
Abstract
Overt picture naming causes activation in both left and right hemispheres as demonstrated by functional magnetic resonance imaging (fMRI) studies. Unexpectedly, a previous experiment revealed that there was no difference in brain lateralization related to handedness. To understand the mechanisms behind this phenomenon and to investigate the influence that each hemisphere exerts over the other in more detail, we analyzed the effective connectivity in the naming network. More specifically, we examined the impact of handedness on hemispheric interactions. Four brain regions activated for correct naming in the previous fMRI experiment served as volumes of interest: Bilateral fusiform gyrus (FG) related to visual-semantic processing and bilateral Brodmann's area (BA) 44 related to phonological processing. We defined three dynamic causal models (DCMs) representing different symmetrical connectivity patterns, compared them using Bayesian model selection, and considered differential connection strengths. For the majority of 31 participants, the same model featuring early integration of visual-semantic processing and independent phonological processing was optimal. This also held true if right-handers, left-handers, and ambidexters were considered as independent groups. For left-handers, the connection of right FG and right BA 44 was significantly stronger compared with the other two groups, and it was significantly stronger than its left-hemisphere equivalent. In right-handers, the according left-hemisphere connection was significant, but connectivities were rather symmetrical. These findings reveal significant and unique right-hemisphere contributions to word production.
Introduction
H
The lateralization of language functions as revealed by Wada tests is correlated to handedness. The left hemisphere is language dominant in >95% of right-handers, but only in 70% of left-handers (Springer and Deutsch, 2001). For the remaining cases of left-handers, either the right hemisphere is dominant or both hemispheres are equally engaged. Using functional transcranial Doppler sonography, it was found that the right hemisphere was language dominant in 7.5% of their right-handed participants (Knecht et al., 2000a) and that the incidence of right-hemisphere dominance increased linearly with the degree of left-handedness (Knecht et al., 2000b).
Various brain networks related to language tasks have been investigated with functional magnetic resonance imaging (fMRI), revealing that language processing recruits tissue in both the left and right brain, albeit most studies show greater leftward activation (Thompson, 2005). However, a clear functional laterality has not been observed in all language tasks and studies. A review of brain imaging studies on word production revealed reliable activations for picture naming in various regions bilaterally including lateral and medial temporo-occipital as well as inferior frontal areas (Indefrey and Levelt, 2004). Springer and Deutsch (2001) indicated that there is a general tendency of the brain to show symmetrical increases in activity even though the task may be initiated by predominantly one hemisphere. It remains largely unresolved as to whether these right hemisphere activations indicate significant and unique contributions to the task or whether they represent co-activations only, and, if the latter, what functional significance they might have.
One approach to investigating hemispheric interactions involves the analysis of functional connectivity of brain regions, for example, by means of psycho-physiological interaction (e.g., Stephan et al., 2003). A more recently developed technique is dynamic causal modeling (Friston et al., 2003). The DCM allows investigating not only functional but also effective connectivity of brain regions, that is, the influences that all activated brain regions of a pre-defined functional network exert over one another and how these are modulated by contextual influences.
The DCM has been successfully applied to a variety of language-related brain networks in healthy speakers as well as patients with aphasia (Allen et al., 2008; Bitan et al., 2005; Chow et al., 2008; Fu et al., 2006; Ghio and Tettamanti, 2008; Heim et al., 2009a, b; Leff et al., 2008; Mechelli et al., 2005; Noppeney et al., 2008; Seghier and Price, 2009; Sonty et al., 2007). In particular, hemispheric asymmetries were investigated regarding the anatomic connectivity patterns of language-related pathways (Powell et al., 2006) or the effective connectivity patterns of lateralized tasks involving not only speech-motor processing but also visual attention (e.g., Siman-Tov et al., 2007) and motor tasks (Eickhoff et al., 2008; Grefkes et al., 2008a, b, 2010; Stephan et al., 2007a, b).
The analysis of effective connectivity in the overt language production network, regarding the influence each hemisphere exerts on the other, has the potential to elucidate the underlying mechanism of left-right interactions in more detail. Moreover, examining the impact of handedness on brain connectivities could reveal novel aspects of brain lateralization. Along these lines, we considered our previous fMRI experiment with overt picture naming in 34 healthy subjects (Abel et al., 2009), which yielded wide bilateral brain activations. Figure 1 shows that correct naming elicited visual processing streams as well as sensory-motor, auditory, self-monitoring, and language areas similar to those previously described by Indefrey and Levelt (2004; see also Vigneau et al., 2006; Whatmough and Chertkow, 2003). Interestingly, activation patterns did not differ between the groups of predominantly left- versus right-handers as defined by the Edinburgh Inventory (Oldfield, 1971) and assessed by two-sample t-tests (threshold: p (uncorrected)=0.001, k ≥10 voxels).

Results from a conventional functional magnetic resonance imaging-analysis of correct picture naming versus rest for all 34 healthy subjects (red color; general linear model-analysis using SPM5, threshold: voxel p uncorrected<0.001 and cluster p uncorrected<0.05) rendered onto the left
Although no differences in laterality were found in the analysis of brain activations (General linear model [GLM] analysis), differential neural interactions underlying these activations might be observable by using DCM (see also Heim et al., 2009b; Schuyler et al., 2010). Competing network models specifying different connectivity patterns can be tested against the data and compared with each other. Resulting parameter values of each DCM comprise the driving inputs (e.g., of a picture), the strengths of endogenous (intrinsic) connections, and the contextual modulations (e.g., by the naming task).
In the current study, we examined the effective connectivity between brain regions by applying DCM to the data obtained during naming (Abel et al., 2009). More specifically, we aimed at revealing differences in the transmission and exchange of information within and between both hemispheres as a function of handedness. Two hypotheses were tested:
(1) The network architecture of both hemispheres differs depending on handedness.
(2) The strengths of connections within the same basic network differ between right-handers, left-handers, and ambidexters.
To test hypothesis (1), we constructed three dynamic causal models (DCMs) that represent three different symmetrical connectivity patterns. The selection of the DCMs was based on a neurocognitive model of language processing (Indefrey and Levelt, 2004). We compared the DCMs by using a Bayesian model selection (BMS) procedure (Penny et al., 2004; Stephan et al., 2009, 2010) and determined the optimal model for the entire group of all 31 participants and for each subject group. Moreover, we investigated possible differences in model parameters between right-handers, left-handers, and ambidexters to test hypothesis (2).
Materials and Methods
Participants, stimuli, and task
The data previously reported in Abel et al. (2009) were used for the DCM analysis. In three out of the 34 healthy subjects, activation maxima did not involve all four regions within the predefined search radius (see below). These three participants were excluded. The remaining 31 subjects had a median age of 28 years (mean 36, range 18–69).
According to the Edinburgh Inventory (Oldfield, 1971), 13 participants were right-handed (Laterality Quotient (L.Q.) ≥80; median age 28), nine were left-handed (L.Q. ≤−80; median age 28), and nine were ambidextrous (L.Q. above or below 80 and −80; median age 34). In fact, 15 participants were women (7 right- and 5 left-handed, 3 ambidextrous), and 16 were men (6 right- and 4 left-handed, 6 ambidextrous). The study was approved by the Freiburg University Ethics Committee. All participants gave their informed written consent.
The stimulus material consisted of 132 black-and-white drawings of concrete objects (Snodgrass and Vanderwart, 1980) (for linguistic parameters of the material, see Abel et al., 2009). The pictures were consecutively presented by using Presentation software version 10.0 (Neurobehavioral Systems), and subjects were to overtly name each picture as fast and accurately as possible. Each participant's vocal responses were sound-recorded, online classified, and later on inspected to assess correctness of their naming responses.
fMRI scanning and stimulus presentation
A Siemens TIM-Trio 3T scanner was used to acquire T2*-weighted echoplanar images (data matrix, 64×64; FOV, 19.2 cm; TE, 30 ms; TR, 2.19 sec; motion and distortion correction by scanner software, Zaitsev et al., 2004) as well as T1-weighted anatomical images (MPRAGE: data matrix, 256×256; TR, 2.2 sec; TE, 2.15 ms; pixel size, 1 mm3). Functional data were continuously acquired from 36 slices in an interleaved order with a thickness of 3 mm covering the cerebrum.
Using an event-related design, each picture to be named was presented for 1.2 sec, followed by a fixation cross for 4.8 sec and a blank screen of a jittered duration (mean 2 sec, range 1–3 sec), thus resulting in a trial duration of approximately 8 sec. The blank screen indicated the end of each naming trial. Sixteen null events were included in the fMRI experiment. We performed four consecutive sessions including 33 trials (140 image volumes=5 min) each. For the analysis, brain images of all four sessions were concatenated, and sessions as well as estimated realignment parameters were used as regressors of no interest.
Statistical parametric mapping
Images were analyzed with SPM5 (
Definition of DCM models
For the three DCMs, we chose brain areas that had been bilaterally activated in the previous fMRI naming experiment (Abel et al., 2009), which were related to main stages of word production in confrontation naming and which were not at the same time implicated in the understanding and monitoring of the words spoken (Indefrey and Levelt, 2004). Two left-hemisphere brain regions and their right homologs, all with activation maxima in the main contrast, were finally selected (see Fig. 1). The bilateral FG has been associated with visual-semantic processing of the picture to be named (e.g., Murtha et al., 1999). Left middle FG (localizable between y=−50 and y=−60 in Talairach space; matches our present volumes of interest (VOIs) of FG, see Table 1 for MNI coordinate) and left pars opercularis of the inferior frontal gyrus have previously been reported to be involved in word-type sensitive coupling in reading (Mechelli et al., 2005). The posterior section of Broca's region (BA 44) has shown to support consecutive phonological processing of the picture name (e.g., Heim et al., 2008). Right BA 44 has been reported to be implicated in phonology as well (e.g., Pugh et al., 1996). Thus, we selected homolog brain regions that covered early semantic (FG) and later phonological (BA 44) processing.
For each of the 34 subjects, VOIs were identified that were closest to the given coordinates and within a spherical search volume of 6 mm radius; for three participants, we did not find activations for all four VOIs at p<0.05 uncorrected.
FG, fusiform gyrus; BA 44, Brodmann area 44; R, Right; L, Left; VOIs, volumes of interest.
Figure 2 depicts an outline of the three DCMs. The number of regions, connections, and driving inputs is the same for all models, implying similar degrees of model complexity. Each DCM entails 6 endogenous connections (partly bi-directional) and two driving inputs. Moreover, each connection may be modulated by the naming task, reflecting the up- or down-regulation of a generally existing connection between two regions during the naming task. The models comprise driving inputs of the picture to both FG as well as ipsilateral connections running from FG to BA 44. The network architecture is presumed to be symmetrical for both hemispheres, accounting for the bilateral activation pattern found in the GLM analysis. Along the same lines, the two ipsilateral paths are held constant across all models, thus supplying equal basic conditions for the performance of the naming task in each hemisphere. To investigate the hemispheric interaction, the models differ with regard to (i) the processing stage at which initial information transmission to the contralateral hemisphere occurs (early visual-semantic exchange and/or later transmission of visual-semantic information to the phonological stage), and (ii) the presence of a direct informational exchange of phonological information. In model #1, there are forward connections from both FG to contralateral BA 44, and bi-directional connections between both BA 44, resulting in an inter-hemispheric information transmission not before the phonological stage and a direct exchange between both BA 44. In model #2, there are bi-directional connections between both FG accompanied by bi-directional connections between both BA 44, yielding inter-hemispheric exchange both at early visual-semantic and later phonological stages. In model #3, information transmission is again present at both stages, but with early visual-semantic exchange and later independent phonological processing at both left and right BA 44, as implemented by connections from each FG to contralateral FG and BA 44. For our model-driven DCM selection, we did not take implausible connection patterns into account, in particular not backward connections from BA 44 to FG.

Outline of the three models tested in the current study. Circles represent the brain regions involved in the networks, and arrows the uni- or bidirectional endogenous connections or the driving inputs into regions. R, right; L, left; PIC, driving input of the picture to be named.
Extraction of fMRI signals for the DCM analysis
The four brain areas with activation maxima in the main contrast of the conventional fMRI analysis served as VOIs (Fig. 1). For each subject and each VOI, the individual local maximum of the main effect (p<0.05 uncorrected) (cf. Heim et al., 2009b; Mechelli et al., 2005) was automatically determined that was closest to the group maximum (see Table 1) within a spherical search volume of 6 mm radius. We ensured that all resulting VOIs overlapped with the appropriate target region by using the WFU PickAtlas and the SPM anatomy toolbox as just described. Three subjects did not show activations reaching the criterion in all four VOIs and were, therefore, excluded from the analysis. For the remaining 31 participants, a time series was extracted as the first principal component of all voxel time series within a sphere (radius 4 mm) centered on the individual local maximum corrected for effects of interest. The average coordinates of the individual local maxima that constituted the centers of the spheres for the time series extraction were as follows (MNI space; mean x, y, z): right FG (31, −56, −16), left FG (−33, −51, −17), right BA 44 (45, 9, 27), left BA 44 (−48, 10, 24).
DCM model selection and parameter test
To compare the three models just outlined, we used a BMS procedure (Eickhoff et al., 2009; Heim et al., 2009b; Penny et al., 2004). We applied the random-effects BMS approach with posterior model probabilities (for methodology, see Stephan et al., 2009, 2010). Thereby, we determined the relative evidence for each DCM, which corresponds to the marginal likelihood p(y|m) of observing the data y in a particular model m integrated over the model parameters. The random-effects model selection procedure (RFX approach) has been included in the latest version of SPM 8.
After determining the “winning” model with BMS, the posterior estimates of its parameters were entered into one-sample t-tests (SPSS 15.0; with additional Bonferroni-correction for multiple comparisons) to investigate whether they differed significantly from zero. Afterward, parameter estimates were compared for all three subject groups (two-samples t-tests). The parameter estimates include the direct influences of driving inputs on the neuronal population activity in a given brain region, the strengths of endogenous connections reflecting the context-independent coupling between these regions, and the modulatory inputs revealing context-dependent changes in the coupling between these regions (Friston et al., 2003; for further details on the DCM analysis, cf. Heim et al., 2009b).*
Results
The same activation pattern found in the original GLM analysis of 34 healthy subjects (Abel et al., 2009) was also observable for the subset of the 31 participants considered for the present DCM comparison (see Supplementary Fig. S1 and Table S1). The main peak of brain activation for correct picture naming was in left FG. As in the previous analysis of all subjects, there were no significant differences in brain activations between the three subject groups in this subset (right-handers, left-handers, ambidexters; two-samples t-tests, threshold: voxel p uncorrected <0.001 and cluster p uncorrected <0.05). There were also no significant differences regarding gender (two-samples t-tests) or age of participants (covariate analysis) while applying this threshold.
We first tested hypothesis (1) by using the RFX BMS approach. As a result, the posterior probability of model #3 exceeded that of all other models for the entire group of subjects as well as, separately, for right-handers, left-handers, and ambidexters (see Fig. 3).
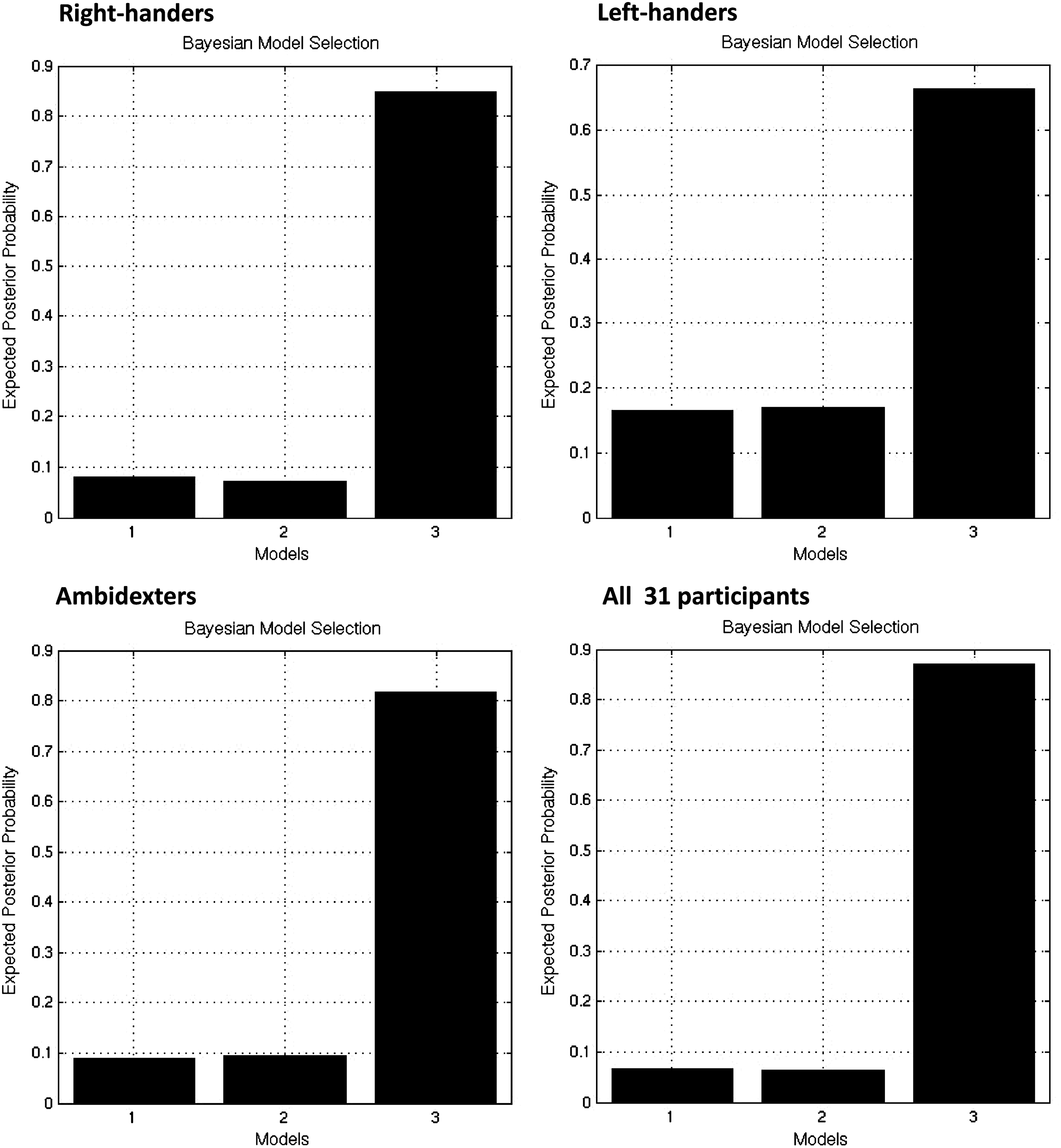
Comparison of all three models by using the random-effects model selection procedure (RFX approach).
To test hypothesis (2), we further considered the appropriate parameters (mean) of winning model #3 for each subject group and all 31 participants (see Fig. 4). In the following, we consequently present results for endogenous connections, driving inputs, and contextual modulations (see also Tables 2 and 3).
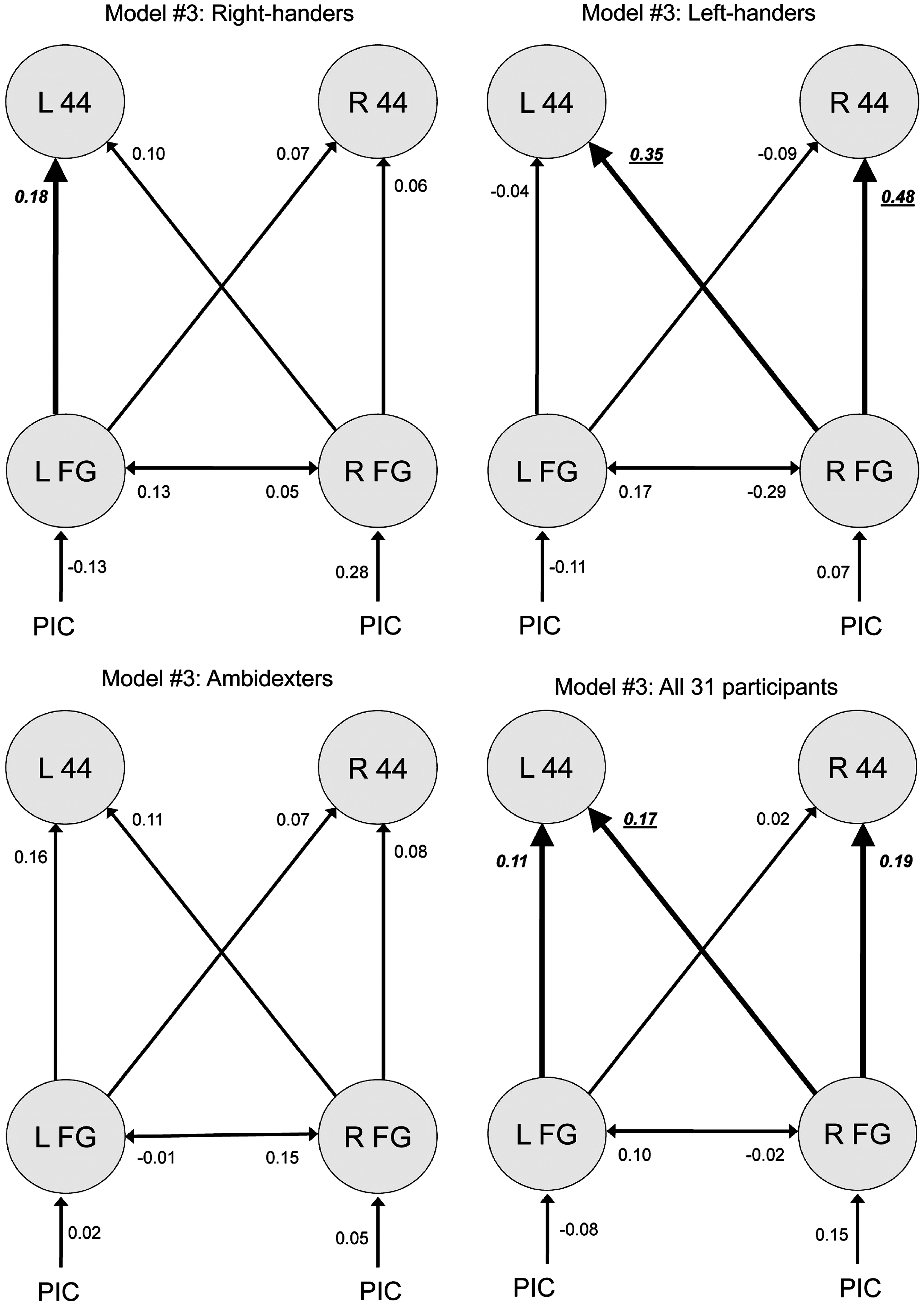
Parameters in favorable model #3 for each subject group and all 31 participants, with significant parameters being emphasized (p<0.05; with Bonferroni-correction in bold face, italics and underlined, without Bonferroni-correction in bold face and italics only).
Significant parameters are emphasized (p<0.05; with Bonferroni-correction in bold face and italics (p<0.0083 for endogenous connectivity; p<0.025 for driving input), without Bonferroni-correction in italics only). A significant difference of the parameter value of a subject group compared with the ones of another group (two-samples t-tests, p<0.05) is presented in the right column, thus indicating whether the parameter value is higher (>) or lower (<) than the parameter value that is compared (see also Supplementary Table S2 and Fig. 5). PIC, Driving input of the picture to be named.
Modulatory effects of the picture task on the connectivity between regions. Significant parameters (p<0.0083 with Bonferroni-correction) are presented in bold and italics. A significant difference of the parameter value of a subject group compared with the ones of another group (two-samples t-tests, p<0.05) is presented in the right column, thus indicating whether the parameter value is higher (>) or lower (<) than the parameter value that is compared.
Endogenous connections of model #3
All significant endogenous connections were positive (see Table 2, Fig. 4). For right-handers, the left-hemisphere connection from FG to BA 44 was significant at p<0.05 (one-sample t-tests; see also Fig. 5). In left-handers, two connections starting at right FG reached significance, targeting left and right BA 44 (p=0.004 and p=0.005, respectively). Without reaching significance, all connection strengths initiating in the left hemisphere were negative in left-handers. For the entire group of all 31 subjects, all three significant effects were found again, with significance being somewhat lower (see Table 2).

Comparison of connection strengths between subject groups in model #3. For each connection with significant strength in at least one group, a small diagram is attached that depicts strength (mean) and its possible significance for each subgroup, as well as differential effects between groups (two-samples t-tests, *p<0.05, without Bonferroni-correction; **with Bonferroni-correction). Dashed lines indicate connections without significant effects.
There were significant differences in the connections' strengths (two-samples t-tests, p<0.05) between the three subject groups as illustrated by Figure 5 (see also right column of Table 2). Significant differences between groups were limited to left-handers in comparison to the two other groups. The connections from right FG to right BA 44 was significantly stronger compared with right-handers (p=0.017) and ambidexters (p=0.026).
Moreover, we found a correlation between laterality quotient (from −100 to +100) and connection strengths: The stronger the right-handedness, the lower the connectivity between right FG and right BA 44 (Spearman's rho, rs =−0.393, p=0.029, two-tailed).
Driving inputs and contextual modulations of model #3
There were no significant effects for driving inputs (see Table 2). Regarding contextual modulations (see Table 3), there was a modulatory effect of the picture task on the connectivity from right to left FG in left-handers (p=0.004), which was stronger compared with ambidexters (p=0.006) due to the consistency of connection strengths in each group (left-handers: mean 0.01, standard error=0; ambidexters mean: −0.01, standard error=0).
Hemispheric asymmetries of model #3
Figure 4 illustrates that the network architecture of model #3 is fully symmetrical, that is, each connection has a contralateral equivalent. However, the mean of connection strengths originating either in the left or right hemisphere appeared to differ—especially in left-handers. Therefore, we statistically compared each connection in the right hemisphere to its left-hemisphere counterpart (paired-samples t-tests, p<0.05), that is, right-to-left FG connection versus left-to-right FG connection, right FG to left BA 44 connection versus left FG to right BA 44 connection, and right-hemisphere FG to BA 44 connection versus its left-hemisphere counterpart. Figure 6 illustrates the hemispheric asymmetries in connection strengths. Connections from FG to ipsilateral BA 44 in left-handers were significantly stronger on the right (in dark gray color) (p=0.032). The same held for connections originating in FG and targeting contralateral BA 44 (p=0.030). The numerical differences found for the other groups did not reach significance.
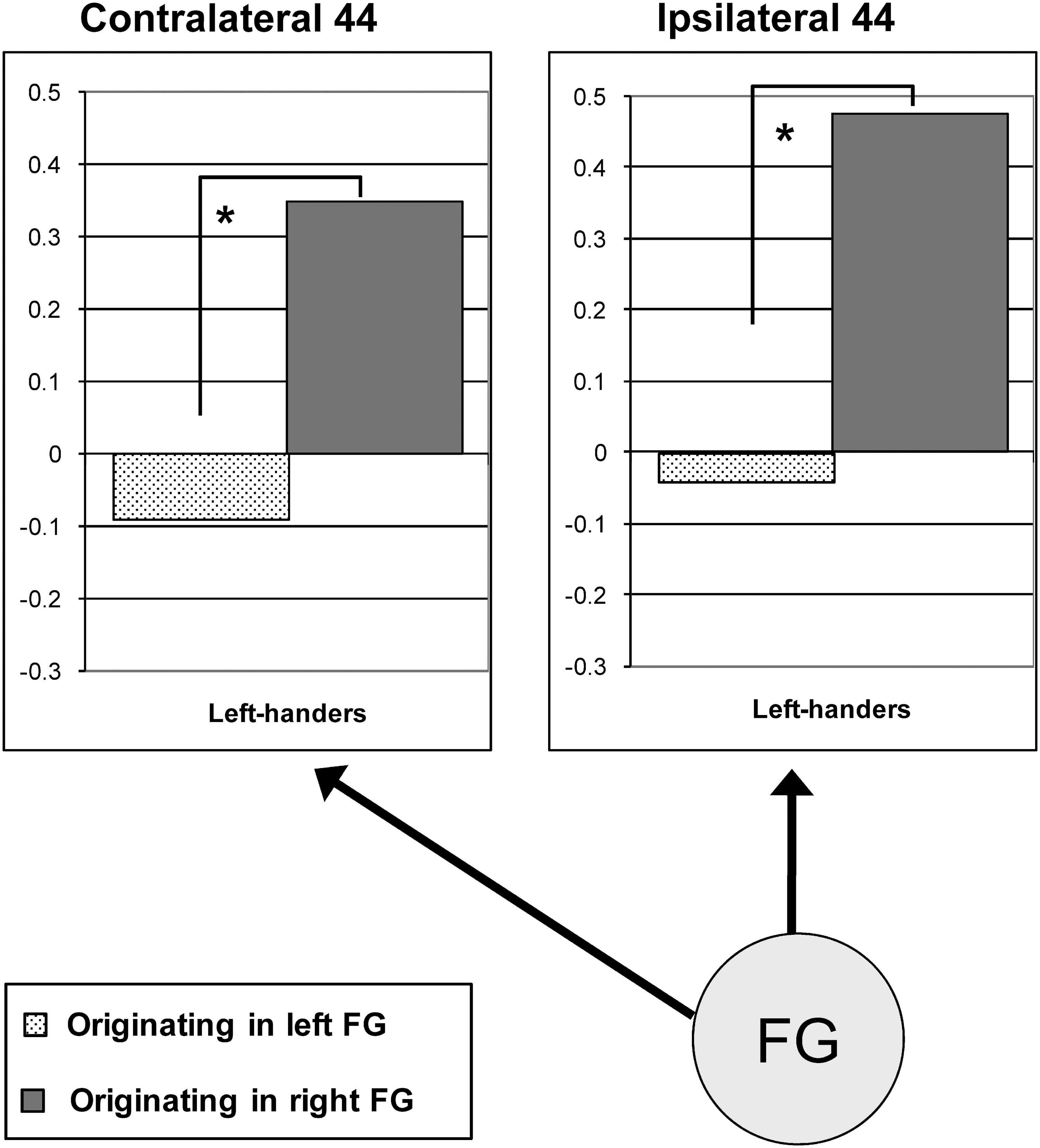
Outline of hemispheric asymmetries in connection strengths for left-handers in model #3. Each diagram represents the end point of the two equivalent connections that originate in either the left or right FG. There were significant effects for left-handers only (Paired-samples t-tests, *p<0.05, without Bonferroni-correction).
Discussion
The current study investigated the effective connectivity in the naming network within and between both hemispheres and the impact of handedness on brain connectivities. The comparison of three DCMs revealed that the same model, which implemented early integration of visual-semantic information and independent phonological processing in picture naming, was optimal for the majority of subjects in the entire group and in all subgroups (right-handers, left-handers, and ambidexters). Moreover, we found right-hemisphere dominance in connection strengths for the group of left-handers, whereas the groups of right-handers and ambidexters presented with rather symmetrical connectivities.
Initially, we re-considered the brain activations for correct picture naming in the 34 healthy subjects of a previous fMRI study (Abel et al., 2009). In the subset of 31 participants considered for the present DCM comparison, we again found widely bilateral brain activations in the GLM analysis without any lateralization associated with handedness. Thus, the well-known dominance of the left hemisphere in human language has not generally been found in the picture naming task as measured by fMRI (see also Indefrey and Levelt, 2004). Recent evidence has shown that both hemispheres exhibit word knowledge (overviews in Gazzaniga et al., 2008; Lindell, 2006). However, the specific contribution of the right hemisphere to semantic and phonological processing, as performed in picture naming, remains largely unresolved. Therefore, we examined whether an analysis of effective connectivity might uncover the role of the right hemisphere in the naming task and reveal lateralized connectivities dependent on handedness. Since the same GLM result may be modeled by different DCMs, we aimed at testing different alternative models to check which best explains the present GLM pattern. We tested two sequential theory-based hypotheses on the connectivities of the naming network. According to hypothesis (1), the brain regions of both hemispheres are differently connected dependent on the subject group. This hypothesis was shown to be incorrect, because the optimal DCM representing the connectivity pattern was basically the same for all groups. Hypothesis (2), stating that the strengths of connections between brain regions differs between groups, turned out to be correct. We found differential effects in left-handers compared with the other two groups.
The selection of the three DCM was theory based, not purely explanatory. Each of the three DCMs in the current study involved the same four brain regions assumed to reflect two main stages of overt picture naming, namely visual-semantic processing in the bilateral FG and phonological processing in the posterior part of Broca's region (BA 44) and its right homolog. These areas were also found to be activated bilaterally in previous studies on picture naming (cf. Indefrey and Levelt, 2004). For the construction of DCMs, two aspects were alternated: (i) the stages at which hemispheric information transmission occurred (early and/or late) and (ii) the presence of a direct exchange between both BA 44. Bi-directional FG connections refer to early information transfer, inter-hemispheric FG to BA 44 connections to later information transfer. The former is interpreted as early integration of visual-semantic information, the latter as transfer of semantic information for subsequent further phonological processing to the contralateral hemisphere. An absence of BA 44 interchange is interpreted as independent phonological processing in the left and right hemisphere.
For the majority of subjects in the entire group and in each subgroup, the DCM with early integration of information, with later exchange of phonological information between both BA 44 becoming rather dispensable (see model #3 in Fig. 2), was the best fit. Thus, the network architecture underlying picture naming was basically the same regardless of handedness, featuring early information transmission to the contralateral hemisphere. We further regarded the connection strengths in model #3. At first, we intend to discuss the hemispheric interactions for the two stages of picture naming for the joint analysis of the entire group of 31 participants.
For phonological processing, the semantic information was predominantly transferred from right FG to Broca's area and its right homolog, and from left FG to ipsilateral BA 44, as revealed by significant positive connection strengths. Pugh et al. (1997) suggested different roles of both hemispheres in phonological processing, with the right hemisphere being responsible for relatively small and the left hemisphere for relatively large phonological units. Poldrack et al. (1999) reported the involvement of right inferior frontal activation (including BA 44) in a syllable judgment task, indicating that the unit size of the right hemisphere should be larger than a single phoneme. The current study indicates that, in principle, right and left BA 44 may be able to process these phonological units independently, which also points to a potentially unique contribution of the right hemisphere. A crucial role of right BA 44 in language processing is supported by studies on right hemisphere compensation in aphasia recovery. A recruitment of right inferior frontal areas (pars orbitalis and triangularis) has shown to be correlated with language improvements in the subacute recovery phase (Saur et al., 2006). Moreover, Crosson et al. (2009) reported that for patients who improved in their naming treatment, activity was concentrated in the posterior portions of the right lateral frontal lobe (motor/premotor cortex and pars opercularis) after treatment. Thus, the right Broca-homolog has shown to exhibit important language faculties that might have the potential to be exploited after left-hemisphere damage. The current study confirms considerable right-hemisphere contributions to word production of healthy participants (particularly in left-handers). Interestingly, we observed that all significant endogenous connections were positive, hinting in general at facilitatory flow of information between the regions. This holds in particular for connections to right Broca homolog in left-handers, stressing its importance only in this particular sub-set of participants. The data offer a putative effective connectivity account of compensatory faculties of the right hemisphere after left-hemisphere damage, as strong connectivities of right-hemisphere regions were associated with semantic and phonological processing. Bilateral brain activations and connectivity patterns revealed that the right hemisphere was involved in lexical processing of our group of 31 healthy participants. Left-handers, who rely especially strongly on the right hemisphere, might have further developed these capacities. Thus, this right-hemisphere potential might be exploited if the left hemisphere is impaired (redundancy recovery as presented in Zahn et al., 2006). Based on the current findings, one would conclude that aphasic right-handed patients after stroke might undergo changes in their effective connectivity patterns, connecting left-hemisphere language regions more strongly to their right homologs. However, this interpretation is still rather speculative; further research will be necessary to test this prediction.
To further elucidate the impact of handedness on brain activation patterns, we intended to reveal differences in connection strengths between subject groups. Right-handers demonstrated a significant endogenous connectivity from left FG to ipsilateral BA 44, which corroborates previous findings on left-hemisphere dominance in right-handers (Knecht et al., 2000a, b; Springer and Deutsch, 2001). However, there was no significant difference in connection strength compared with another subject group or the equivalent connection of the right hemisphere, indicating rather symmetric processing.
In contrast, left-handers showed significant positive strengths of connections originating in right FG, which were significantly stronger compared with other subject groups and/or to those of the left hemisphere. Moreover, although there were increased interactions along right-to-left connections (with the FG to BA 44 connection reaching significance), there were decreased interactions along connections originating in the left hemisphere—although not significantly so. Thus, for left-handers, the information transfer between both hemispheres was strongly asymmetrical. Such asymmetry between left and right FG has previously been reported by Stephan et al. (2007b) for the language domain. These authors were able to show that the transfer of verbal information in a lateralized task was asymmetrically enhanced from the right non-specialized to the left specialized hemisphere. In our case of left-handers, a prevailing right-to-left transfer was accompanied by a predominance of right-hemisphere connectivities. Further, inter-hemispheric inhibition was exerted by the left hemisphere, as revealed by negative (non-significant) connection strengths. Its inhibitory influence has previously been maintained by Moscovitch (1977), who claimed that the regularly language-dominant left hemisphere suppresses the opposite hemisphere. In the current study, the left hemisphere tended to inhibit the right hemisphere only in case of left-handedness (and for the entire group), presumably to counterbalance the stronger right-hemisphere connectivities.
Regarding the other two subject groups, there were no significantly or, at least, substantially decreased interactions. In right-handers, the information transfer appeared to be rather symmetrical. Although visual information preferentially reached the right FG, information was distributed to all regions, with emphasis on left BA 44. In ambidexters, the right FG was least privileged by visual input, which may explain the distinctive information flow from left to right FG.
These observations underscore that, although language production is to some extent a motor action, its nature very much differs from other motor tasks. Although in hand motor tasks (e.g., Grefkes et al., 2008a), for example, symmetrical reciprocal inhibition between the motor cortices of both hemispheres is observed, the pattern for Broca's region and its homolog in the current study was different. In line with the underlying assumptions from the Levelt model (e.g., Indefrey and Levelt, 2004) and earlier work on cognition-motor-interactions during language production (Eickhoff et al., 2009), the data show that Broca's region exhibits an activation pattern distinct from that of real motor areas (Grefkes et al., 2008a), and rather more similar to that of cognitive regions (e.g., Siman-Tov et al., 2007; Stephan et al., 2003).
We also would like to discuss a non-significant but highly interesting result. Numerically, the dominance of input to right-sided FG was observable for the entire group (and even for each subgroup) (see Fig. 4). Even though these results did not reach significance, they may inform us about the information flow in model #3. They may indicate that visual information of the picture to be named tends to be predominantly prepared in right FG regardless of handedness. Previous results on visual object priming in the left or right visual field revealed dissociable object recognition subsystems in both hemispheres (Marsolek, 1999), that is, a specific-exemplar subsystem operates more effectively on the right and an abstract-category subsystem on the left. Using fMRI, Koutstaal et al. (2001) were able to show that the specific and abstract distinction is localized in right and left FG, respectively. Thus, in our paradigm, visual-semantic processing might particularly rely on right FG, because the depicted objects to be recognized were concrete. However, left-hemisphere processing may be additionally required to map the specific picture to a category, which accounts for the bilateral activation of FG in the GLM analysis and the superiority of the present model featuring early visual-semantic integration. However, main peak of brain activation was in left and not right FG. The discrepancy between GLM and DCM analysis might indicate that processing in left FG is more effortful, thus requiring stronger activation amplitudes, whereas the right FG has a central role requiring stronger connectivities.
In general, some variability in the lateralization of connectivities even within the three groups was quite pronounced (see also Allen et al., 2008). This also holds for right-handers, even though their lateralization has been reported to be less variable than the one of left-handers (Springer and Deutsch, 2001). We attribute the unexpected, principally symmetrical connectivities found in right-handers and ambidexters to the task. Picture naming involves the processing of concrete items that is supported by not only the left but also the right hemisphere. Further, the stronger right-hemisphere connectivities between visual-semantic and phonological stages found in left-handers correspond to previous findings of a more frequent right-hemisphere language dominance in left-handers (Knecht et al., 2000a, b ; Springer and Deutsch, 2001). This prevalence of right-hemisphere connectivities may be the mechanism underlying the better prognosis for recovery from aphasia in left-handers compared with right-handers (Springer and Deutsch, 2001).
One limitation of the study is the missing examination of gender-related differences in activation patterns. However, this does not exclude possible connectivity differences caused by gender. This cannot be further examined, as we did not control for hormonal status in women, and there would not be enough participants left in each group to perform group analyses.
Conclusion
To conclude, this is to our knowledge the first investigation of the relation between brain asymmetry and handedness that regards differences in effective connectivity. We observed a difference in brain laterality by using DCM, which could not be found in the standard analysis of brain activation with GLM in correct overt picture naming. There was no difference in the nature of the connectivity patterns, but rather in the strengths of connectivities. For the majority of subjects in the total sample as well as in each subgroup, the DCM was optimal that combined early visual-semantic exchange between both FG, later information transfer to both BA 44, and independent phonological processing in each BA 44.
Right-handers presented with a strong left-hemisphere FG to BA 44 connectivity. However, there was a relatively symmetric information transfer for both the groups of right-handers and ambidexters. On the contrary, the group of left-handers showed stronger connection strengths in the right compared with the left hemisphere. Therefore, the bilateral brain activations observed in the GLM analysis were mirrored by the rather symmetrical connectivities in right-handers and ambidexters, but not by the asymmetric ones for left-handers. Finally, the current study supports previous findings on significant and unique right-hemisphere contributions to word production and may offer an effective connectivity account of compensatory lexical faculties of the right hemisphere after left-hemisphere brain damage. The right hemisphere again has shown to be better than its reputation in the language domain.
Footnotes
Acknowledgment
Financial support for this project was provided by the Federal Ministry of Education and Research (BMBF) to WH (support code 01GW0662).
Author Disclosure Statement
No competing financial interests exist.
*
Since Bonferroni-correction is extremely conservative in the presence of posterior dependencies among parameters tested and is even dispensable if a single contrast is applied, that is, a linear combination of these parameters (Stephan et al., ![]() ), uncorrected results (p<0.05) are also taken to be significant.
), uncorrected results (p<0.05) are also taken to be significant.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.