated molecular functions. Identifying the unique intrinsic molecular changes driven by intramuscular DNA electrotransfer will aid in the design of preventative and therapeutic gene therapies.
Introduction
The application of external electric fields (electroporation or electrotransfer) is an option for the delivery of nonviral nucleic acid-based therapeutics and vaccines into tissues such as skeletal muscle. While plasmid DNA (pDNA) enters all skeletal muscle cell types to express the encoded protein, regenerated rather than preexisting muscle fibers are primarily responsible for long-term gene expression. 1 Pulse application may induce muscle damage, which initiates signaling and the formation of new muscle fibers by the proliferation of satellite cells. 2 In addition, pDNA is an activator of fundamental cellular responses through the activation of intracellular DNA-specific pattern recognition receptors.3–5 Our previous studies have focused specifically on immune effects in response to nucleic acid electrotransfer in cells in vitro,6–9 3D models,10,11 tumors,12–15 and normal tissues.16,17
Skeletal muscle RNA sequencing data were previously analyzed for differentially expressed genes (DEGs) focusing on inflammatory pathways. 17 In this study, we used a broad approach, gene ontology (GO) analysis, to identify the molecular functions modulated by the delivery of pDNA with external electric fields. Annotations in GO databases may be manual or automated and are based on several types of evidence. These annotations incompletely represent current knowledge and are inherently biased. 18 DNA electrotransfer combines two components that may activate molecular functions, defined as “the activity of a gene product at the molecular level,” 19 DNA injection and pulse application. In spite of the limitations of GO databases, we found that the most significantly regulated terms 4 h after intramuscular DNA electrotransfer were compatible with DNA electrotransfer and its individual components.
Materials and Methods
pDNA delivery and animals
This analysis used previously described RNA sequencing data. 17 University of South Florida Institutional Animal Care and Use Committee approved all protocols (protocol R IS00007249, 2019). For the study, 7- to 8-week-old female C57Bl/6J mice (Jackson Laboratories, Bar Harbor, ME) were used. Briefly, for the experimental group, a plasmid that did not encode a coding sequence expressed by mammalian cells, gWiz Blank was injected into the right caudal thigh muscle followed by the application of eight 20-millisecond pulses with a 2-needle medical-grade stainless steel electrode with a 5 mm gap at a voltage to distance ratio of 100 V/cm (BTX Harvard Apparatus, Holliston, MA) and a frequency of 1 Hertz with an ECM830 pulse generator (BTX). In the control group, muscles were not manipulated. In the pDNA groups, the muscle tissue was injected with gWiz Blank and in the electroporation-only group the muscle was subjected to the application of pulses. Animals were euthanized after 4 h, and muscle samples were collected and RNA sequenced. 17
In the previous study, statistical evaluation was performed to identify DEGs in pairwise comparisons using DESeq2 20 and focusing on inflammatory pathways assigned through Kyoto Encyclopedia of Genes and Genomes (KEGG)21–24 using the R package GSAR. 25 In the present study, GO analysis 26–28 was performed to identify functional enrichment in DEGs using topGO Version 2.42.0 29 using the weight algorithm and Fisher's exact test; p-values less than or equal to 0.05 were considered statistically significant. The weight algorithm is designed to eliminate nonspecific terms while prioritizing terms supported by other terms in the GO graph network. Gene symbols in the text are as per the Mouse Genome Database. 30
Results and Discussion
Genome-wide expression analysis was performed for a skeletal muscle control group and mice that received intramuscular control vector pDNA injection, pulse application alone, or pDNA injection followed by electrotransfer to identify differentially regulated genes. GO enrichment analysis identified 22 molecular functions upregulated by electroporation alone (Fig. 1). Toll-like receptor 4 binding, myogenic regulatory factor (MRF) binding, and 2′-5′-oligoadenylate synthetase (OAS) activity were the three most enriched terms.
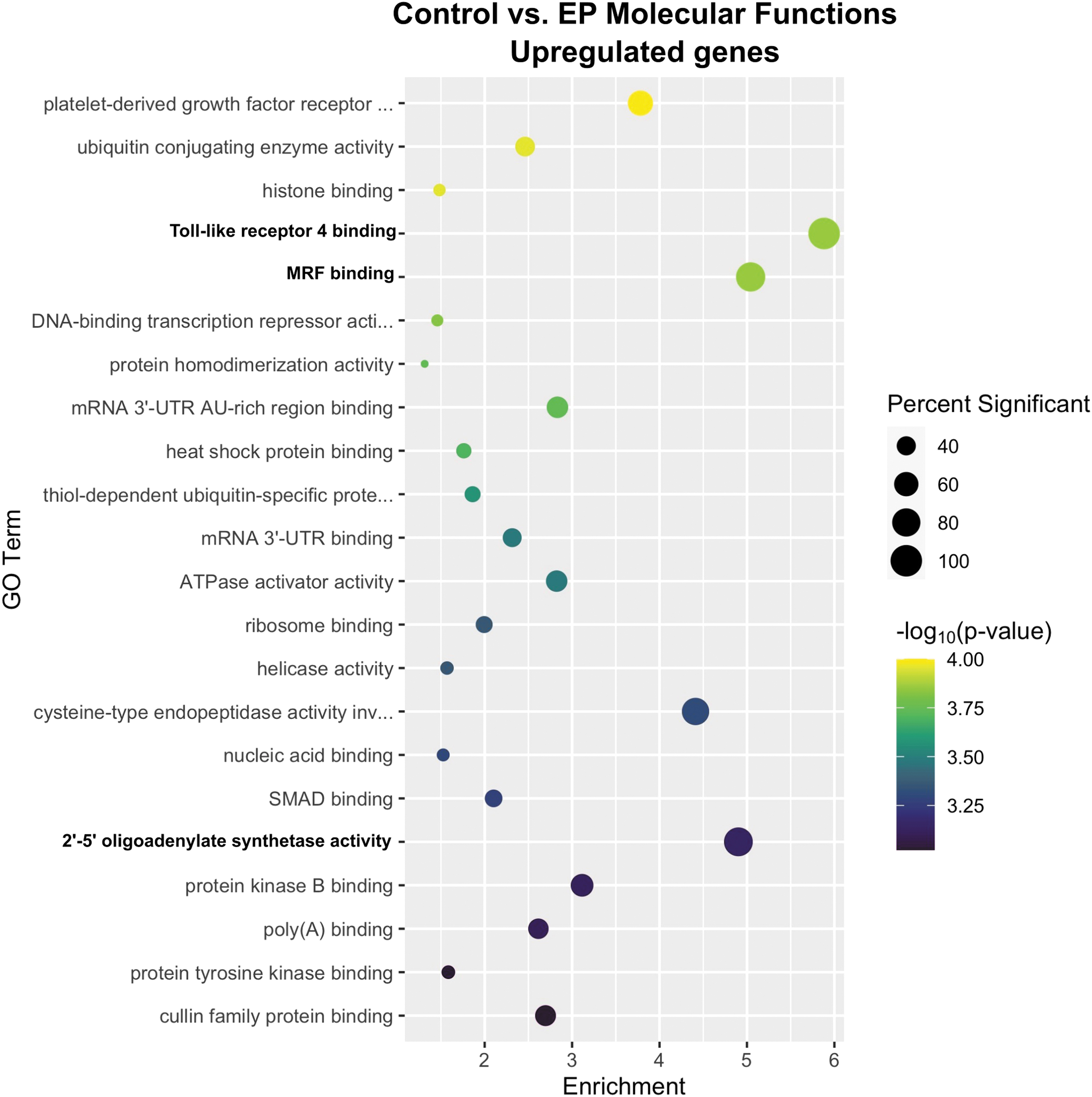
Enrichment profiles for the primary upregulated molecular functions between the control and electroporation-only groups. The size of each circle indicates the percentage of genes in that category that were differentially expressed and the color indicates −log10 (p-value) of DEGs annotated in the specific GO biological process. The x-axis shows enrichment. Enrichment is defined as the number of DEGs in that category divided by the number of genes in that category expected to be found by chance (Enrichment = Significant/Expected). It is a measure of how many more times genes within a term were found than expected by chance. Terms with a p-value ≤0.05 included in the graph with terms sorted from lowest significant p-value to highest significant p-value. n = 4–5 per group. DEG, differentially expressed gene; EP, electroporation; GO, gene ontology.
The most regulated genes in the toll-like receptor 4 binding term were lymphocyte antigen 96 (Ly96 or MD2) and S100 calcium-binding protein A8 (S100 A8 or calgranulin A). Ly96 complexes with TLR4 to activate an inflammatory cascade. 31 The calcium sensor S100 A8 is expressed by neutrophils and monocytes. 32 Extracellular S100 A8 is an alarmin that also binds TLR4 to activate an inflammatory response. 33 The gene for TLR4 is not included in this term. The classical ligand for TLR4 is lipopolysaccharide, but this receptor may also be indirectly activated by a number of damage-associated molecular patterns such as oxidized phospholipid, β-defensin, high-mobility group protein 1, heat shock proteins, heparin sulfate, and hyaluronic acid. 31 This group received pulses only; lipopolysaccharide was not a component. Therefore, it is possible that an alternative TLR4 ligand is initiating an inflammatory process. The mRNAs of a number of proinflammatory chemokine RNAs are regulated by pulse application; C-C motif chemokine 2 (CCL2) protein is produced in electroporated muscle. 17
The MRFs Myf5, MyoD, MyoG, and MRF4 34 are basic helix-loop-helix transcription factors controlled by MAPK signaling and play a role in satellite cell function to regenerate skeletal muscle35,36 as well as muscle maintenance. 37 The most highly regulated gene was the zinc finger transcription factor Kruppel-like factor 5, which is an essential regulator of muscle differentiation acting in concert with myogenic transcription factors such as MyoD and Mef2. 38 Skeletal muscle regeneration may be regulated in response to muscle damage from pulse application.
The OAS family was originally described as antiviral proteins activated by the genome of double-stranded RNA (dsRNA) viruses. 39 Since dsRNA may also be produced endogenously, these proteins may be activated upon the stress-mediated escape of dsRNA from mitochondria or nuclei after pulse application. 40
GO enrichment analysis identified 35 molecular functions downregulated by electroporation only (Fig. 2). The three most regulated terms were ADP-ribosylation factor binding, RNA polymerase I core promoter sequence-specific DNA binding, and Lin-11, Isl1, and MEC-3 (LIM) domain binding.
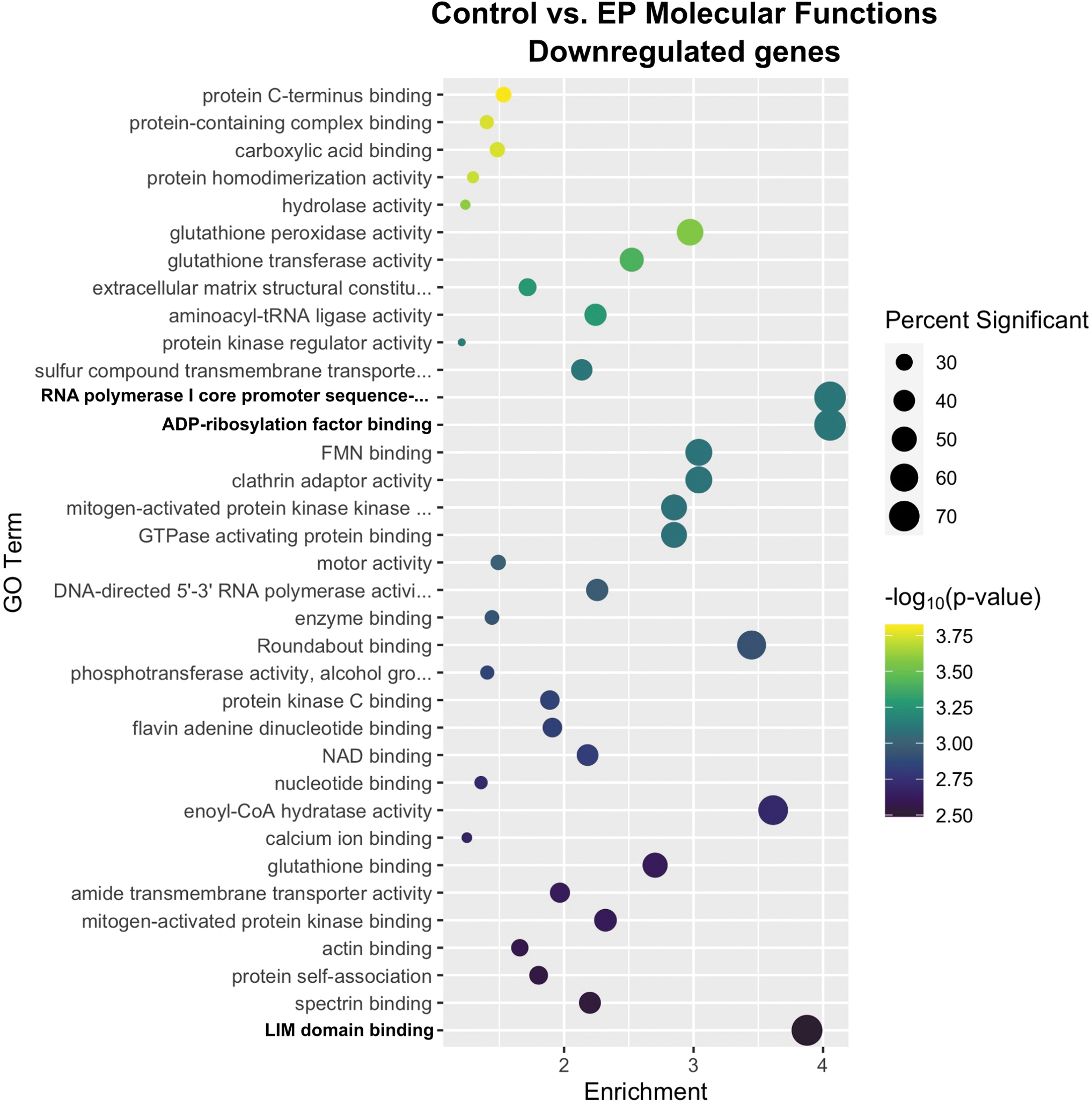
Enrichment profiles for the primary downregulated molecular functions between the control and electroporation-only groups. Interpretation as described in Fig. 1. n = 4–5 per group.
ADP-ribosylation factors are small GTP-binding proteins 41 that reversibly modify proteins and to control a myriad of cellular processes, including immune responses. 42 These factors can activate or suppress inflammatory pathways. 43 This term is likely regulated in response to the innate immune response upregulated by pulse application alone.
The negative regulation of genes encoding RNA polymerase I core promoter accessory factors, particularly those within the preinitiation complex such as the downregulated gene TATA box-binding protein-associated factor RNA polymerase I subunit C (Taf1c), 44 implicates a reduction in ribosomal RNA synthesis in the nucleolus. This reduction may generally control protein synthesis, 45 but is difficult to deduce the precise purpose of negative regulation with respect to the skeletal muscle response to pulse delivery.
LIM domain-containing proteins are mechanosensors that promiscuously bind many proteins 46 and are involved in several biological functions, including cytoskeletal reorganization. 47 These proteins bind to stressed actin filaments 48 and participate in actin polymerization. 49 Several groups have described transient cytoskeletal perturbations within cells in response to pulse delivery,15,50–57 including gene expression changes. 15
After pDNA injection, lipoteichoic acid binding, lipopeptide binding, and CCR2 and CCR3 chemokine receptor binding, which significantly overlap in function, were the most highly enriched terms (Fig. 3). Upregulation of the genes triggering receptor expressed on myeloid cells 2 (TREM2), TREM 3, and TLR2 in the lipoteichoic acid and lipopeptide-binding terms indicates the influx of myeloid cells such as monocytes and macrophages into the muscle.58,59 While chemokine receptor RNAs regulated in the molecular function analysis, the CCR2 ligands, CCL7 and CCL12, and the CCR3 ligand CCL24,60,61 which are chemotactic for these cells, were upregulated. Chemokine receptors such as CCR3 may also participate in muscle regeneration.62,63 Several related chemokine receptor terms were also upregulated in this group.
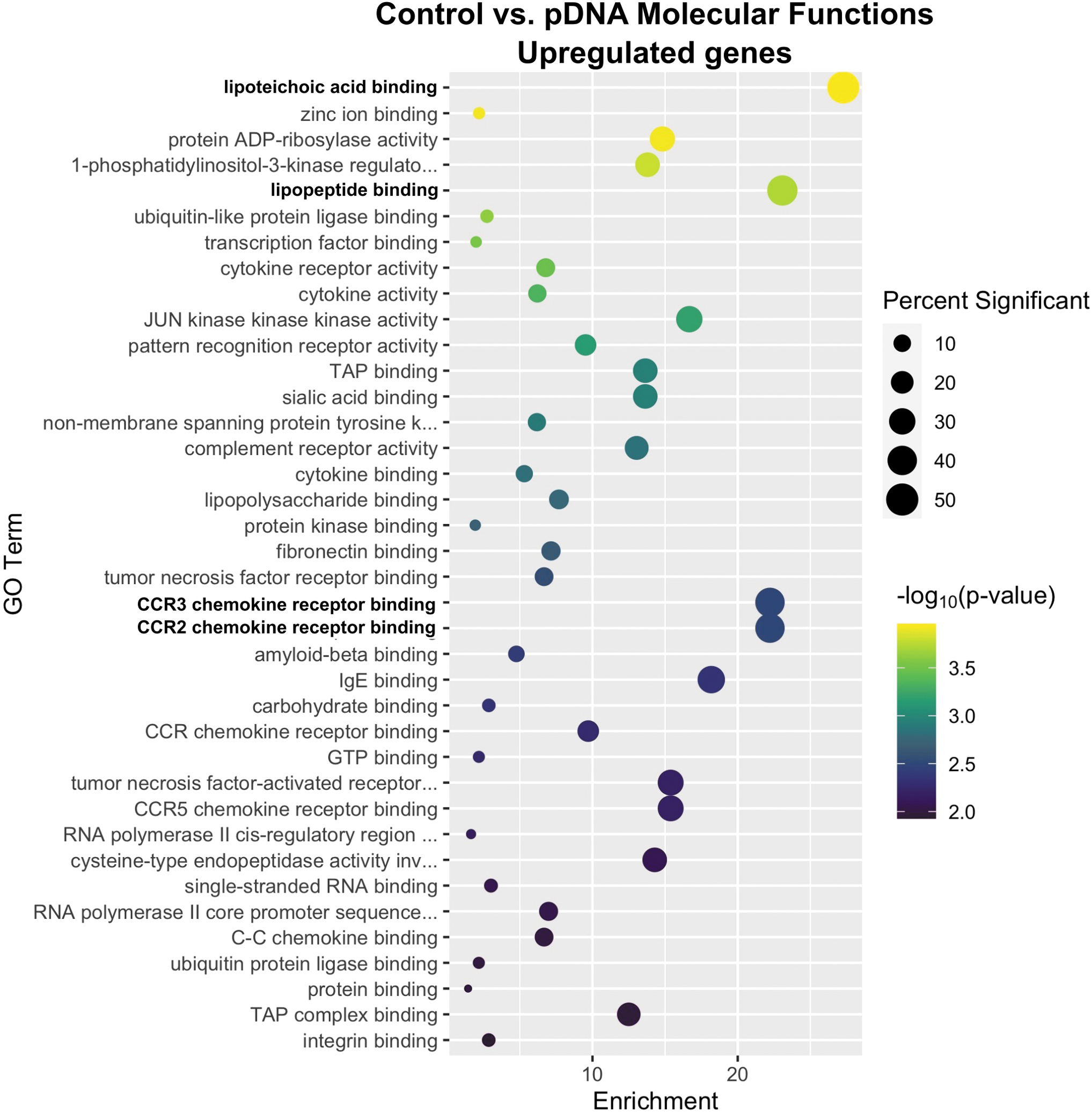
Enrichment profiles for the primary upregulated molecular functions between the control and plasmid injection-only groups. Interpretation as described in Fig. 1. n = 4–5 per group.
GO enrichment analysis identified 50 molecular functions downregulated by plasmid injection (Fig. 4). Natriuretic peptide receptor activity was highly enriched, whereas 13 secondary terms were equivalently enriched. Natriuretic peptides and their three receptors (NPR1, NPR2, NPR3) have diverse autocrine and paracrine functions. 64 NPR1 and NPR2 are guanylyl cyclases while NPR3 functions in peptide clearance. 65 This activity is well characterized in the heart; few studies have been performed in skeletal muscle. In this tissue, NPR1 expression correlates with oxidative metabolism before and after chronic exercise. 66 Since these peptides may be anti-inflammatory, 67 they may be released in response to the chemokine expression associated with pDNA injection. 17 Receptor gene expression is negatively regulated by agonist binding, 65 which may explain the downregulation of NPR gene expression.
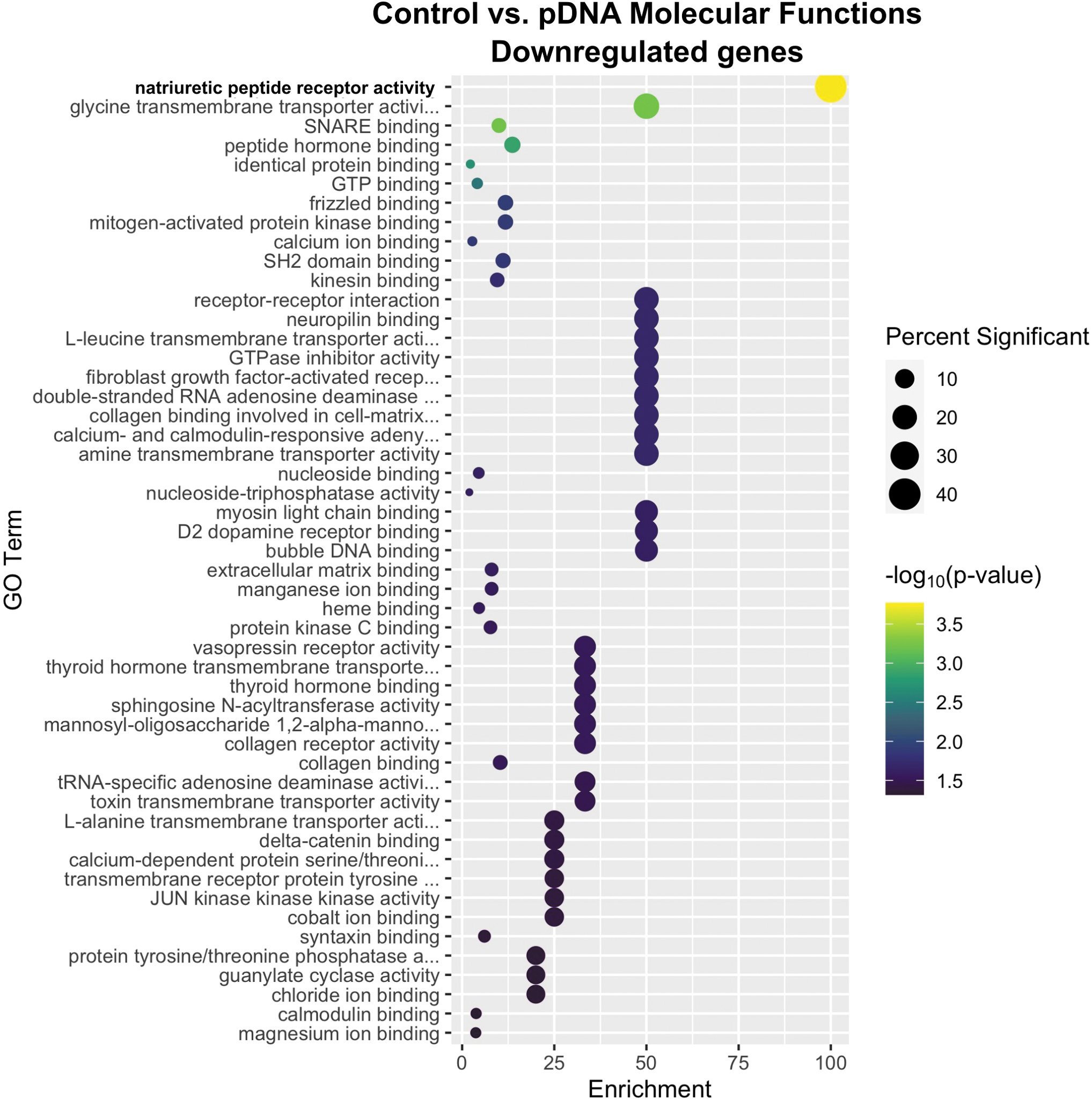
Enrichment profiles for the primary downregulated molecular functions between the control and plasmid injection-only groups. Interpretation as described in Fig. 1. n = 4–5 per group.
GO enrichment analysis identified 13 molecular functions upregulated by pDNA electrotransfer (Fig. 5). The three most significantly enriched terms were MRF binding, nuclear import signal receptor activity, and ATPase activator activity.
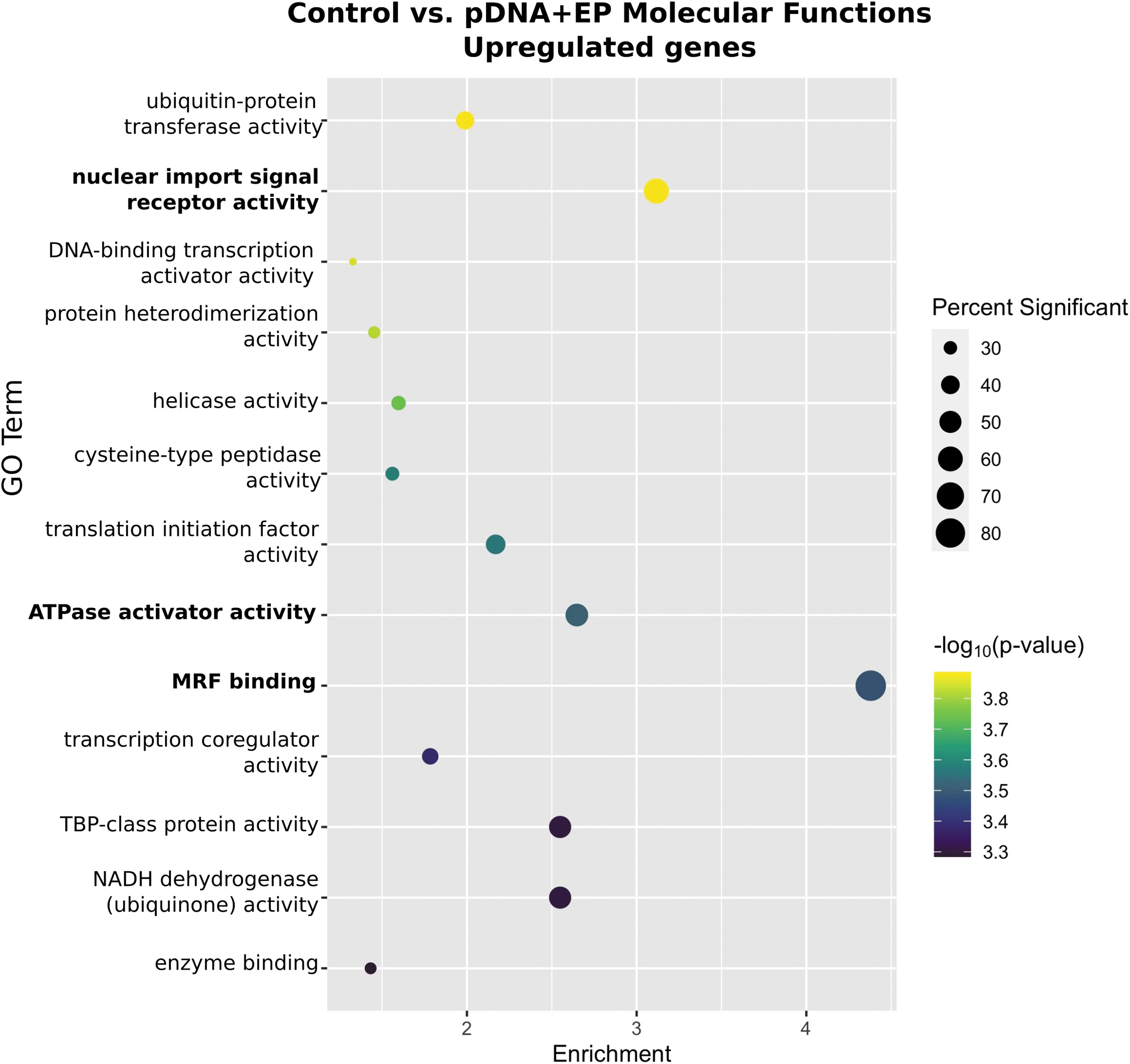
Enrichment profiles for the primary upregulated molecular functions between the control and plasmid electroporation groups. Interpretation as described in Fig. 1. n = 4–5 per group.
Similar to electroporation alone (Fig. 1), MRF binding was a highly enriched GO term with pDNA electrotransfer. This confirms the importance of this term to muscle regeneration after pulse delivery.
Nuclear import signal receptor activity was upregulated, which is likely a consequence of nuclear entry by the plasmid. Plasmid import into the nucleus requires encoded DNA nuclear targeting sequences (DTSs),68,69 which may be cell specific. 70 While plasmid easily enters the nucleus during mitosis, transcription factors bind these sequences to mediate nuclear entry of DNA in nondividing cells. 71 GO terms related to transcription factor activity, including DNA-binding transcription activator activity and transcription coregulatory activity, were significantly upregulated to a lesser degree.
Several general DTSs have been described. 72 We compared the gWiz Blank sequence with these general sequences using the default settings of EMBOSS Needle. 73 This plasmid lacked the 72 base-pair SV40 enhancer sequence68,74 or the longer glucocorticoid hormone receptor-binding sequence. 75 However, the glucocorticoid hormone receptor-binding sequence encodes two hexamers with a three-nucleotide spacer (GGTACAnnnTGTTCT) 76 and a similar element (GCTACAGAGTTCTT) is located in the plasmid's origin of replication. NFkB 77 is known to activate the CMV enhancer, 78 and an NFkB-binding site (GGGACTTTCC) 79 is located in the CMV promoter and intron of gWiz Blank. Finally, nuclear entry can be driven by hypoxia-responsive elements. 80 The CMV promoter is intrinsically hypoxia sensitive 81 ; two hypoxia response consensus elements (A/GCGTG) 82 are encoded in the CMV promoter and intron. Taken together, it is conceivable that the nuclear localization of gWiz Blank and related vectors is driven by glucocorticoid hormone receptor, NFkB, or by hypoxia-inducible factor binding.
In smooth muscle, importin b1, importin 7, the GTPase Ran, Ran-binding protein P1 (RanBP1), and RanBP2 bind DTSs. 83 A study in a cancer cell line confirmed a large number of protein-bound pDNA, including importins, Ran-binding proteins, microtubule-based motor proteins, transcription factors, heterogeneous nuclear ribonucleoproteins, heat shock chaperones, and histones. 84 We found a number of regulated importin and transportin mRNAs as well as the mRNAs of a RAN-binding protein and the chaperone Hikeshi (Fig. 2a), indirectly supporting these observations.
ATPase activator activity 85 provides ATP for contraction and cellular homeostasis. The mRNAs of several subunits of Na+/K+ ATPase are regulated, indicating that this pathway may be activated during muscle regeneration. 85
GO analysis revealed that 29 molecular functions were downregulated by pDNA electrotransfer (Fig. 6). Platelet-derived growth factor (PDGF) binding, clathrin adaptor activity, and glutathione peroxidase activity were the three most downregulated terms after pDNA electrotransfer. Interestingly, glutathione peroxidase activity was also significantly regulated by pulse application but to a lesser degree. Additionally, PDGF binding, downregulated in this group, was an upregulated term after pulse application alone.
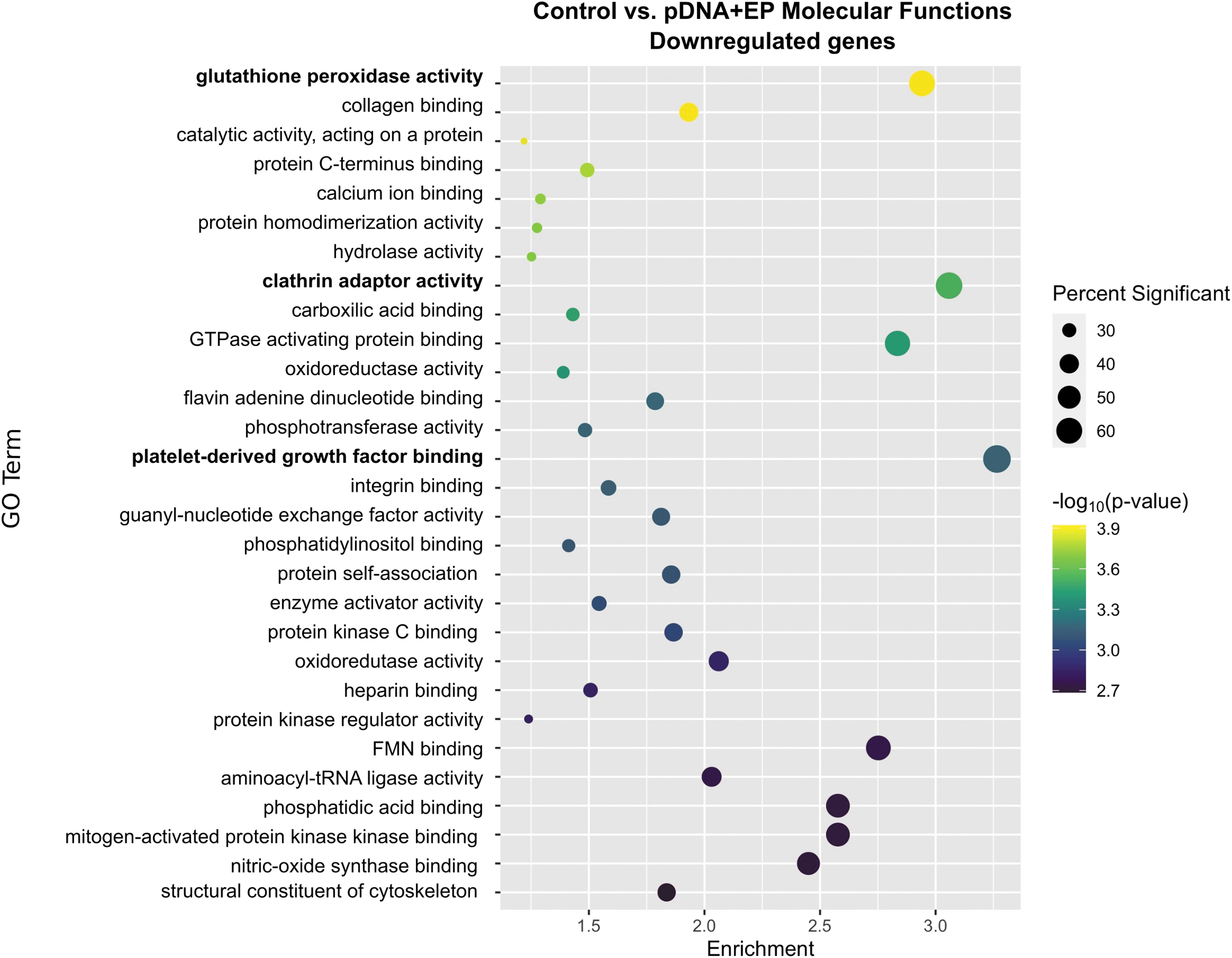
Enrichment profiles for the primary downregulated molecular functions between the control and plasmid electroporation groups. Interpretation as described in Fig. 1. n = 4–5 per group.
PDGF binding was the most enriched and significantly downregulated GO term in this group. Regulated DEGs in the PDGF-binding pathway is shown in Figure 6. While PDGF alpha was slightly upregulated (p < 0.05), other mRNAs associated with PDGF signaling were downregulated. PDGF is known to stimulate the secretion of basement membrane proteins, including Type IV collagen, in skeletal muscle. 86 We found that the mRNAs of several collagen genes are downregulated. The regulation of PDGF binding after pDNA electrotransfer could suggest a recovery mechanism from muscle damage. PDGF signaling is reduced with myogenic differentiation. 87
Broadly, endocytosis defines several mechanisms by which cells internalize cargo molecules, 88 including the clathrin-mediated endocytic pathway. Over 50 proteins participate in clathrin-mediated endocytosis. 89 These mechanisms play an important role for pDNA entry during transfection by electrotransfer,90,91 including in muscle. 92 Treatment with chlorpromazine, monodansylcadaverine, or dynasore, inhibitors of clathrin-mediated endocytosis, significantly inhibits plasmid entry into cells 90 and subsequent gene expression. 93 Within cells, plasmid colocalizes with markers of clathrin-mediated endocytosis. 91 Knockdown of clathrin heavy chain by siRNA affects transfection efficiency. 94 We detected both upregulated and downregulated genes encoding primarily adapter proteins, although in sum, this pathway downregulation may be the aftermath of the plasmid endocytosis driven by electric pulse application. While glutathione-S-transferase A1 (gsta1) mRNA is upregulated, the remaining regulated genes in the GO term glutathione peroxidase activity were downregulated.
Glutathione-S-transferase conjugates glutathione to a variety of substances; glutathione peroxidase is highly specific to glutathione and catalyzes the reduction of H2O2 or organic hydroperoxides. 95 This enzyme is one of the primary cellular antioxidant enzymes and is inducible in skeletal muscle, particularly after contraction during high-intensity exercise. 96 This pathway may have been initially upregulated in response to the induction of reactive oxygen species, which has been described during pulse application,97,98 but downregulated after 4 h during muscle recovery.
Conclusion
Each muscle manipulation generated specific molecular function changes. Several regulated terms in the pulse application group overlapped those in the pDNA electrotransfer group. However, in some cases, different terms were regulated between the pulse application, pDNA injection, and pDNA electrotransfer groups. However, in each case, the regulated terms were relevant to the group. In spite of the ambiguity that can be associated with GO analysis, the regulated terms tended to agree with previously published observations.
Footnotes
Authors' Contributions
A.S.C.: Validation, Formal Analysis, Investigation, Data curation, Writing—Original Draft, Data curation, Writing—Review & Editing, and Visualization. J.T.: Validation, Investigation, and Writing—Review & Editing. K.K.: Validation, Investigation, and Writing—Review & Editing. M.Z.: Validation, Investigation, and Writing—Review & Editing. J.G.: Validation, Formal Analysis, Data curation, Writing—Review & Editing, and Visualization. L.H.: Conceptualization, Methodology, Validation, Writing—Review & Editing Supervision, and Funding acquisition.
Disclaimer
The content is solely the responsibility of the authors and does not necessarily represent the official views of the funders.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The research reported in this publication was supported by the National Cancer Institute of the National Institutes of Health under award number R01CA196796 and by the Department of Medical Engineering.