
Abstract
Limited information is available concerning changes that occur in the brain early in human immunodeficiency virus (HIV) infection. This investigation evaluated resting-state functional connectivity, which is based on correlations of spontaneous blood oxygen level-dependent functional magnetic resonance imaging (fMRI) oscillations between brain regions, in 15 subjects within the first year of HIV infection and in 15 age-matched controls. Resting-state fMRI data for each session were concatenated in time across subjects to create a single 4D dataset and decomposed into 36 independent component analysis (ICA) using Multivariate Exploratory Linear Optimized Decomposition into Independent Components. ICA components were back-reconstructed for each subject's 4D data to estimate subject-specific spatial maps using the dual-regression technique. Comparison of spatial maps between HIV and controls revealed significant differences in the lateral occipital cortex (LOC) network. Reduced coactivation in left inferior parietal cortex within the LOC network was identified in the HIV subjects. Connectivity strength within this region correlated with performance on tasks involving visual-motor coordination (Grooved Pegboard and Rey Figure Copy) in the HIV group. The findings indicate prominent changes in resting-state functional connectivity of visual networks early in HIV infection. This network may sustain injury in association with the intense viremia and brain viral invasion before immune defenses can contain viral replication. Resting-state functional connectivity may have utility as a noninvasive neuroimaging biomarker for central nervous system impairment in early HIV infection.
Introduction
I
The onset of these changes is not clear. Despite the extensive neuropathology seen at autopsy, there is an assumption that the brain is spared injury in early stages of infection. The flu-like symptoms of initial viral infection, termed acute HIV, soon resolve and are followed by an asymptomatic period that may extend over a decade in some individuals. However, an intense viremia occurs in association with acute HIV and the initial virus host interaction. In the period of initial infection, before the host has mounted an immune response that can control viral replication, viral particles in plasma expand rampantly, rising to an average peak of 10 million copies/mL (Goldman and Ausiello, 2008). During this high-grade viremia, virus is widely disseminated throughout the body (Wawer et al., 2005). The immune system is profoundly perturbed and many potentially neurotoxic factors are markedly upregulated in the attendant cytokine storm. Brain viral entry in this early period of infection is well established (Chiodi et al., 1988; Chiodi and Fenyo, 1991; Davis et al., 1992). As host immune defenses are mounted, levels of HIV RNA in plasma generally decline to a steady state that may be undetectable in many individuals for long periods (Schacker et al., 1996). Nevertheless, the virus cannot be eradicated and may persist, quietly replicating in reservoirs such as the brain, where, owing to the blood brain barrier, it remains sheltered from neutralizing antibodies or antiviral therapy. Taken together, this evidence suggests the possibility that the onset of injury underlying cognitive impairment may date to early infection.
Synaptodendritic neuronal injury may be a critical mechanism underlying HAND (Bruce-Keller et al., 2003; Moore et al., 2006; Orenstein et al., 1988) for a review, see (Ellis et al., 2007). Dendritic injury (Masliah et al., 1997) and reduced synaptic density (Everall et al., 1999) have been associated with presence and severity of HIV neurocognitive impairment. Synaptodendritic injury, involving anatomic and functional damage to pre- and postsynaptic structures, has deleterious effects on neuronal networks and on brain connectivity, disrupting the highly integrated functioning of neural systems required to process information (Ellis et al., 2007). This injury may be clinically silent, however, in early, more potentially reversible stages (Moore et al., 2011). Capabilities for in vivo analysis of resting-state functional connectivity, which is based on correlations in spontaneous blood oxygen level-dependent (BOLD) signal oscillations between brain regions, may afford critical insight into possible functional brain changes occurring early in infection and inform an understanding of the cognitive deficits that manifest later in the clinical course. Resting-state networks, which are covarying regions identified by resting-state functional connectivity analysis, show close correspondence with networks identified by explicit task-related activation (e.g., the motor network) (Biswal et al., 1995) and deactivation (e.g., the default mode network) (Raichle et al., 2001). Such evidence supports the premise that functional networks of the brain “in action” and “at rest” share the same underlying neuronal communication substrate (Smith et al., 2009). These intrinsic correlations have been used to investigate the architecture of brain systems and to identify potential impairment in functional networks in neurological and neuropsychiatric disorders (e.g., Greicius, 2008; Jang et al., 2011). Because this approach has a physiologic basis and is not dependent on symptom presentation, it may be sensitive to subtle changes occurring early in HIV infection.
This investigation tested the hypothesis that alterations in brain functional connectivity can be detected by resting-state functional magnetic resonance imaging (fMRI) early in HIV infection. Resting-state networks were evaluated in participants of the Chicago early HIV infection study. This cohort includes recently infected subjects, defined as within ∼1 year of initial HIV-1 exposure, and age-matched seronegative controls (Ragin et al., 2011). To evaluate cognitive function in early infection and to examine patterns of relationship with resting-state connectivity, subjects were also evaluated with a neuropsychological test battery that has been used extensively in studies of HIV neurological outcome (for example, Schouten et al., 2011; Sevigny et al., 2007).
Methods
Subjects
Participants included 15 HIV and 15 age-matched, seronegative control subjects from the Chicago early HIV infection cohort study. Of the subjects, 13 HIV and 15 controls were right handed. Study exclusion criteria included history of neurological disorder, stroke, head trauma, opportunistic central nervous system (CNS) infection, psychosis, or MR contraindication. Informed consent approved by the Northwestern University Institutional Review Board was obtained from each subject prior to the experiment.
For enrollment of HIV subjects, the likelihood of recent infection was assessed based on clinical history criteria, including either a recent prior negative test result or compelling information indicating probable time of initial viral exposure. Blood samples were collected from all subjects. Seropositivity was assessed in all study participants by enzyme-linked immunosorbent assay and western blot. To independently assess the relative recency of infection, blood samples from the HIV subjects were also analyzed using an early infection assay (Blood Systems Research Institute, San Francisco), which has been specifically designed to evaluate individuals whose antibody response against the virus is still evolving following seroconversion. Based on current assay cutoff values, the period since infection was conservatively estimated to an average of <1 year for this sample (average, 363.3 days). In the seropositive subjects, absolute CD4 cell counts ranged from 162 to 1115/mm3, with mean and standard deviation (SD) of 506.8±227.3 mm−3 and median of 474 mm−3; plasma viral load ranged from 37 (i.e., undetectable) to 346,000 copies/mL, with mean and SD of 59,581±10,780 and median of 5,550 copies/mL. Seven HIV subjects were treatment naive when evaluated. Of those receiving treatment, the majority were taking atripla, a single medication that combines efavirenz, tenofovir DF, and emtricitabine (one nonnucleoside and two nucleoside reverse transcriptase inhibitors). Further clinical details for the HIV subjects are presented in Table 1.
HIV, human immunodeficiency virus.
Neuropsychological assessments
All participants received a neuropsychological evaluation on the same day as the MR scan. The neuropsychological test battery assessed motor skills, cognitive flexibility, abstraction, verbal memory, nonverbal memory, and visuoconstructional skills. This battery has been used in clinical trials and neurological outcome studies in HIV infection (e.g., Sevigny et al., 2004). Specific cognitive tests included the Rey Auditory Verbal Learning Test (verbal memory), the Rey-Osterrieth Complex Figure Delayed Recall Test (visual memory), and the Rey-Osterrieth Complex Immediate Recall Test (constructional skills) (Rey, 1941). Psychomotor skills were assessed using the Digit Symbol Test (Wechsler, 1981) and motor skills were assessed using Grooved Pegboard (GP) (dominant and nondominant hand) (Klove, 1963) and Timed Gait tests. Reaction times were measured using the California Computerized Assessment package (Miller et al., 1991). Frontal systems were assessed with Verbal Fluency (Benton and Hamsher, 1976) and Odd-Man-Out (Flowers and Robertson, 1985) tests. General intellectual performance was assessed with the National Adult Reading test (Nelson and O'Connell, 1978; Stern et al., 1992) (see Table 2).
HIV and Control groups were compared using independent t-tests.
Spearman correlations (r) between neuropsychological performance measures and brain connectivity strength; p values have been adjusted for FDR.
p<0.05, FDR corrected.
SD, standard deviation; FDR, false discovery rate.
MR imaging protocols
Scanning was performed at the Northwestern University Center for advanced MRI using a Siemens whole-body scanner (Siemens Trio Tim B15 software, Elangen, Germany) with a 12-channel head coil. The imaging protocol included structural and functional sequences.
Structural MRI
High-resolution 3D T1-weighted MRI scans were acquired using a magnetization-prepared rapid gradient echo sequence (MPRAGE) (TR=2300 ms, TE=2.91 ms, flip angle=9°, field of view=256×256 mm, voxel dimension=1 mm isotropic, acquisition time=10 min).
fMRI (resting-state BOLD)
Whole-brain functional imaging was performed using a gradient echo EPI sequence (TR=2500 ms, TE=20 ms, field of view=220 mm, in-plane resolution=1.72×1.72 mm, imaging matrix=128×128, slice thickness=3 mm, 40 slices with no gap, GRAPPA, PAT:2, number of reference line=24, phase partial Fourier=6/8, acquisition time=588 sec; all images were acquired axially in the same axial plane defined by the anterior and posterior commissures). During the resting-state scan, subjects were instructed to relax and keep their eyes open.
Image analysis
Data analysis was carried out using FSL tools (
Brain connectivity and clinical measures
Spearman correlations were calculated between connectivity strength, determined by the average beta-values in the areas that exhibited group differences (Control > HIV) and neuropsychological test scores separately for HIV and control groups. Relationships between connectivity strength and measures of systemic disease progression (HIV viral RNA in plasma and CD4 viral load) were also examined in the HIV group. Within the regions showing between-group differences (Fig. 2C), average beta-values were extracted from the subject-specific spatial maps for the lateral occipital cortex (LOC) component from the final step of the dual regression (note that the beta maps are not normalized by the residual noise). These measures were then correlated with cognitive test results from the neuropsychological evaluation. Analyses were carried out using Matlab 7.6.0 (Mathworks, Inc.). Independent t-tests were used to compare the cognitive measures in the HIV and control groups. To adjust for multiple comparisons, all p values were false discovery rate (FDR) corrected using SAS (Cary, NC).
Results
Neuropsychological performance
As shown in Table 2, the HIV subjects demonstrated poorer performance on two neuropsychological tests, Digit Symbol (p=0.033, FDR corrected) and Letter-Number Sequencing (p=0.033, FDR corrected). Trend-level differences were identified in the HIV group for Rey Auditory Verbal Learning (Delayed Recall) (p=0.093, FDR corrected) and Rey Complex Figure Recall (Delayed Recall) (p=0.069, FDR corrected).
Resting-state functional connectivity
The major resting-state networks (executive control, default mode, sensory motor, primary visual, lateral occipital, auditory, etc.) were identified by visual comparison of the 36 components with previous reports (Beckmann et al., 2005; Damoiseaux and Greicius, 2009) and are shown in Figure 1. Gaussian mixture models were fitted to the individual z maps to infer voxel locations significantly modulated by the associated time course to allow for meaningful thresholding of the z images. The remaining 28 components have been included in Supplementary Figures S1–S28 (Supplementary Data are available online at

Eight group independent component analysis component maps from 30 subjects:
Group main effects for Control (Fig. 2A) and HIV subjects (Fig. 2B) are shown in Figure 2. Significantly decreased coactivation within the LOC was identified in the HIV compared with the Control group. This decreased area was located in the inferior parietal cortex, according to Juelich Histological Atlas (Caspers et al., 2008) (maximum Z value at X=−39, Y=−69, Z=24, 27 voxels, 216 mm3) (Fig. 2C).
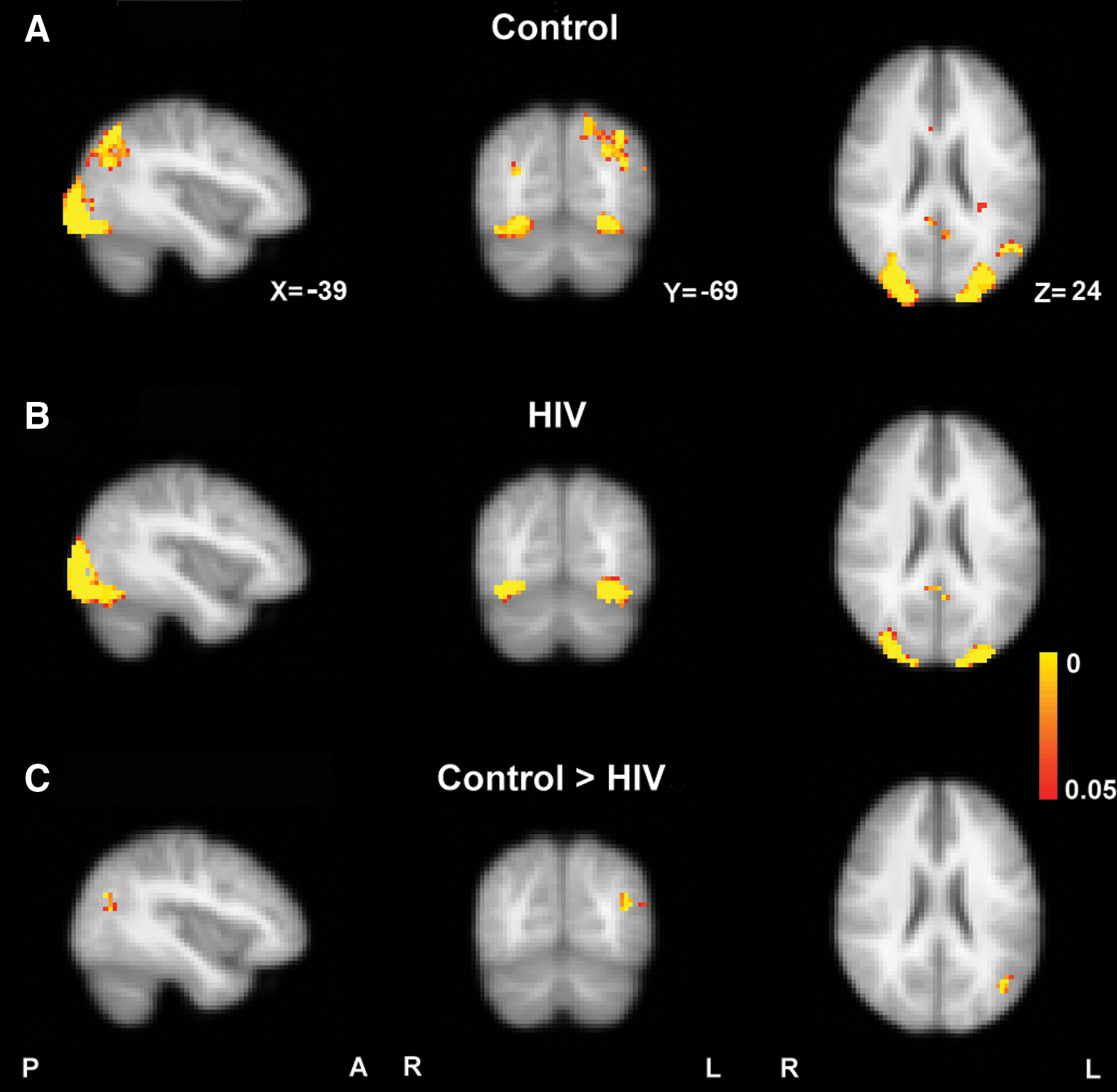
Group main effect for
Each voxel's beta-value represents the coactivation strength with the overall component time series. Accordingly, a significant group difference for a given voxel reflects focal connectivity reduction or enhancement relative to the associated overall resting-state network (Zhou et al., 2010). Note that this correlation can be either positive or negative. The average beta-value within the regions exhibiting statistical group differences (Fig. 2C) were extracted and compared in HIV and controls. As shown in Figure 3, on average, the control group demonstrated a positive correlation (average beta-value=5.08), whereas the HIV group demonstrated a negative correlation (average beta-value=−4.20).
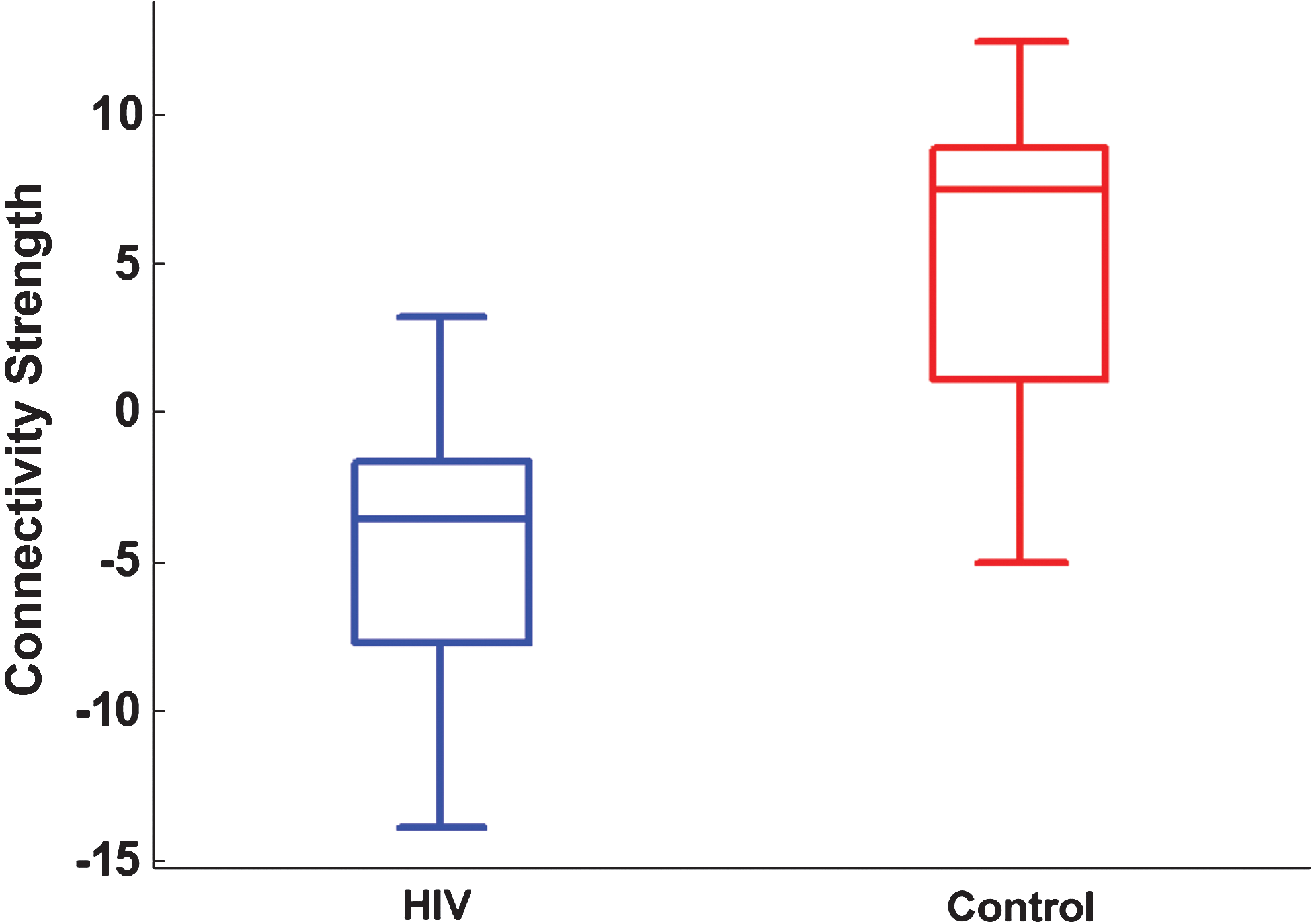
The box plot of average beta-values within regions exhibiting statistical group differences (Control > HIV, regions shown in Fig. 2C) for HIV and Control. Color images available online at
Correlations between brain connectivity and clinical measures
Statistically significant correlations were identified between connectivity strength and GP dominant scores (r=−0.775, p=0.013, FDR corrected) (Fig. 4A and Table 2) and with Rey Figure Copy performance (r=0.705, p=0.046, FDR corrected) (Fig. 4B and Table 2) in the HIV group only. There were no significant correlations for the control group (shown in Fig. 4C, D for comparison). There were no significant correlations between connectivity strength and markers of systemic disease progression: viral load in plasma and CD4.
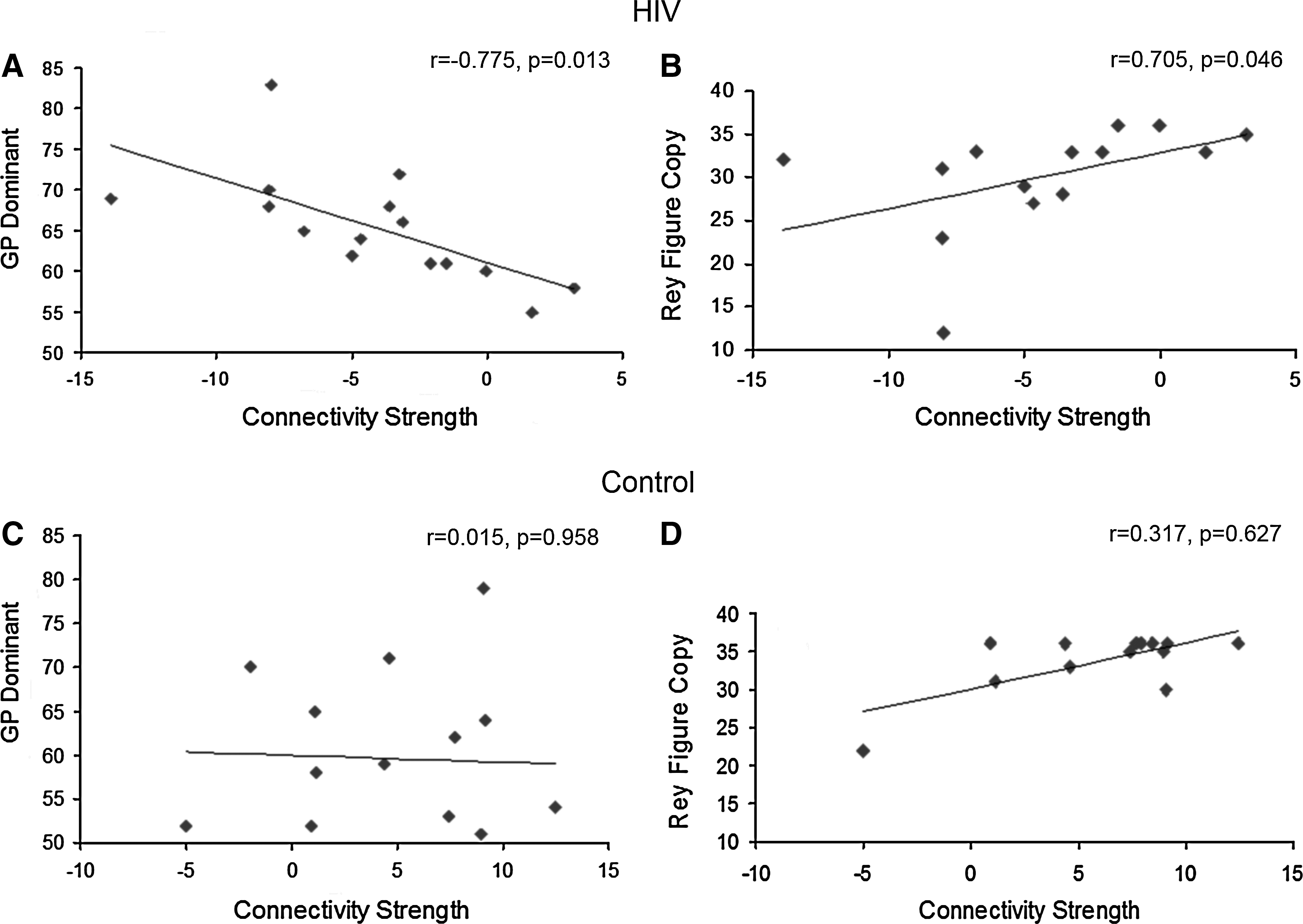
Top row: in the HIV group,
Discussion
Structure and function of various neural systems may be altered in HIV infection. On neuropsychological testing, impairment may be evident in executive function, motor skills, speed of information processing, episodic memory, attention/working memory, language, and visuoperception [for a review, see (Woods et al., 2009)]. HAND is diagnosed by impairment in two or more of the following domains: verbal, attention/memory, executive, speed of information processing, sensory perceptual, and motor skills. Severity is classified on the basis of interference with everyday function: asymptomatic (no interference with activities of daily living), mild neurocognitive disorder, and HIV-associated dementia (marked interference) (Antinori et al., 2007). Ongoing injury to the brain may be asymptomatic for prolonged periods of the clinical course before cognitive symptoms begin to manifest. A neuropsychological study of subjects in very early, acute HIV infection (n=39), in more advanced infection (n=63), and in controls found significant performance differences only for the more advanced infection group (Moore et al., 2011). In contrast, cognitive function in the acute infection group tended to be lower than controls, and the differences were not significant. An emphasis of this study is whether there are biomarkers that can detect changes occurring in the brain, early in the course of infection, and during periods that may be otherwise asymptomatic. This comprehensive analysis of resting-state networks indicates that impaired functional connectivity in the LOC network can be detected early in HIV infection. This network is defined as the component that includes the occipital pole extending laterally toward the occipito-temporal junction and more dorsally into posterior parietal regions. The region that differed in HIV subjects occupied the superior parietal lobule (7PL, posterior part of BA 7 near the parieto-occiptial sulcus laterally, one of the subdivisions of superior parietal cortex) (Scheperjans et al., 2005) and inferior parietal lobule (PGp, one of the seven subdivisions of inferior parietal cortex defined in probabilistic cytoarchitectronic maps (Caspers et al., 2008). Specifically, recently HIV infected subjects exhibited reduced coactivation of the inferior parietal cortex within the LOC network (Fig. 2C).
The LOC functional brain network is a highly robust network that has been reported by many prior investigations. Studies based on lower-order ICA decomposition have identified a single LOC network (Beckmann et al., 2005; Smith et al., 2009). Studies employing higher-order ICA decomposition (>50 components) have detected multiple LOC subnetworks, such as the V5 motion, ventrolateral visual stream, and dorsal visual stream networks (Abou-Elseoud et al., 2010; Smith et al., 2009). Of these LOC subnetworks, the dorsal visual stream serves two distinct functions: (1) spatial awareness, involving construction of a detailed map of the visual field and (2) guidance of action through detection and analysis of motor actions. The dorsal visual stream commences with purely visual function in the occipital lobe before gradually serving spatial awareness functions at its termination in the parietal lobe. The posterior parietal cortex is essential for visuo-motor integration, that is, integration of motor movements with visual perception (Zhang et al., 2008). Lesions in the posterior parietal cortex result in deficits in reaching and fine-motor coordination with visual guidance (Nelson and Luciana, 2008). A study in which primary resting-state functional networks were mapped onto “behavioral domains” (experimental paradigm classifications) in the BrainMap database, the LOC network corresponded to “cognition space” (Smith et al., 2009). The parietal cortex is also part of the lateralized “frontal-parietal” networks (Fig. 1G/H), which have been implicated in several cognition/language paradigms. It is worth noting that functionally distinct systems may overlap anatomically, that is, a particular brain area may participate in multiple distinct functional networks, and thus is not surprising to observe overlapping ICA spatial maps, where a particular region of the brain is involved in different resting-state networks (Beckmann et al., 2005).
Functional connectivity between different brain networks has been studied intensively using multimodal brain imaging techniques, for example, fMRI, positron emission tomography (PET) (Friston et al., 1993), and electroencephalography (EEG)/magnetoencephalography (MEG) (Wang et al., 2007, 2008; Rajagovindan and Ding, 2008). Resting-state functional connectivity is an indirect measure that reflects anatomic connectivity and synaptic efficiency (the ability of a presynaptic neuron to activate a postsynaptic neuron) (Buckner, 2010). Possible persistent sources contributing to resting-state functional connectivity include monosynaptic and polysynaptic anatomic connectivity, anatomic connectivity to a common source, and differences in synaptic efficiency across anatomically connected systems. Confounding sources include gross anatomic variability and atrophy (Buckner, 2010). Diminished connectivity between posterior parietal cortex and the visual network could reflect impairment in anatomic connectivity or reduction in synaptic efficiency across the posterior parietal cortex and the extrastriate visual cortex, resulting in impaired integration of visual perceptual/visual spatial information required for visuo-motor integration.
Relationships between functional brain connectivity and cognitive measures were identified only in the HIV subjects. Specifically, significant correlations were indicated between functional connectivity and GP (dominant) (r=−0.775, p=0.013) and with Rey Figure Copy (r=0.705, p=0.046). GP (dominant hand) is a timed test of complex, visual-motor coordination, where longer time required indicates poorer performance. Rey–Osterrieth Complex Figure Copy assesses visuo-construction and visuo-motor integration (perceptual organization) (Spreen and Strauss, 1998) (lower scores here indicate poor performance). The directionality of the observed correlations with functional connectivity (negative for GP and positive for Rey-Figure Copy) suggest that strong negative functional connectivity corresponds to impaired visuo-motor performance. Significant correlations were not indicated in the control group, suggesting that the pattern observed for the HIV subjects is unlikely to be explained by individual differences in task performance. Based on the observed group differences in functional connectivity and brain-behavior correlations, it may be surmised that circuit alterations in the LOC network, indexed by changes in functional connectivity measures, may contribute to impaired cognitive function in HIV subjects. The groups did not differ in performance on these cognitive tests. This may be because of the small sample size. The findings also suggest the possibility that resting-state analysis may afford a preview of dysfunction that is not evident on the basis of impairment in neuropsychological performance measures and thus functional connectivity changes may represent a sensitive biomarker of changes in the brain.
The results reported here together with other neuroimaging findings may provide the basis for an imaging biomarker in the clinical setting of HIV infection. A study of recently infected HIV subjects using arterial spin labeling MRI found significant reductions in resting-state cerebral blood flow (rCBF) in both visual cortex and lenticular nuclei (Ances et al., 2009). This finding suggests the possibility of a selective pattern of neuronal or vascular injury in these regions. Further, rCBF was reduced in neuropsychologically unimpaired HIV individuals, suggesting that detectable rCBF changes occurred prior to neuropsychological impairment. Reduced functional connectivity observed in recently infected subjects may reflect early changes in gray matter. Anatomic imaging studies of subjects infected for longer periods indicate that brain atrophy is most severe (15%–20% loss) in parietal and frontal lobes (primary and association sensorimotor cortex) and the extent of gray matter loss in these areas correlates with severity of cognitive impairment (Chiang et al., 2007; Thompson et al., 2005). Changes in white matter have also been shown in subjects infected for longer periods (Chen et al., 2009; Ragin et al., 2004; Stebbins et al., 2007; Tate et al., 2010). Toward integrating structure and function, Tate et al. showed that GP dominant scores were correlated with number of white matter “streamtubes” (a quantitative measure of white matter integrity) in HIV subjects (Tate et al., 2010). In light of evidence that BOLD signal correlations between regions are mediated at least in part by direct and indirect anatomic projections (Damoiseaux and Greicius, 2009), functional connectivity changes observed in this study may indicate structural and anatomic connectivity changes, such as neuronal death and synaptodendritic injury (Lindl et al., 2010), early in HIV infection.
This in vivo imaging investigation indicates altered functional connectivity in LOC cortex network early in HIV infection. Changes observed in this network may reflect injury due to the dramatic viremia and brain viral invasion associated with initial exposure to the virus before host cellular immune defenses can contain viral replication. Approximately half of the HIV subjects had commenced treatment. The possibility of treatment neurotoxicity has been suggested by findings indicating improved cognitive function in HIV patients who discontinued medication (Robertson et al., 2011). The sample size precludes formal analysis of treatment on the LOC network changes observed here. Further studies with larger samples are needed to determine effects of treatment, including possible neurotoxic effects, on the LOC network.
Conclusion
In summary, this investigation of resting-state functional connectivity identified differences between HIV and controls in the LOC network. No differences were indicated for other resting-state networks, such as default mode, executive control, motor, primary visual, and auditory. Alterations in LOC network functional connectivity may represent a putative causal factor for impaired performance on neuropsychological tests requiring visuo-motor coordination. Evidence of resting-state changes within the first year of initial HIV infection demonstrates the promising utility of this approach to yield new insights concerning cognitive deterioration and neurological progression. These results need to be replicated in larger, longitudinal studies to determine whether there are changes in brain functional connectivity with disease progression and to evaluate the potential of LOC network functional connectivity changes as a prospective, early biomarker of CNS injury in HIV infection.
Footnotes
Acknowledgments
The authors thank Hongyan Du from Northshore University Health System, Evanston, IL, for her assistance in statistical analysis and Drs. Christen F. Beckmann and Xinian Zuo for their helpful discussion regarding dual regression technique. This research was supported by the National Institute of Mental Health (grant no. MH080636).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.