
Abstract
The ability to generate distinct spatiotemporal patterns of cytosolic calcium concentration (Ca2+cyt) is a fundamental property of eukaryotic cells and underlies responses to external stimuli as well as directing downstream processes involved in morphogenesis, growth, and even developmental fate. In this review, we consider a number of well-studied and less well-studied photosynthetic plant and algal systems from the point of view of the different Ca2+ channel types that underlie spatiotemporal Ca2+cyt patterns. These include pollen tubes, root hairs, moss protonema, algal rhizoids, and single-celled algae. We show that similar spatial and temporal Ca2+cyt patterns can be brought about by the coordinated activities of a range of Ca2+ channel types. Most significantly, these channel types vary widely between different photosynthetic groups, indicating that the conserved necessity to generate spatiotemporal Ca2+ signals is satisfied by divergent underlying mechanisms, likely reflecting the different evolutionary pressures on ion transport mechanisms across the photosynthetic eukaryote clades.
Introduction
The existence of localized Ca2+ fluxes across the plasma membrane in polarized plant or algal cells was postulated as early as 1974 by Jaffe et al. 1 who determined that positive current entry into the germinating rhizoid of a brown algal zygote or the tip of a pollen tube was carried at least in part by Ca2+ ions. Although it was initially considered that net positive current entry into the growing apex of a polarized cell could provide an electrical driving force for polarization, views have since evolved toward more specific roles for localized cytosolic calcium concentration (Ca2+cyt) elevations in directing or regulating site-specific cell growth through the formation of functional ion gradients. Calcium ions play fundamental roles in a wide array of growth, morphogenetic, motility, and metabolic processes in plants, linking bioelectrical events with metabolic and molecular controls. 2
Regulation of Ca2+cyt results from the coordinated activities of channel-mediated influx of Ca2+ into the cytosol from extracellular and/or intracellular sources along with counter-transport of Ca2+ through energized pumps and transporters on plasma membrane and organellar membranes. This review focusses on ion channels that give rise to Ca2+ elevations localized to particular regions of the cytosol, paying particular attention to how these may have changed during evolution within the green lineage (Viridiplantae) and how they differ in other photosynthetic lineages that have been studied so far. We explore how convergent evolution has given rise to similar physiological and developmental properties through the recruitment of different Ca2+ toolkits.
Substantial gains and losses of Ca2+ channels have occurred during the evolution of Viridiplantae from single-celled chlorophyte algae to multicellular Embryophytes.3–5 Most significantly, the loss of canonical animal-like voltage-dependent cation channels (VDCCs), inositol trisphosphate receptors (InsP3Rs), and transient receptor potential (TRP) channels and expansions of the families of glutamate receptor channels (GLRs) and cyclic nucleotide gated channels (CNGCs) are key features of the evolutionary pathway to multicellular vascular plants. However, these substantial gains and losses are less apparent when considering the evolution of other multicellular photosynthetic eukaryotes, such as the brown algae (Phaeophyta). 3 This raises key questions around common mechanisms for generation of spatiotemporal Ca2+cyt signals in different cell types in vascular plants and parallels in other photosynthetic lineages.
Common Mechanisms and Subtle Differences in Ca2+ Channels Giving Rise to Localized Ca2+cyt Elevations in Polarized Plant Cells
A number of model cellular systems, including stomatal guard cells, root hairs, and pollen tubes, have allowed more detailed dissection of the Ca2+ signaling machinery from channel-mediated initiation of Ca2+ signals to decoding and physiological, growth and developmental responses in vascular plants. Pollen tube and root hair growth occurs in a highly polarized “tip growth” manner (anisotropic), rather than through more uniform elongation typical of many other plant cell types.
The presence of tip-focused gradients of Ca2+cyt in root hairs and pollen tubes (e.g., Refs.6–9 ) (Fig. 1) as well as spatial inhomogeneities in guard cell Ca2+cyt 10 demonstrates that plant cells can generate spatiotemporal patterns of Ca2+ in the absence of canonical animal-like Ca2+ signaling components such as InsP3Rs, ryanodine receptors, and 4-domain VDCCs. The roles of CNGCs, GLRs, and mechanosensitive OSCA (Reduced hyperosmolality–induced Ca2+ increase) Ca2+ channels are well described in regulation of stomatal aperture (see Refs. 10 and 11 ). Indeed, although the stomatal guard cell remains arguably the best-studied cellular model for plant Ca2+ signaling the significance of spatial patterns of Ca2+cyt elevations in guard cells is unclear.
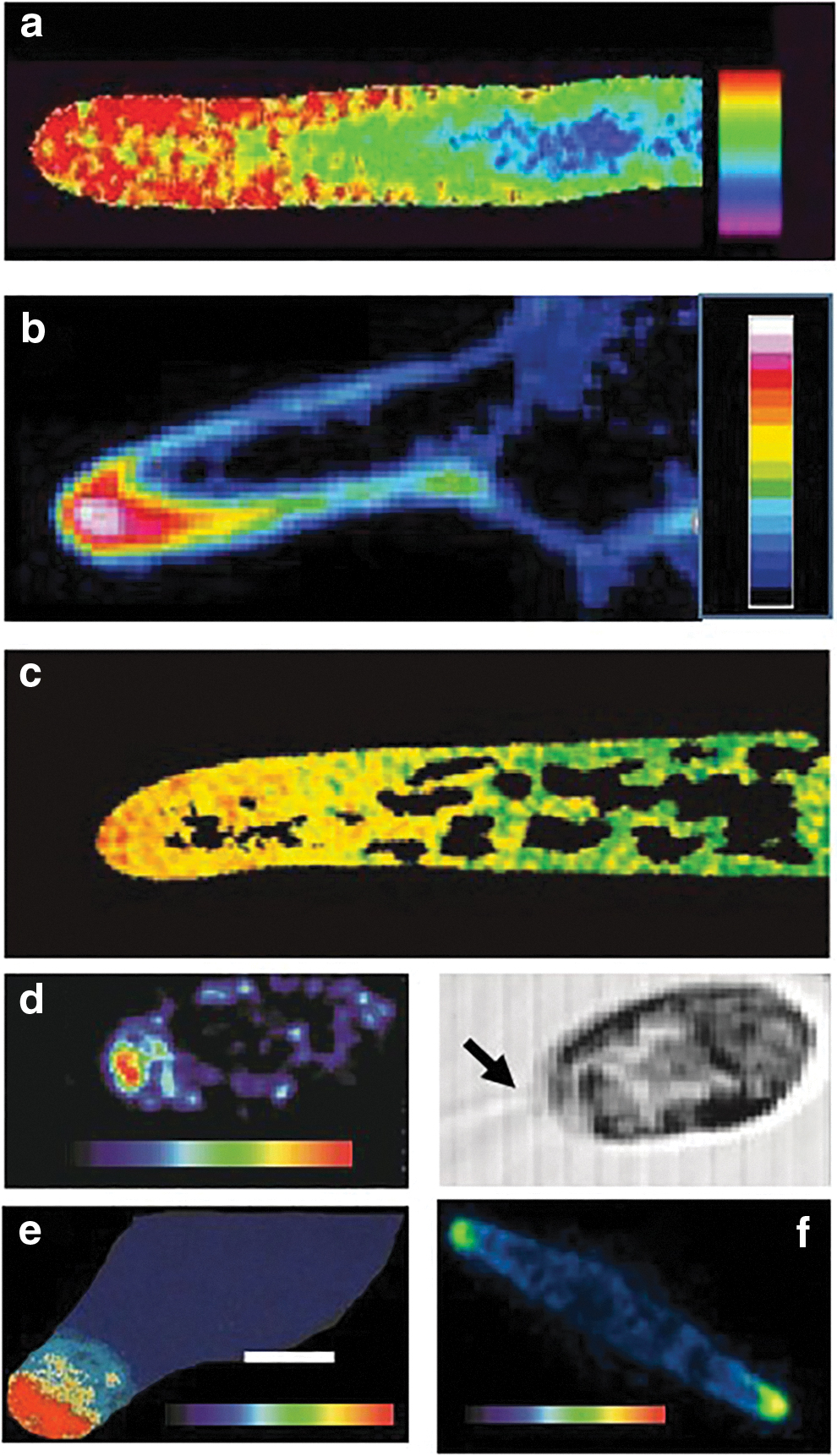
Examples of visualized Ca2+cyt elevations in apical regions of
The roles of specific ion channels in the generation of localized Ca2+ signals is becoming clearer in a number of different plant and algal systems, most notably growing pollen tubes and root hairs. In growing pollen tubes both CNGCs and GLRs contribute to the initiation and maintenance of a tip-focused Ca2+cyt gradient that may be either oscillatory or stable during growth.2,7,8,12 The Ca2+-permeable CNGC18 is preferentially localized at the growing Arabidopsis pollen tube tip and likely forms an inactive heterotetramer with CNGC8, which becomes active through interaction with calmodulin (CaM2) at low Ca2+cyt, leading to increased Ca2+ influx through the CNGC complex.12–15 In the presence of Ca2+ CaM2 dissociates from the CNGC18/8 heterotetramer, closing the channel. 14
GLRs (AtGLR1.2, AtGLR3.7, and ATGLR3.316–18) also contribute significantly to shaping of the pollen tube Ca2+cyt gradient.7,10,12 It is also proposed that GLRs function in signaling between the female synergid tissue and the growing pollen tube by acting as a ligand for d-serine produced by the ovule tissue. 16 Further regulation of GLR activity and localization has been shown to be brought about by CORNICHON homologues (AtCNIHs) that may both enable correct localization as well as channel activation.17,18 A role for mechanosensitive channels in pollen germination and growth has also been demonstrated. A homologue of a bacterial MscS channel (MSL8) is required for pollen germination and growth18,19 although to date the mode of action in relation to tip growth and influence on Ca2+cyt has not been demonstrated.
Regulation of root hair Ca2+cyt appears to involve complex interactions between a range of different plasma membrane ion channel types. A signaling hub of CNGCs (CNGC5, CNGC6, CNGC9, and CNGC14) play the prominent role in the control of localized apical Ca2+cyt in the root hair apex. In particular CNGC14, in turn regulated by CaM7 plays a specific role in both Ca2+cyt and cell integrity and growth.12,20–22 However, CNGCs are not the only players in root hair polarized growth and Ca2+cyt control. Electrophysiological studies have implicated roles for nonselective cation channels (NSCCs), 23 hyperpolarization-activated calcium channels,24,25 depolarization-activated calcium channels, 25 reactive oxygen species (ROS), NADPH oxidase (RBOHC) and Annexin 1 in modulating root hair Ca2+cyt levels and regulating growth.23,26,27 Given the enrichment of Annexin 1 in root hairs 28 the role of annexins in maintenance of the apical Ca2+cyt gradient warrants further investigation.
As well as normal root hair growth, Ca2+ signaling underlies responses to external biotic stimuli. Challenge of root hairs with pathogen-associated molecular patterns (PAMPs), such as flagellin (flg22) elicits a rapid dose-dependent depolarization of the plasma membrane potential. 29 Although flg22-induced increases in Ca2+cyt have not been reported in root hairs, Ca2+cyt oscillations at the single cell level that are mediated by plasma membrane CNGC2 and CNGC4 (death no defense: DND1 and DND2) have been visualized in other cell types in response to flg22 or chitin. 29 This Ca2+ signaling is necessary for effector-triggered immunity.30,31
Activation of these channels in Arabidopsis is brought about through the PAMP receptor brassinosteroid insensitive 1-associated kinase 1-flagellin sensing (BAK1-FLS2) receptor complex, which phosphorylates the brassinosteoid induced kinase BIK1, which in turn activates the CNGC2-CNGC4 heteromeric channel.12,31 BIK1 also plays a role in activation of RBOHD and ROS production, which may further activate CNGC2/CNGC4 through the ROS sensor hydrogen peroxide induced Ca2+ increases (HPCA1). 32 OSCA1.3/1.7 is also phosphorylated by BIK1 in response to flg22 and was also shown to be involved in the flgg22-induced Ca2+cyt increase in guard cells. 11 Although the signaling pathways are not yet defined, there is also evidence that GLRs may be involved in responses to a range of PAMPs.12,33
Distinct from plasma membrane channel-mediated increases in Ca2+cyt, mycorrhizal (Myc) factors from arbuscular mycorrhizal fungi and nodulation (Nod) factors from rhizobia can give rise to Ca2+ elevations localized to the nucleus. 34 In the legumes Medicago trunculata or Lotus japonicas, Nod factor perception by LysM-type plasma membrane receptors is conveyed to the nuclear envelope through a cytosolic mevalonate pathway. 35 Interaction between CNGC15 and the CASTOR/POLLUX/DMI1 channels is required for the release of Ca2+ into the nucleoplasm.36,37 Whereas DMI1 has been proposed to behave as a K+ channel, more recent evidence suggests that both CNGC15 and DMI1 are Ca2+ channels.37,38 Interestingly, Nod factors have also been shown to produce tip-focused Ca2+cyt elevations arising from Ca2+ influx in Medicago root hairs39,40 independent of nuclear Ca2+cyt spiking that are involved in efficiency of infection.
Physcomitrium
A variety of external cues can induce Ca2+cyt elevations in caulonema or protonema cells of the moss Physcomitrium patens.41–44 In caulonemal cells, UV light induced two distinct patterns of Ca2+cyt elevation, one that initiated from the cell apex and another that initiated at the junction of the neighboring cell. 43 In an elegant imaging study of Ca2+cyt and actin Bascom et al. 45 showed that oscillating fluctuations of Ca2+cyt regulated the assembly and disassembly of actin in the protometal apex, with direct impacts on protonemal growth. The underlying mechanisms giving rise to oscillating Ca2+cyt in the protonemal tip have not been determined.
However, both GLRs and CNGCs have been shown to underlie Ca2+cyt elevations in Physcomitrium. Application of glutamate to protonema cells produced a spatially restricted Ca2+cyt elevation as well as an electrical depolarization that propagated along the cell. 46 PpGLR1 and GLR2 are present in protonema and are responsible for the glutamate-activated Ca2+cyt increase in sperm cells. 47 Although CNGCs are also significant components of the Physcomitrium Ca2+ signaling machinery, 48 their roles in maintenance of the apical Ca2+cyt signal remain to be established. Other components of the Physcomitrium Ca2+ signaling machinery include two tonoplast-localized homologues of mechanosensitive PIEZO channels that contribute to the normal growth, size, and [Ca2+]cyt oscillations of caulonemal cells. 49
Charophyte Algae
Certain algae such as the giant celled Charophytes and filamentous Klebsormidiophytes sit on the streptophyte lineage to vascular plants. Rhizoids of Charophyte algae elongate by tip growth but despite significant physiological studies of Charophyte membrane transport (e.g., Ref. 50 ) the Ca2+ relations of growing rhizoid tips have not been explored in detail. Although the Chara genome encodes seven CNGCs and two ORAI channels that have been implicated as Ca2+ channels in nonembryophyte plants5,50 it appears to lack GLR channels.
Chara also lacks the typical animal channels represented by VDCCs, TRPs, and InsP3Rs 51 although the related alga Klebsormidium may possess an animal-like VDCC.51,52 Given the important evolutionary position of the Charophyte algae, more intensive efforts at elucidating the general and specific features of Ca2+ signaling are warranted, particularly considering the substantial gene losses that have occurred after the split of the charophytes and land plants, or after the divergence of the chlorophyta (green algae) and streptophytes (including Chara).
Chlamydomonas
Imaging studies have revealed that the single-celled green alga Chlamydomonas is able to generate distinct spatiotemporal elevations of Ca2+cyt in response to different external stimuli. Single transient Ca2+cyt elevations in response to NaCl initiate in the apical region with propagation of Ca2+ to the rest of the cell depending on stimulus strength, 53 consistent with the possession of an InsP3R and the possible operation of Ca2+-induced Ca2+ release (CICR) from intracellular stores. In contrast, hypo-osmotic treatments give rise to repetitive spatially uniform elevations in the cell body as well as the flagella, although flagella Ca2+ elevations are not in sync with Ca2+cyt, so do represent spatially distinct signals. 53
While complex interactions between different cellular compartments likely play important roles in shaping Chlamydomonas Ca2+cyt signatures, 54 a number of plasma membrane channel types have been identified with roles in spatiotemporal patterning of Ca2+. Strikingly, although Chlamydomonas does possess GNGCs and GLRs (although these gene families have not undergone the expansion seen in vascular plants and mosses), the channels identified so far that give rise to spatiotemporal Ca2+cyt signatures in green algae are quite distinct from their counterparts in vascular plants and mosses.
Plasma membrane localized Channelrhodopsins (CHR1, CHR2) are light-activated cation channels.55,56 ChR1 is permeable to other cations H+, Na+, Ca2+, and K+, but its permeability to H+ is greater than other ions. At physiological conditions, it is thought that H+ are the major conducting ions. Chr2 is a relatively NSCC. ChR activation by blue-green light gives rise to rapid localized plasma membrane depolarization and rapid cation influx in the eyespot region likely leading to localized Ca2+cyt elevations in the cytosolic eyespot region.54–56
ChRs are involved in two distinct motile responses to light, photophobic and phototaxis. ChR1 likely mediates the photophobic response, demonstrated by a brief period of backward swimming after exposure to high light. ChR-mediated membrane depolarization in response to high light activates a flagellar membrane localized VDCC (CAV2), 57 triggering an inward flagellar Ca2+ current that underpins the change in flagellar waveform during the photophobic response. Chlamydomonas also shows both positive and negative phototaxis depending on the light intensity.
Phototactic orientation is proposed to be due to the differential sensitivities of the cis and trans-flagella to flagella Ca2+. 58 Both ChR1and ChR2 are involved in regulation of phototaxis.56,59 Significantly, ChR1 shows Ca2+-dependent photophosphorylation that determines the phototactic direction through regulation of flagellar beat pattern. 59 Since their initial discovery in Chlamydomonas, a diverse range of cation and anion conducting channelrhodopsins have now been identified in many algal lineages, including cryptophytes, haptophytes, and dinoflagellates. 60 Channelrhodopsins have also been identified in some basal streptophytes (Mesostigma, Klebsormidium, and Coleochaete), but are absent from land plants. 60
A number of stimuli such as acidification, photoshock and high external Ca2+ lead to the deflagellation response in Chlamydomonas whereby cells excise their flagella. Ca2+ imaging experiments demonstrated that this was associated with localized rapid (within 1–2 s) elevation of Ca2+cyt in the apical region of the cell. 61 The deflagellation response to low pH was absent in mutants of a TRP channel ADF1 (TRP15), which is proposed to reside at the flagellar transition zone. 62
More recently, Wu et al. 63 showed that CaM, enriched in the apical cell body, interacts with ADF1 and is essential for the acid-induced deflagellation response. Interestingly TRP channels have recently been shown to underlie the response of Chlamydomonas to pathogenic bacteria. 64 Bacterially derived orfamide A and its derivatives cause cytosolic Ca2+ elevations through activation of four TRP channels (TRP15, TRP11, TRP5, and TRP22), which can lead to deflagellation and subsequent immobilization of Chlamydomonas cells through a signaling pathway distinct from the effect of low pH.
A number of other ion channels are strongly implicated in the generation of localized Ca2+ signals in Chlamydomonas. Flagellar-localized Ca2+ elevations have been visualized and shown to be responsible for regulation of retrograde intraflagellar transport.65,66 In addition to CAV2, several TRP channels and an InsP3R are likely localized on the flagellar membrane.54,67 Indeed, at least 15 further Ca2+ channel types have been identified in Chlamydomonas 54 with, as yet no clearly described function in the generation of complex spatiotemporal Ca2+ signaling.
Stramenopile Algae
The fucoid algae belonging to the Phaeophyta evolved multicellularity completely independently from the plant lineage (Streptophyta). 68 The polarizing Fucus zygote and the growing Fucus rhizoid have provided much important physiological information on the role of localized Ca2+ elevations in both symmetry breaking during early development and tip growth of the rhizoid cell. Ca2+ entry at the rhizoid pole of the zygote gives rise to locally elevated Ca2+ and is one of the earliest indicators of de novo cell polarity that underlies differential cell fate after asymmetric first division of the zygote.69–71 In growing rhizoids, patch clamp studies have shown the existence of mechanosensitive Ca2+-permeable channels that are preferentially activated at the growing rhizoid tip and correspond to a dynamic localized region of cytosolic Ca2+ in that region.72,73
There are also roles for ROS and InsP3-dependent propagated release of Ca2+ from endoplasmic reticulum stores in regulating Ca2+cyt at the rhizoid apex during growth and in response to external stimuli such as osmotic treatments 74 (Fig. 2). Indeed, the genome of the related brown alga Ectocarpus encodes an animal-like InsP3R 3 . The Ectocarpus genome also encodes three GLRs but remarkably no CNGCs, presenting a completely different set of potential players in the toolkit for generation of localized Ca2+ elevations. Strikingly, the Ectocarpus genome encodes 18 TRP-like channels and 4 animal-like VDCC channels. 3 Their roles as molecular counterparts of the channels that have been characterized electrophysiologically remains to be established.
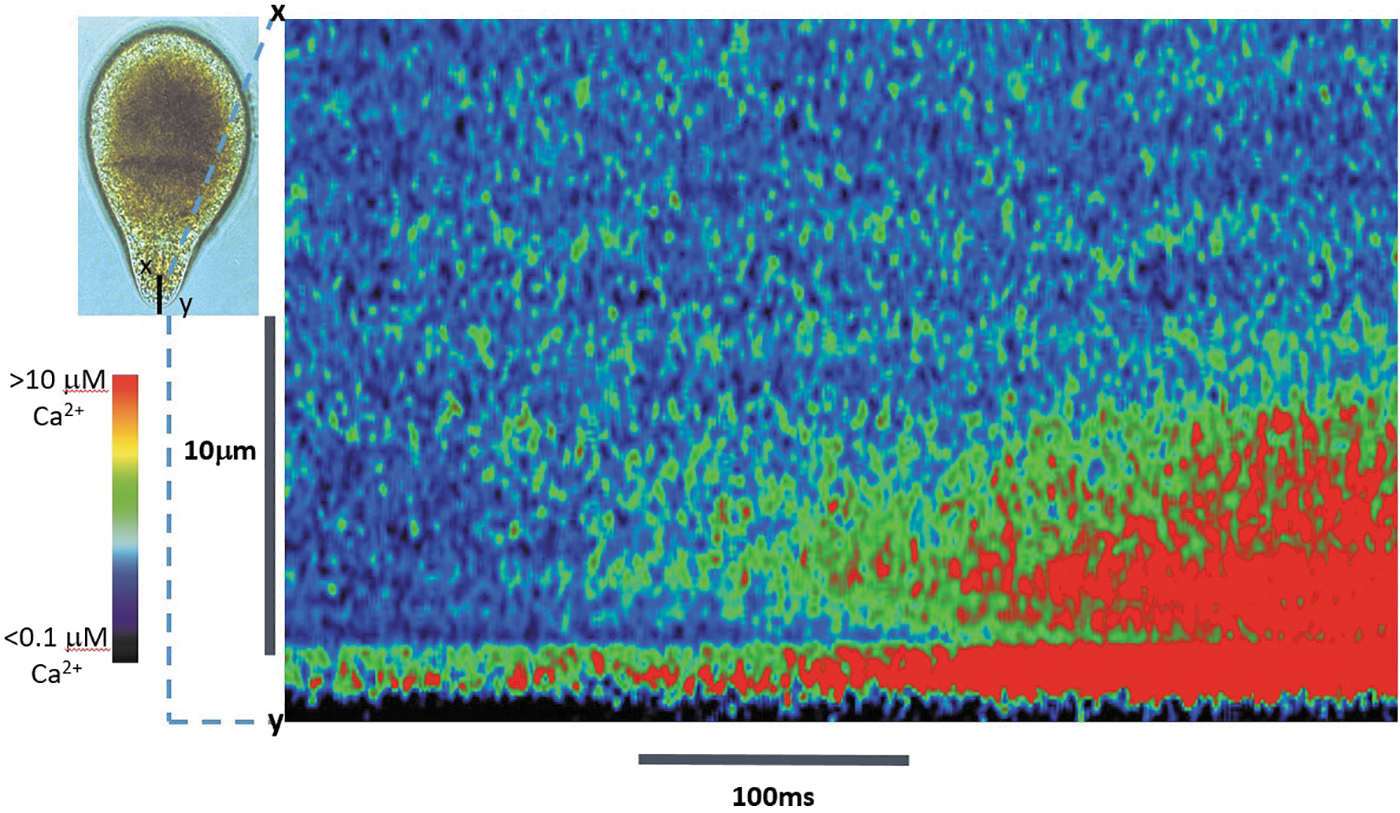
Ca2+cyt elevations at the tip of a growing Fucus rhizoid cell in response to hypo-osmotic treatment. Red regions represent high Ca2+cyt (>1 μM), blue represents resting Ca2+cyt (ca 100 nM). The kymograph visualizes Ca2+cyt elevations along confocal line scans across transect x–y (left inset) for a 1 s period. Increasing frequency of discrete Ca2+cyt elevations lasting 1–10 ms localized within 1–2 μm of the plasma membrane is followed by increased frequency of discrete highly localized Ca2+cyt elevations in the subapical ER-rich region. These propagate as a wave of elevated Ca2+cyt in both apical and subapical directions, indicating that both Ca2+ influx through plasma membrane channels and release of Ca2+ from intracellular stores are important for determining Ca2+cyt dynamics at the rhizoid tip. Color scale bar represents [Ca2+cyt] ranging from low (<1 μM, blue) to high (>1.0 μM, red). Adapted with permission from Ref. 71 ER, endoplasmic reticulum.
Fucoid algae are members of the major eukaryote Stramenopile lineage, which also includes a range of unicellular protists, including the single-celled diatoms. The diatom Phaeodactylum tricornutum has been shown to generate sophisticated spatiotemporal patterns of Ca2+cyt elevation in response to a range of different stimuli, including hypo-osmotic shock, cold shock, and nutrient (P) availability.75–77 In response to cold or osmotic shock Ca2+cyt elevations initiate at one or both cell apices, where they either remain localized or propagate as transcellular waves of elevated Ca2+ throughout the rest of the cell.75,76 Phaeodactylum lacks VDCC, GLR, and CNGC channels but does possess one TPC, four TRP channels and seven members of the MscS mechanosensitive channel group as well as three members of a novel class of eukaryotic single pore domain bacterial-like channels (EUKCATA1–3)3,78 that were recently shown to behave as voltage-gated Ca2+ channels when expressed in HEK cells. 78
The roles of these channels in generation of spatiotemporal Ca2+cyt signals remains to be elucidated. Crispr/Cas9 knockout mutants of EUKCATA1 retain the ability to generate Ca2+ signals in response to cold, osmotic, and nutrient cues but are unable to generate Ca2+cyt elevations in response to plasma membrane depolarization and are defective in Ca2+-dependent gliding motility. 78 With the advent of routine genetic manipulation and the increasing availability of genomic information, Phaeodactylum presents an ideal model to explore the molecular nature of spatiotemporal Ca2+ signals in an organism that lacks many of the channel components of the signaling toolkit that are present in green algae and plants.
Conclusions
CNGCs and GLRs play predominant roles in vascular plant and moss Ca2+ signaling, including increasingly well-characterized localized signaling pathways involved in polarity. However, a different picture emerges when considering Chlamydomonas, which possesses additional sensory and signaling components, including ChRs along with more animal-typical Ca2+-permeable channels in the plasma membrane and flagella (Fig. 3). Although certain of these are related to flagellar function and motility, it is also clear that localized Ca2+ elevations in response to external stimuli with signaling functions in the cell body are brought about by channel types that are not typical of vascular plants or mosses.
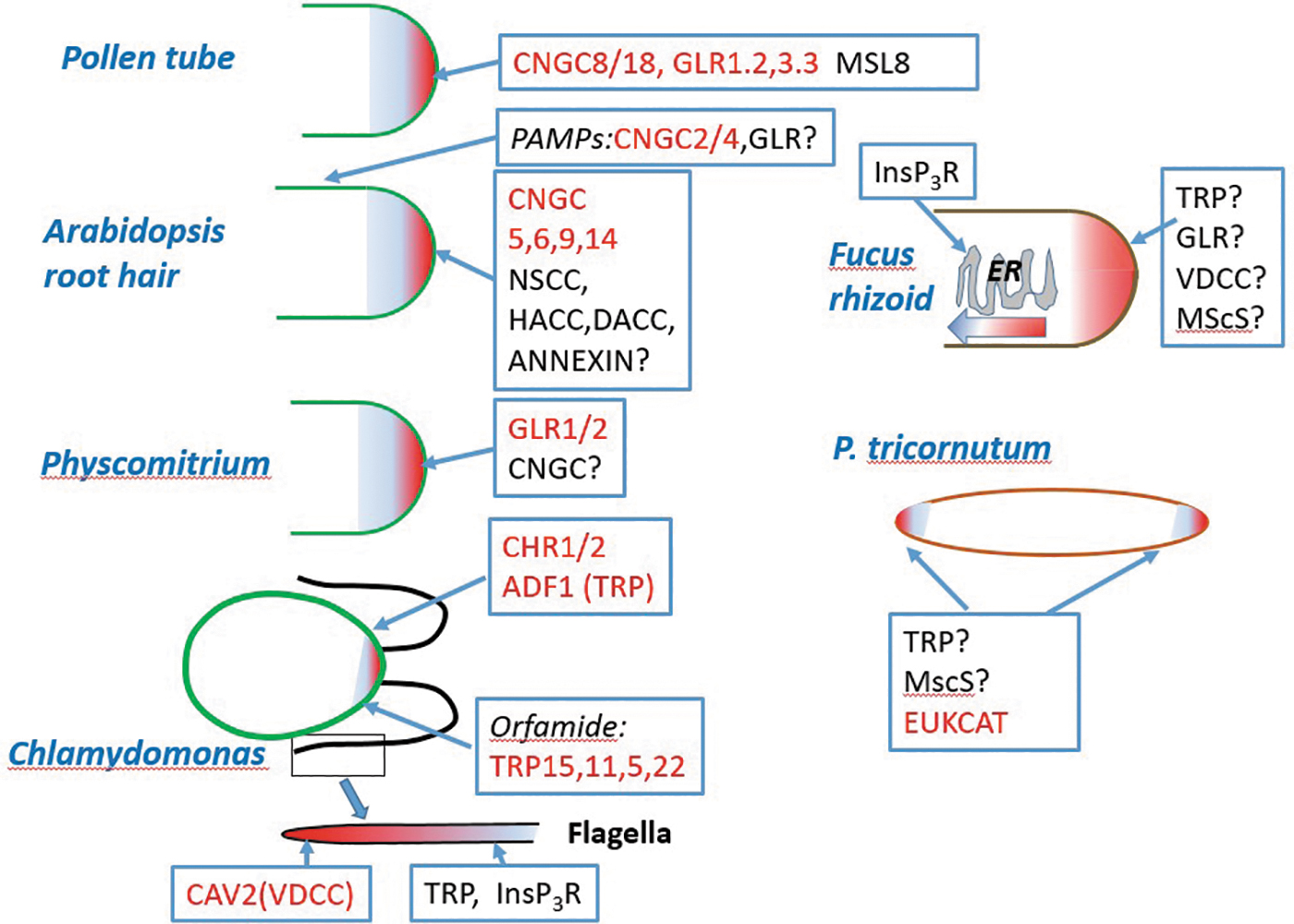
Schematic summary of the predominant Ca2+-permeable plasma channels underlying localized Ca2+cyt dynamics in pollen tubes, root hairs, moss protonema, Chlamydomonas cell body and flagella, Fucus rhizoid, and cell apices of the diatom Phaeodactylum. Channels with molecular identities involved in Ca2+cyt dynamics are shown red.
In the Phaeoophyta, a photosynthetic lineage that evolved multicellularity independently, tip-localized Ca2+ elevations similar in nature to those occurring in pollen tubes and root hairs differences are also apparent. However, the mechanisms underlying their initiation and propagation appear to be strikingly different between these major groups, based on physiological and genomic information currently available (Fig. 3). Ascribing identified channels to the Ca2+ signaling physiology in brown algae remains to be done but should provide fresh insights into how similar Ca2+ signals may be generated by the operation of different ion channel types.
Yet further divergence in spatiotemporal Ca2+ signaling mechanisms is apparent when considering other members of the Stramenopile lineage, such as the diatoms (Fig. 3). While the ion channels underlying spatial Ca2+ signaling in this group are yet to be identified, the absence of GLRs and CNGS at least in the pennate diatom Phaeodactylum and the presence of bacteria-like single pore domain voltage-sensitive Ca2+-permeable channels (EUKCATS) that are restricted to certain unicellular protists add further potential for divergence in Ca2+ signaling mechanisms in photosynthetic eukaryotes.
The ability to generate spatiotemporal patterns of Ca2+cyt is a fundamental property of photosynthetic eukaryotes. A number of evolutionary drivers have resulted in substantial divergence in the molecular players, as exemplified in this comparative survey of Ca2+ channels. However, these different complements of the Ca2+ signaling toolkit produce often strikingly similar patterns of Ca2+ associated with processes such as growth and response to external abiotic and biotic stimuli, further emphasizing the fundamental importance of being able to generate sophisticated precisely regulated spatiotemporal Ca2+ signals for cellular function.
Footnotes
Authors' Contributions
C.B. conceived the article. C.B. and G.L.W. cowrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study is supported by a Leverhulme Trust Emeritus Fellowship (EM-2021-067) (C.B.), an ERC Advanced Grant (ERC-ADG-670390) (C.B.), and a Natural Environment Research Council grant (NE/T000848/1) (G.L.W.).