
Abstract
Developmental morphogenesis reliably builds species-specific large-scale anatomies. Crucially, regulative development and regeneration after diverse and unpredictable damage illustrate that groups of cells deploy context-sensitive, homeostatic algorithms that can navigate anatomical state space in flexible ways. While the molecular machinery necessary for this process is increasingly becoming characterized, the algorithms that guide growth and form are still poorly understood. To drive progress in regenerative medicine and bioengineering, constructive models need to be formulated and tested that show the kinds of information exchange sufficient for specific morphogenetic competencies. Here, we propose a computational model of planarian regeneration that relies on cell-cell communication via gap junction channels and the use of stress as a driver of homeostatic change. We simulate key experiments in the planarian model system and show that this framework is sufficient to demonstrate the observed regenerative behavior of planaria under bioelectric perturbation. This model suggests testable hypotheses in vivo, and it provides a framework for design strategies for control circuits in synthetic bioengineering.
Introduction
Embryogenesis, regeneration, and remodeling of metamorphosis are remarkable processes in which large numbers of cells create and repair a species-specific target morphology.1,2 Understanding this process is crucial not only for basic evolutionary developmental biology, but also for the discovery of efficient interventions in regenerative medicine.
The ability to specify what shape a collective of cells will build is at the heart of problems such as birth defects, regeneration from traumatic injury and degenerative disease, cancer, and aging. Thus, it is imperative to gain a quantitative understanding of the dynamics that allow individual cells to cooperate toward a reliable large-scale outcome.
One of the most important and least understood processes is the cessation of growth when the correct pattern is complete. For example, animals such as axolotls can restore perfect copies of amputated limbs, tails, eyes, jaws, and other organs 3 : rapid growth and remodeling ceases precisely when the correct anatomical structure is completed.
Morphogenesis is not simply a hardwired set of steps but is context-sensitive and able to handle unexpected situations: regardless of where a limb is amputated, salamanders can regenerate exactly what is missing, no more and no less. Not only can this process restore structures from standard configurations (e.g., a normal limb amputated at the elbow), but it can also achieve the desired goal from evolutionarily unexpected starting configurations.
For example, tadpoles with scrambled positions of craniofacial organs still make normal frogs, 4 because the eyes, nostrils, and mouth move around in a novel path from abnormal initial positions to reach a correct frog anatomy. This capacity has been cast as a kind of anatomical homeostasis, progressively reducing the error between the current state and a specific anatomical setpoint (the target morphology).1,5
How do cellular collectives navigate morphospace 6 to reach the correct anatomies despite perturbations? A number of things are known about this process. First, the necessary endogenous communication between cells occurs via chemical, biomechanical, and bioelectrical signals.7,8 Second, the genome does not provide an actual blueprint of the anatomy, but instead specifies the micro-level hardware (proteins) of each cell; it is the developmental physiology that processes the information required to implement morphogenesis.9–11
Third, the setpoint toward which cells build is specified, in part, by bioelectrical prepatterns—spatiotemporal distributions of resting potential, 12 established by ion channels and shaped by electrical synapses known as gap junctions. Indeed, specific manipulation of ion channel and pump function has been shown to induce ectopic organs, 13 duplicate the primary body axis,14,15 repair defects of the brain, 16 trigger regeneration of appendages under non-regenerative conditions, 17 prevent oncogene-driven tumorigenesis,18,19 and induce morphologies appropriate to other species without genomic editing or transgenes. 20
While the molecular components necessary for development and regeneration are increasingly characterized,21–25 many deep problems (such as the expected morphology of chimeric organisms) remain, 26 because we do not yet understand the algorithms that are sufficient for cells to collaborate toward complex anatomical outcomes. It is necessary to design and test models of minimal circuits that would recapitulate system-level morphogenetic repair processes. Here, we specifically focus on bioelectric components of such circuits, in planarian flatworms—a model system well known for its remarkable morphogenetic control 27 and its reliance on biophysical mechanisms.28–30
In planaria, not only is the number and location of heads specified by a gradient of resting potential, 31 but also once reset to a two-headed pattern, fragments continue to regenerate as two-headed with no further manipulation, despite the wild-type genome.32,33 The ability to stably re-specify the encoding of information guiding cellular morphogenesis suggests that powerful interventions may be identified that induce desired repair and remodeling in biomedical contexts by manipulating bioelectric signaling (akin to the neural decoding and optogenetic actuation in neuroscience34–37 ).
All of these approaches, as well as the understanding of the evolution and self-repair of morphologies, require a better understanding of emergent minimal bioelectric dynamics sufficient for growth control. Thus, here we computationally model bioelectric communication between cells to uncover dynamics sufficient for the establishment of complex morphologies. Such models have been analyzed earlier, but they did not include detailed spatial analyses of gap junctional paths or a stress (error) parameter.38–40 In our model, cells use a combination of morphogen signaling and the propagation of stress—an explicit parameter reflecting the distance between the current shape and the encoded target morphology.
We show that this is sufficient to recapitulate two basic results from the classic planarian experimental corpus, 41 including a single cut, and amputation in the presence of a gap junction inhibitor. Our model is a possible springboard for understanding the anatomical homeostasis of cellular collectives and for designing novel bioengineering constructs such as self-repairing living machines.
Models and Methods
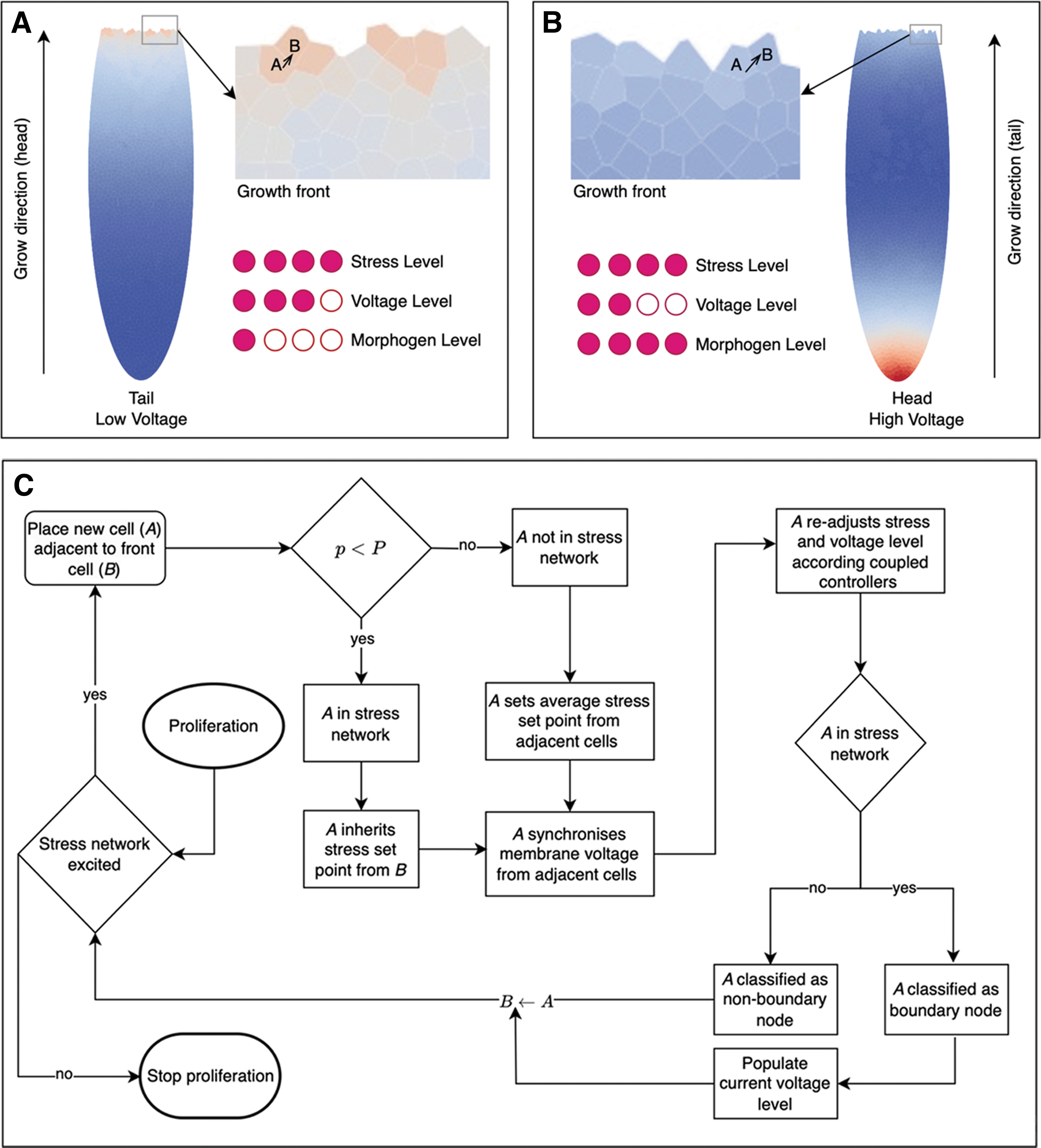
We are interested in how long-range communication paths control and guide the collective regeneration processes in (morphological) space and time. It is now known that gap junctions and ion channels establish compartment boundaries for organs,42–50 and that alterations of bioelectric properties influence the shaping of organs such as the brain, as well as control of the transition from normal tissue to tumorigenesis, at long range. 51 Thus, we built a model to better understand the control of morphogenesis during regeneration by bioelectric dynamics propagating through tissues. An overview of our model approach is depicted in Figure 1.

Model overview:
We describe the stress network generation in Figure 2 as follows:

Stress network generation.
Coupled stress-voltage controllers
We consider two coupled proportional integral controllers for stress and membrane voltage (Fig. 1B). The control action from one controller influences the other and vice versa through a constant factor, α.
The discrete errors are given by:
The discrete control action for the stress controller without coupling is given by:
We introduce the coupling between the two as follows.

We arrive at first-order difference equations of the following kind:

In this scenario, the two controllers are additively coupled. Other scenarios are possible. However, we show that with this simple coupling function we get a rich set of complex state space trajectories.
In the continuous limit, we can derive a coupled system of integro-differential equations:
For a systematic derivation, see Appendix A.
We list all variables and descriptions in the table given next:
Stress network based on Potts model
The Hamiltonian for the q-state Potts model on a graph
The partition function Z is given by
For a directed graph
Potential propagation via gap junctions
We use the concept of node potentials in a connected network and the Laplacian to propagate changes in membrane potentials throughout the network. We consider a connected graph
Let
We classify the nodes of G into two categories: boundary nodes (set B) and non-boundary nodes (set N). Boundary nodes are nodes in the stress network governing the propagation of changing membrane potentials, whereas non-boundary nodes do not participate in the stress network. The Laplacian matrix L can be partitioned based on these categories as:
where
The intracellular dynamics between stress and membrane voltage controller drive changes in membrane potentials resulting in a potential difference
From the second set of equations, we can write:
Results
Toy model reveals the stabilizing effect of noise
We first describe an experiment with a toy network to assess the impact of varying levels of stress and voltage coupling among cells on system stability in planarian regeneration. Initially, we generate simulations based on a simplified network of three nodes, representing cells, and we intentionally exclude the role of voltage propagation via gap junctions.
We investigate two core scenarios: one with constant, stable connections between cells, and the other with one connection randomly eliminated at each time step, introducing instability. Further, we segment the investigation into three unique conditions based on different stress and voltage coupling levels, aiming at understanding how these variations affect system disturbances and stability. The broader purpose of this initial analysis is to enhance our comprehension of the intricate cellular mechanisms involved in regeneration.
As discussed in the Models and Methods section, our study is primarily built on dual attributes inherent to each cell: a stress controller and a voltage controller (Fig. 1). A fundamental assumption in our work is that these two mechanisms are internally coupled within each cell, with cellular interactions mediated through the influence of stress among neighboring cells.
We hypothesize that a wide spectrum of stress influences from various cells augments the overall stability of the system, in comparison to a system where stress influence originates from a fewer number of cells. To illustrate this, we use a simple network composed of just three nodes, representing three cells. Our focus is on the dynamics of stress coupling and stress-voltage coupling, excluding voltage propagation via gap junctions for simplicity. This represents the first quantitative model (Fig. 3) implementing the qualitative idea proposed recently, 57 that stress sharing is a mechanism for coordinating cell behavior into larger-scale anatomical homeostasis.
Growth and proliferation processes.
We delve into two specific cases. The first scenario involves stable stress connections between cells (represented by the edges in our network) that persist throughout the simulation (see left column in Fig. 4). This implies that all connections between cells are constantly maintained. In contrast, the second scenario introduces a degree of instability, with one connection being randomly eliminated at each time step during the simulation (right column, Fig. 4).

Toy network with three nodes. Nodes represent cells, and edges represent intercellular stress coupling between cells. Main columns. Right column: fully connected cell network during simulations. Left column: unstable connections during simulation (we randomly drop one connection each time step). Number of time steps: 100. α denotes the coupling strength between stress and voltage controller. λ denotes the stress coupling between cells. Sub-columns in each panel: stress and voltage levels as a function of time. Coordinates of the state-space; x = stress level, y = voltage level. In all configurations: voltage and stress setpoints drawn from a normal distribution
Our research is further diversified into three distinct scenarios based on varying levels of stress and voltage coupling: Scenario A (Fig. 4A), where both stress and voltage couplings are sufficiently high (α = 0.3 and λ = 0.3); Scenario B (Fig. 4B), where stress coupling is high and voltage coupling is low (α = 0.05 and λ = 0.5); and Scenario C (Fig. 4C), where stress coupling is low and voltage coupling is high (α = 0.5 and λ = 0.05). In all scenarios, unstable connections resulted in higher system disturbances.
Our results point to a counterintuitive role of noise in the system: it appears to enhance the overall system stability within the cellular dynamics of planarian regeneration. We interpret this by considering that a higher volume of incoming signals—due to a larger number of coupled cells—increases the probability of certain signals exhibiting destructive interference, which, in turn, stabilizes the system.
Model scenario 1: bisecting a wild-type planarian
We next conducted an experiment focusing on a bisected wildtype planarian. This is a more difficult case than the two-cuts scenario modeled in many planarian papers, because in the case of two cuts it is easy to visualize a morphogen gradient that sets the pattern of the fragment. However, in the case of one cut, the cells on either side of the cut were direct neighbors (in effect, at the exact same positional information level58,59) and yet have directly opposite anatomical fates (head and tail respectively). This shows that the wound identity cannot be set locally and must arise as the result of long-range communication 5 ; those are the dynamics being modeled here.
We created simulations under defined parameters to observe the distinct growth paths, namely tail-to-head and head-to-tail. Our intent was to discern the impact of intracellular stress mechanisms and cell network voltage propagation on membrane potential dynamics during the various phases of planarian regeneration in our model.
The analysis of a wildtype planarian, cut down the middle, is illustrated in Figure 5A, where diverse colors indicate varying membrane potentials in arbitrary units. We investigated two distinct growth trajectories—tail-to-head (top row) and head-to-tail (bottom row), guided by the model outlined in Figure 3. Our simulations considered specific coupling constants for both voltage and stress controllers
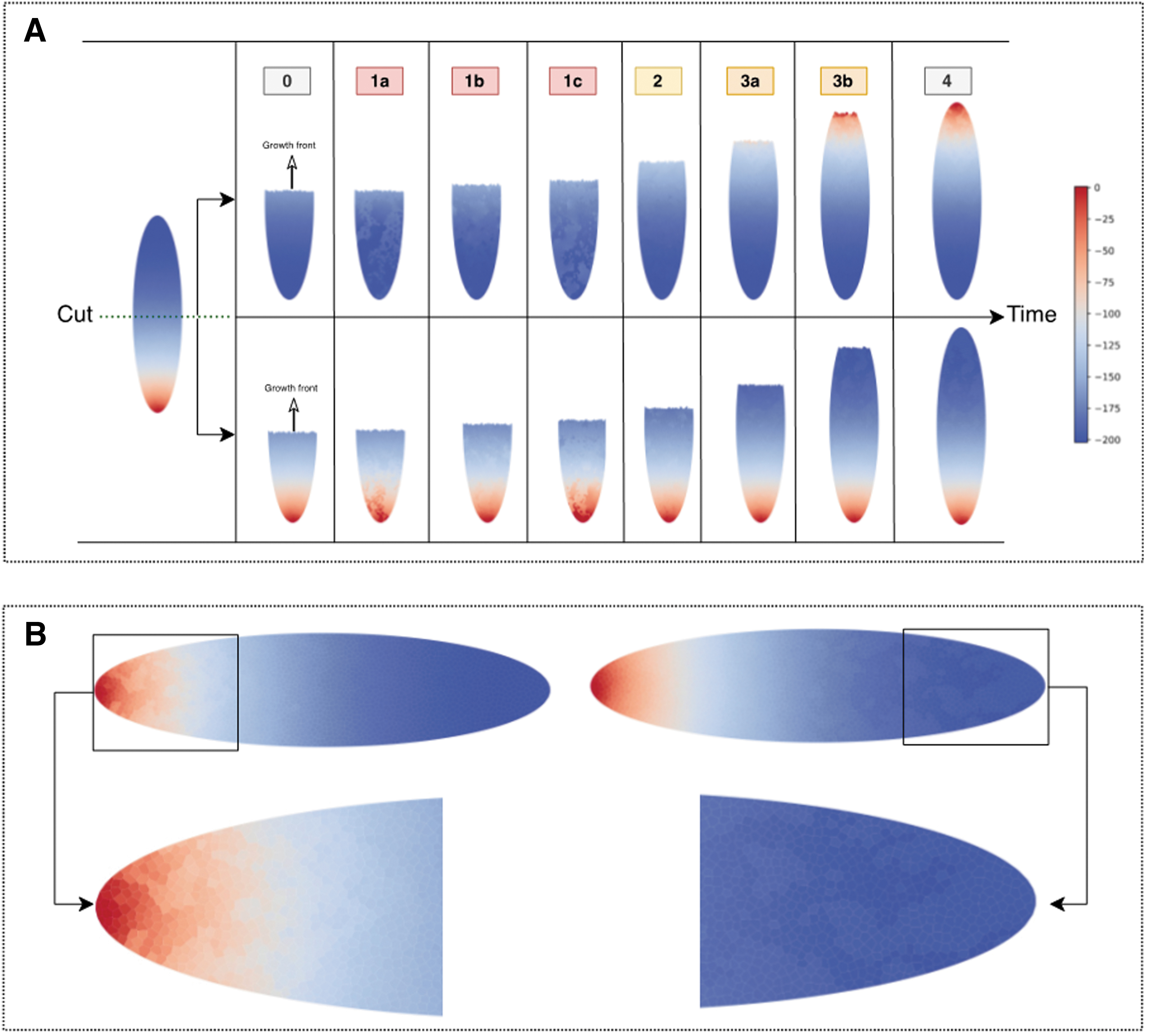
Model predictions in the case of a single bisecting cut.
The experimentation was structured to traverse three significant stages of regeneration, all discernible through alterations in membrane potential dynamics influenced by intracellular stress mechanisms and voltage propagation through the cell networks.
The results generated from this experiment were multifaceted. Initially, we observed vigorous oscillations in membrane potentials during phases 1a to 1c, indicative of robust synchronization incited by intracellular stress voltage dynamics. This high-activity phase transitioned into a brief relaxation phase, Phase 2, where oscillatory activity was significantly diminished in areas where tissue proliferation was complete.
The final phase, Phase 3, was characterized by increased synchronization activity along the stress networks, particularly evident toward the latter part of the regeneration process. Interestingly, despite the intense activity, the regeneration process did not perfectly replicate the original planarian, as shown by the “inherited” membrane voltage profile of the regenerated parts, particularly observable in Phase 4.
This inheritance of stress network structure manifested differently in regenerated heads and tails. The average membrane potential in regenerated heads was 5% to 10% higher, whereas regenerated tails exhibited an increased average potential of 2% to 7%, across 10 independent simulations.
Wildtype planarian, cut in the middle, with partial and full gap junction blockage
One of the key observations about long-range control in planaria is the finding that blocking gap junctions after amputation induces permanent changes in the target morphology of planaria.33,62 In this section of our study, we aimed at elucidating the role of gap junctions, crucial elements facilitating intercellular communication and voltage synchronization, in planarian regeneration.
Specifically, we investigated the effects of partial and total blockage of these junctions on the regeneration process and the resulting membrane voltage structures. To accomplish this, we employed computational simulations with conditions emulating varying degrees of gap junction blockage, and we studied the subsequent regeneration under these modified conditions.
We performed an experiment where we modified conditions to emulate blockage of gap junctions, a crucial element in cellular communication and synchronization. As illustrated in Figure 6, we began with a wildtype planarian that was cut in the middle. We then observed the resulting growth under two distinct conditions: partial gap junction blockage and total gap junction blockage.

Wildtype planarian grows into two-headed planarian after exposure to gap junctional blocker. Blockage of gap junctions (such as by octanol33,62) has been simulated by randomly setting off edges between adjacent cells. We set off two to three connections per cell per timestep.
The parameters for this experiment were kept consistent with those used in Figure 5. Under partial blockage, our intention was to observe the effects of incomplete voltage synchronization between neighboring cells. With total blockage, we aimed at investigating the growth patterns when the gap junctions were entirely nonfunctional.
Under partial blockage, we noted irregular voltage membrane structures, particularly near the head region of the regenerating planarian. We attribute this to the higher stress levels found in the newly proliferated cells, which caused disturbances in the membrane voltage levels. Interestingly, these disturbances were sustainable and caused deviations in the stress setpoint of the new cells, and due to the partial loss of gap-junction based voltage synchronization, these disturbances were persistent.
On the other hand, when the gap junctions were completely blocked, the regenerated planarian still developed two heads, but the voltage disturbances near the head region were significantly reduced compared with those observed under partial blockage. Irrespective of the degree of blockage, the early stages of regeneration (phases 1a–1c) exhibited similar dynamics to the experiment without gap junction blockage.
However, from Phase 2 onward, we noted increasing disturbances due to the lack of voltage synchronization during regeneration. Remarkably, these observed patterns proved to be stable under both conditions.
Discussion
Many prior efforts have sought to model signaling relevant to planarian regeneration.38–40,63–69 Our model adds to this a detailed simulation of gap junctions (bioelectric synapses) and a stress parameter to make explicit the mechanistic dynamics of a homeostatic loop that progressively seeks to reduce error. 2 Future work will expand the model itself, and apply it to the full range of known planarian functional experiments, to refine it and generate testable predictions.
Additional experiments that need to be simulated include the persistence of the two-head state in subsequent cuts in the absence of octanol,33,62 the effects of pharmacologically induced voltage change on the number of heads,14,31 the appearance of a destabilized (randomly choosing one-head vs. two-head in fragments) phenotype,32,61 and the effects of anxiolytic drugs such as serotonin transporter inhibitors.70–72
Subsequently, the model can be expanded to include known molecular pathways involved in planarian regeneration downstream of the voltage signals21,73–77 and other cell behaviors such as proliferation and migration.78–80
In both our simplified network model and the gap-junction blockage experiment, we discovered that disruption in the connections within the system—either by randomly eliminating connections in the model or by blocking gap junctions in the experiment—resulted in increased system instability. The resilience we initially observed, as well as the subsequent sustainable deviations in membrane voltage distributions, strongly underscores the critical role of these connections in maintaining system stability.
In the case of the gap junction blockage experiment, we noted persistent and sustainable deviations in the membrane voltage levels of the regenerated planarian compared with the normal voltage distribution in the head region of wildtype planarians. This outcome corroborates our findings from the simplified network model where a lack of stable connections resulted in increased disturbances within the system.
Taken together, these results suggest a hypothesis that can be tested in planarian experiments in vivo: stability is highly dependent on the integrity of connections, whether they be stress influences among cells or gap junctions facilitating voltage propagation.
Additional experiments suggested by this model include degrees of reduction of stress; targeting stress perception could be accomplished by means of anxiolytic drugs,81–85 enabling the planarian model system to shed light on the most interesting and still largely mysterious aspect of morphogenesis: the ability of the system to detect deviations from target morphology and execute self-limiting growth and repair activity.
One limitation of the planarian model system is that it is currently not possible to directly visualize either gap junctional states or stress molecules non-destructively. Being able to visualize this in the living state while planaria regenerate is a key goal for the development of non-invasive optical monitoring techniques, and it will greatly enhance the study of how this versatile, robust model species reliably produces whole bodies from tissue fragments.
In addition to planaria, this model will be adapted to understand size control of organs (e.g., liver) or embryonic/regenerating structures in other model systems, including mammals. In turn, cycles of modeling and empirical testing will reveal sufficient mechanisms by which groups of cells coordinate and make organ-level anatomical decisions.
This knowledge is essential not only for basic evolutionary developmental biology (origin of specific forms and evolvability of repair mechanisms 86 ) but also for attempts to exert rational control over large-scale growth and form in biomedical and synthetic bioengineering settings.87–91
Footnotes
Acknowledgments
We thank Julia Poirier for her help with the manuscript.
Author Disclosure Statement
M.L. is a cofounder of, and the Levin lab receives funding from, a company called Morphoceuticals, which seeks to induce biomedical regeneration by modulating bioelectrical signaling.
Funding Information
M.L. gratefully acknowledges support via Grant 62212 from the John Templeton Foundation (the opinions expressed in this publication are those of the author(s) and do not necessarily reflect the views of the John Templeton Foundation) and the Guy Foundation Family Trust (103733-00001).