
Abstract
Brain injury and subsequent plasticity of sensory and corticospinal pathways play an integral role in determining paretic hand function in congenital hemiplegia. There is limited knowledge regarding the relationship between the disruption of sensorimotor thalamic pathways projecting into the primary motor cortex and motor control. This study sought to investigate the relationship between the structural connectivity of motor networks that anatomically link the brain stem with the precentral and postcentral gyri with paretic motor sensory function by using an automated analysis strategy. Magnetic resonance imaging structural connectivity was measured by using high-angular-resolution diffusion imaging, probabilistic tractography, and the anatomic parcellation of high-resolution structural images in 16 children with congenital unilateral periventricular white-matter damage. Connectivity of the corticospinal and corticothalamic pathways was determined by using an asymmetry index based on the number of streamlines contained within these projections and compared with measures of paretic hand function and bimanual coordination. For cortical development, the volume of the ipsilesional precentral gyrus was significantly reduced. For connectivity measures, the numbers of streamlines in corticospinal tracts and corticothalamic pathways within the ipsilesional hemisphere were decreased compared with the contralesional side. The sensorimotor thalamic projections were more significantly correlated with paretic hand functions than were the corticospinal tracts. These data support the concept that preservation of sensorimotor thalamic pathways that directly project into the primary motor cortex has more influence on motor function control of the paretic hand than does preservation of corticospinal tracts.
Introduction
U
No studies have explored specific components of the thalamo-cortical pathways in children with congenital hemiplegia, and in particular the distinction of the projections to the somatosensory cortex from those to the primary motor cortex, which are known to play a different role in the neurophysiology of movement (Gharbawie et al., 2010). This study sought to investigate the relationship between the structural connectivity of the corticospinal tract and the thalamocortical projections to the precentral and the postcentral gyrus, with clinical measures of hand function and control. To achieve this goal, the study used a fully automated method for measuring the connectivity of white-matter pathways within this motor feedback system using high-angular-resolution diffusion imaging (HARDI) with constrained spherical deconvolution, which describes diffusion in complex white-matter networks using a fiber orientation distribution function (Tournier et al., 2007). Whole-brain probabilistic tractography was applied to define the corticospinal tract and thalamocortical projections involving the precentral and the postcentral gyri, which were delineated from the anatomic parcellation of high-resolution structural images. This approach is an extension of previously reported strategy for structural connectivity analysis (Pannek et al., 2010).
Materials and Methods
Participants
MRI data were acquired from 20 participants with congenital hemiplegia. However, only 16 high-resolution data sets could be satisfactorily anatomically parcellated by using Freesurfer (Athinoula A. Martinos Center for Biomedical Imaging;
Data expressed with a plus/minus sign are mean summary measures±standard deviation.
For normalization, corticomotor volumes have been divided by the intracranial volume.
AHA, Assisting Hand Function; AI, asymmetry index for the corticospinal tracts and corticothalamic sensory pathways; CST, corticospinal tract; JHFT, Jebsen Test of Hand Function; MUUL, Melbourne Assessment of Unilateral Upper Limb Function; preCG, precentral gyrus; postCG, postcentral gyrus.
MRI protocol
MRI data were acquired by using a 3T Siemens TimTrio (Siemens). Along with several radiologic scans, a high-resolution structural image was acquired for each participant by using a 1-mm3 isotropic three-dimensional T1 MPRAGE (field of view, 24×25.6×17.6 cm; repetition time/echo time/inversion time (TR/TE/TI), 2300/2.26/900 ms; flip angle, 9°). The imaging time was 9:14 min. The HARDI parameters were as follows: number of axial slices, 60; field of view, 30×30 cm; TR/TE, 9200/112 ms; slice thickness, 2.3 mm; acquisition matrix, 128×128 with a 2.3-mm in-plane image resolution; acceleration factor, 2; and a diffusion encoding gradient strength, b=3000 s·mm−2. Sixty-eight diffusion-weighted images were acquired at each location, consisting of eight low (b=0) and 60 high diffusion-weighted images. The acquisition time for the diffusion data set was 9:40 min. A field map was acquired by using two two-dimensional gradient recalled echo images (TE1/TE2, 4.76/7.22 ms) to assist the correction for distortion due to susceptibility inhomogeneity.
MRI analysis
Structural images
Cortical parcellation was performed on structural images with the Freesurfer image analysis suite in native space. Intensity inhomogeneity of the images was corrected (Sled et al., 1998) and nonbrain tissue was removed by using a hybrid watershed/surface deformation procedure (Segonne et al., 2004). The cerebral cortex was parcellated into 33 units per hemisphere based on gyral and sulcal structure (Desikan et al., 2006; Fischl et al., 2004) along with a mask that delineated the brainstem structures that made up the cerebral peduncles. This automated cortical parcellation procedure is similar to manual delineation of regions in terms of accuracy (Desikan et al., 2006). The Freesurfer delineated brainstem masks were visually inspected for each participant and manually edited where required. A termination mask for tractography was generated from the structural image to prevent streamlines from crossing the cortical folds (Pannek et al., 2010).
Diffusion processing
Diffusion-weighted images were corrected for eddy current distortions by using tools provided with FMRIB's Diffusion Toolbox (FDT, part of FMRIB Software Library FSL;
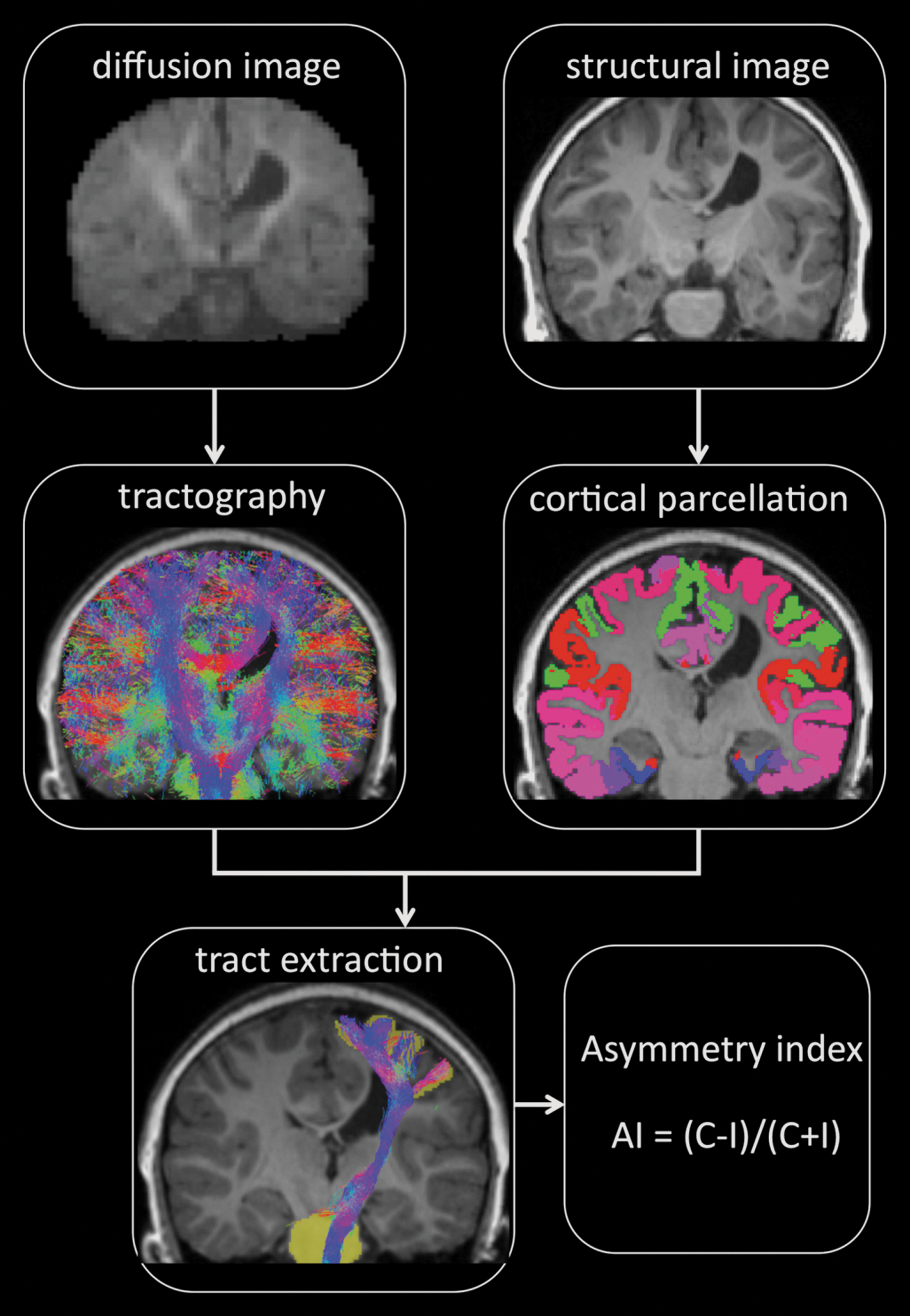
Steps involved with the automated analysis of connectivity measures. AI, asymmetry index; C number of streamlines within the corticospinal tract and sensory pathways for the contralesional hemisphere; I, number of streamlines within the corticospinal tract and sensory pathways for the ipsilesional hemisphere.
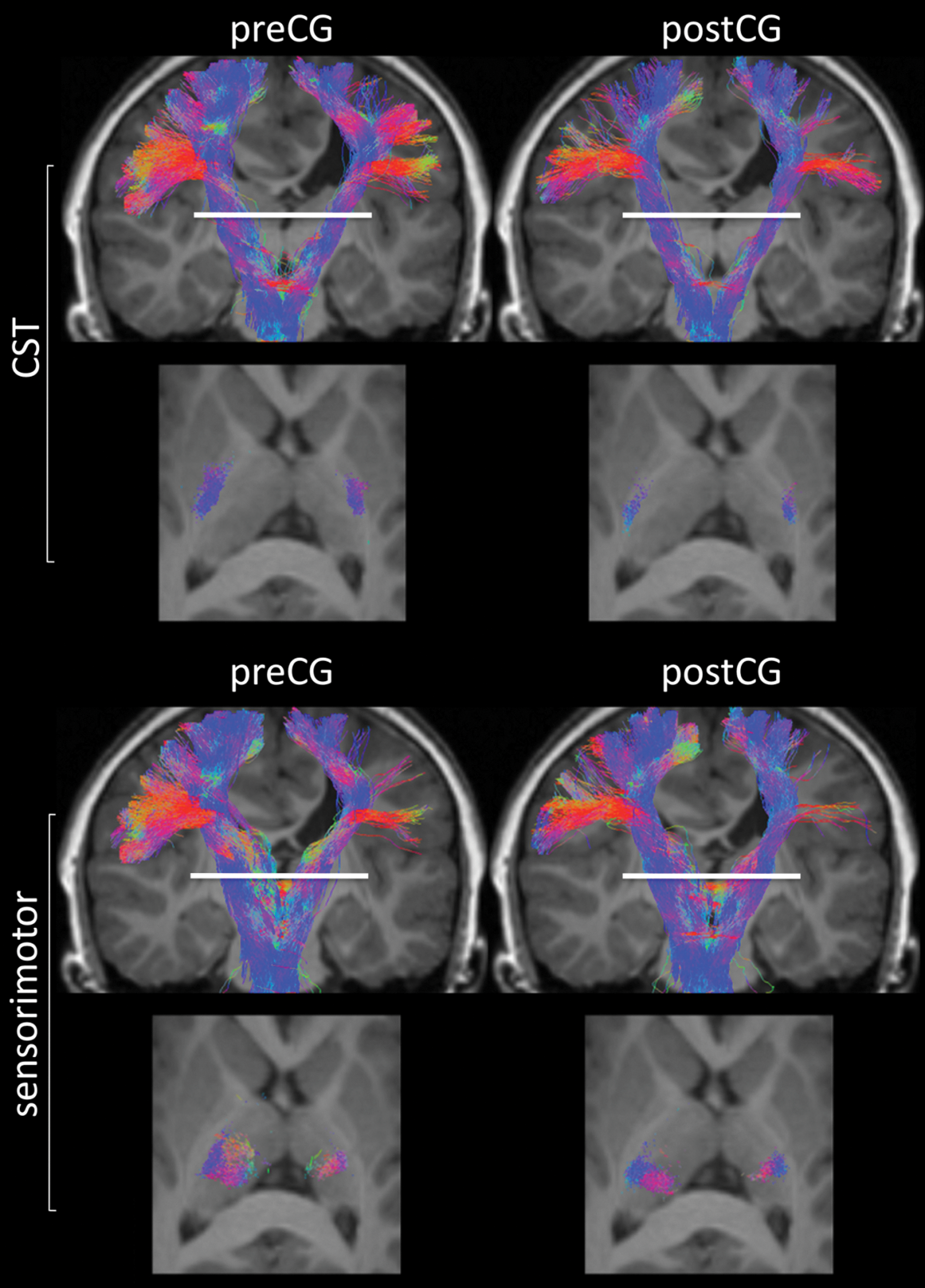
Representative images showing the automated delineation of primary motor (CST) and sensorimotor pathways for a participant with right hemiplegia. The two top rows of images represent all corticospinal tracts that project between the brain stem and the precentral (preCG) (left) and postcentral (postCG) gyri (right), and the two bottom rows show similar images but for the sensorimotor pathways that project via the thalamus. The axial images show projections through the posterior limb of the internal capsule (top) and the ventral lateral nucleus (bottom) at the level of the internal capsule (white line).
Connectivity measures and statistical analysis
To ensure generation of connectivity indices in diffusion space, the whole-brain track density maps were nonlinearly registered (Andersson et al., 2008) to the structural image, and the inverse transformation was applied to the parcellated cortical masks. The accuracy of nonlinear registration of the track density map to structural image was assessed for each participant before proceeding to the next processing step. A termination mask was applied to prevent streamlines from crossing cortical folds. White-matter pathways, connecting the multiple cortical regions, were then generated by hit-testing every streamline's terminal ends with every cortical parcellation (Pannek et al., 2010; Park et al., 2008). From this whole-brain analysis, streamlines that represent the corticomotor pathways linking the Freesurfer-defined precentral gyrus brainstem mask and postcentral gyrus brainstem mask were automatically extracted.
To differentiate the sensory from the corticospinal pathways, a thalamic mask generated from the Freesurfer parcellation was used as an inclusion mask to ensure that the sensory pathways included only streamlines that projected through the thalamus. In contrast, the thalamic mask was used as an exclusion mask to capture only those corticospinal streamlines passing through the posterior limb of the internal capsule. By using this approach, two separate corticospinal pathways linking the precentral gyrus brainstem and postcentral gyrus brainstem via the posterior limb of the internal capsule were investigated. Two separate sensory pathways linking the precentral gyrus brainstem and postcentral gyrus brainstem via the thalamus were also investigated. The same brainstem mask was used to delineate both the corticospinal tract and corticothalamic white-matter pathways. Figure 2 shows representative examples of these corticomotor pathways. Because the streamline counts for these projections are not quantitative (Jones 2010), an asymmetry index was generated for each pathway for every participant using the following expression:
where C and I represent the number of streamlines within the corticospinal tract and sensory pathways for the contralesional and ipsilesional hemispheres, respectively. In each case, each of the sensory and corticospinal tract projections was visually inspected to ensure that no false-positive streamlines were contained within the white-matter pathways.
For assessment of volumetric measures of the precentral and postcentral gyri, derived from the Freesurfer cortical parcellation, asymmetry indexes were generated by using an equation similar to that given above. In this case, C and I refer to the normalized volumes, based on the intracranial volume, for the precentral and postcentral gyri within the contralesional and ipsilesional hemispheres, respectively. The relationship between the asymmetry index and hand function scores were assessed by using the Pearson product-moment correlation. Correlations were deemed significant after correction for multiple comparisons based on a false discovery rate of 10%. Differences in volumes of the precentral gyrus and postcentral gyrus between the ipsilesional and contralesional hemispheres were determined by using a two-tailed t-test (IBM SPSS Statistics [Version 19.0]).
Results
Demographic information and clinical motor function scores for the impaired hand are reported in Table 1. When cortical measures was assessed, there was a significant reduction (p=0.016) in the normalized intracranial volume of the ipsilesional precentral gyrus compared with the contralesional hemisphere (Table 1). Volume of the postcentral gyrus did not significantly differ between the ipsilesional and the contralesional hemisphere (p=0.347). For correlations of the asymmetry indexes derived from normalized volumes of the precentral gyrus with motor functional measures of the impaired hand, a significant negative correlation was found for the AHA (r=−0.61; p=0.0015), see Table 2. The correlation between the asymmetry index derived for the normalized volumes of the precentral gyrus and the JHFT was not significant after correction for multiple comparisons (r=0.52; p=0.0421). There were no correlations between the asymmetry indexes derived for the postcentral gyrus and paretic hand function scores that reached a level of significance (Table 2). The significant negative correlation with the AHA suggests that a reduction in volume of the precentral gyrus within the affected hemisphere is associated with reduced performance on the motor task.
Correlations in bold are statistically significant after correction for multiple comparisons: * p=0.0003; ** p=0.0061; *** p=0.0013; **** p=0.0015
AHA, Assisting Hand Function; JHFT, Jebsen Test of Hand Function; MUUL, Melbourne Assessment of Unilateral Upper Limb Function; PLIC, corticospinal tract projecting through the posterior limb of the internal capsule; pre-CG, cortical volume of the precentral precentral gyrus; post-CG, cortical volume of the postcentral gyrus; sensorimotor (thalamic), corticothalamic sensory pathways.
The mean asymmetry index derived for the connectivity measures for the corticospinal tract and sensory white-matter pathways are given in Table 1. The magnitude of the mean asymmetry index indicates a reduction in motor and sensory streamlines within the ipsilesional hemisphere compared with the contralesional side for both the precentral and the postcentral gyrus. Correlations between asymmetry index connectivity measures and functional hand scores are presented in Table 2, and scatter plots are provided in Figure 3. For the corticospinal tracts projecting from the precentral and postcentral gyrus, no correlations with impaired hand function were statistically significant. However, for the thalamic sensorimotor pathways projecting into the precentral gyrus, there were significant correlations between the asymmetry indexes derived for these connections and all motor performance scores. The functional correlations for the sensorimotor pathways projecting into the precentral gyrus for the JHFT, MUUL, and AHA were r=0.80 (p=0.0003), r=−0.67 (p=0.0061), and r=−0.62 (p=0.0013), respectively.
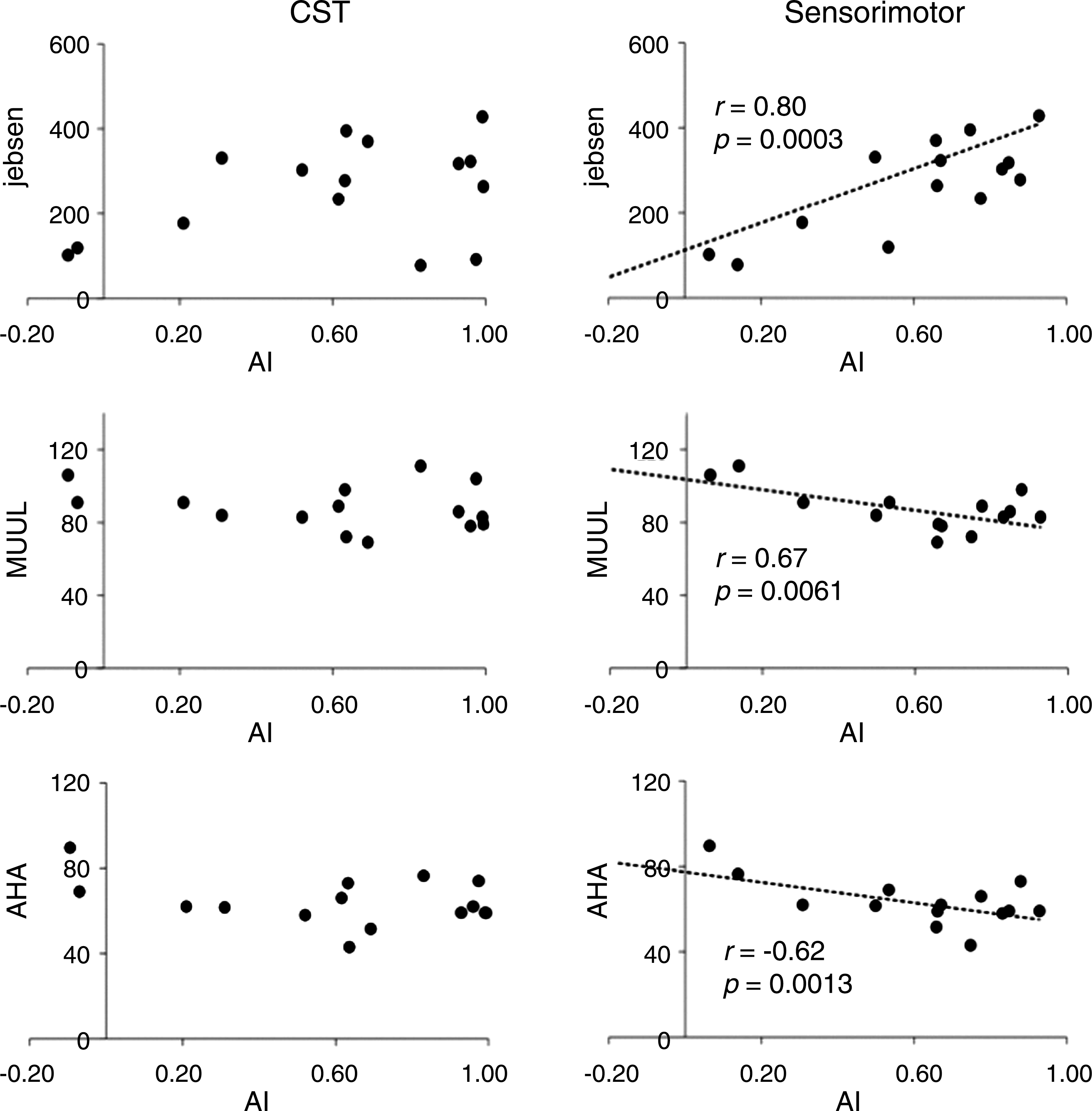
Scatter plots showing the correlation of asymmetry indexes (AI) derived for both corticospinal tract (CST; left) and thalamic sensory pathways (right) projecting into the precentral gyrus with motor function scores. AHA, Assisting Hand Function; Jebsen, Jebsen Hand Function Test; MUUL, Melbourne Assessment of Unilateral Upper Limb Function.
Correlations for the asymmetry indexes derived for the sensorimotor pathways of the postcentral gyrus were not significant. On the basis of an asymmetry measure, these results indicate that an increased number of streamlines within these sensorimotor pathways in the ipsilesional hemisphere may be associated with improved motor performance of the paretic hand. Furthermore, the significant positive correlations between sensory asymmetry index measures derived for the precentral gyrus and JHFT scores, along with the strong negative correlations with the MUUL and AHA scores, highlight the importance of preservation of sensorimotor thalamic pathways that project into the primary motor cortex in maintaining paretic hand motor function.
Discussion
This study introduces a new automated analysis, based on MRI structural connectivity, to investigate the relationship between the integrity of motor-sensory pathways that project into the primary motor and somatosensory cortex and paretic hand function in children with congenital hemiplegia. This strategy relies on the use of HARDI with constrained spherical deconvolution, which describes the diffusion of water in complex white-matter networks using a fiber orientation density function (Tournier et al., 2007). In conjunction with whole-brain probabilistic tractography and information describing the anatomic parcellation of cortical structures, specific white-matter pathways projecting from and into the primary motor and somatosensory cortex were investigated. Such an approach reduces the known limitations of the standard tensor model, which fails to adequately resolve crossing fiber populations (Jones 2010; Tournier et al., 2007) and the need for manual placement of regions of interests to direct fiber tracking. This approach has been successfully used to anatomically parcellate the corpus callosum on the basis of mapping interhemispheric commissural projections (Pannek et al., 2010). Because this approach relies on the accurate parcellation of anatomic structures, a limitation of this automated strategy is its inapplicability to study participants with significant cortical pathologic abnormalities or ventricular enlargement due to excessive white-matter injury. However, of the 20 participants recruited into this study, data from 16 patients were successfully analyzed.
An example of the white-matter tracts involved with the motor-sensory feedback loop that were generated using the automated connectivity analysis is shown in Figure 2. There is clear delineation of the corticospinal tracts projecting through the posterior limb of the internal capsule and sensorimotor pathways projecting through the thalamus. The anatomic location of the sensorimotor pathways that project via the thalamus (i.e., precentral gyrus connections projecting anterior to postcentral gyrus connections) agree with results reported from other studies targeting the functional parcellation of the human thalamus using structural and functional connectivity measures (Behrens et al., 2003; Broser et al., 2011; Johansen-Berg et al., 2005; O'Muircheartaigh et al., 2011). Furthermore, it is well known from retrograde tracer studies in primates that axons within the sensorimotor system contain projections that transfer sensory input from dorsal horn cells to primarily the ventral lateral nucleus, along with other regions within the thalamus (Martin et al., 2007). Similarly, corticothalamic axons project from the ventral lateral nucleus to layer V within the primary motor cortex to provide sensorimotor feedback (Deschênes et al., 1994; Stepniewska et al., 1994). Retrograde tracer studies in the macaque brain have shown that the ventralis posterior lateralis oralis, which also receives input from the cerebellum, provides the most substantial input to a portion of the hand representation on the precentral gyrus (Holsapple et al., 1991; Shindo et al., 1995). In contrast, the ventralis lateralis oralis, which also receives input from the basal ganglia, provides input to supplementary motor areas (Shindo et al., 1995). The dominant hypothesis as to the function of the pathways involving the ventralis posterior lateralis oralis is that it helps to generate movements in reaction to external stimuli, while the ventralis lateralis oralis pathways assist in the generation of internally triggered willful movements (Middleton and Strick, 2000). It is thus not surprising that the involvement of these pathways results in a higher impairment of hand function, which generally involves both of these components.
With regard to the corticospinal tract, the number of corticospinal streamlines projecting from the ipsilesional motor cortices was reduced. This is a consistent finding in many studies (Eyre 2007; Hoon et al., 2009; Staudt et al., 2002; Staudt 2010; Thomas et al., 2005; Trivedi et al., 2010; Yoshida et al., 2010). All participants had radiologic evidence of unilateral lesions within periventricular white-matter regions, with transcranial magnetic stimulation studies showing the presence of crossed corticospinal projections from the rolandic cortex of each hemisphere. The magnitude of the asymmetry index for these motor pathways supported the concept of a reduction in corticospinal tract projections from the ipsilesional precentral and postcentral gyrus.
It is important to emphasize that MRI diffusion tractography measures are not quantitative and that streamline count should not be used as a quantitative measure of connectivity for group-wise analyses (Jones 2010). For this reason, the current study used a summary measure of connectivity based on the ratio (asymmetry index) of the number of streamlines in motor pathways within the impaired versus nonimpaired hemispheres. This study did not investigate the integrity of corticomotor white-matter pathway using measures of fractional anisotropy and mean diffusivity. Previous studies have shown that summary indexes based on streamline count are more sensitive measures for investigating altered corticomotor white-matter pathways in children with congenital hemiplegia compared with fractional anisotropy or mean diffusivity (Thomas et al., 2005; Yoshida et al., 2010).
The most novel finding from this study is the reduction in the number of thalamo-cortical pathways projecting into the primary motor cortex within the affected hemisphere. The concept of a disruption of somatosensory pathways, especially within neural circuits incorporating the posterior thalamic radiations, in cerebral palsy has been reported previously (Hoon et al., 2009; Yoshida et al., 2010). However, the current results show for the first time the disruption of thalamo-cortical white-matter pathways that project directly into the sensorimotor cortex. As shown in Figure 2, the reduction in motor and sensorimotor tracts within the affected hemisphere is apparent with the surviving white-matter tracts projecting around the site of white-matter damage. In addition, the thalamo-cortical projections correlated significantly with hand function, as assessed with different functional scales. Importantly, the data support the concept that the motor system requires feedback from sensory systems to shape the development of the motor cortex and efferent motor pathways (Eyre 2007; Martin et al., 2007). This model is extended by showing that preservation of sensory pathways that directly project into the primary motor cortex from the impaired arm may have more influence on motor function control of the paretic hand than does preservation of corticospinal tracts.
From these findings it is not possible to determine whether the reduction of thalamo-cortical fibers in the impaired side is the direct result of the brain damage or, rather, the effect of postlesional mechanisms of plastic reorganization. Recent studies have demonstrated that motor outcome in children with unilateral brain damage is related to neuroplastic processes that are, at least partially, independent of the size and site of the lesion, and are heavily influenced by the levels of postlesional activity within the perilesional cortex (Eyre 2007). It cannot be excluded that similar mechanisms may affect the reorganization of thalamocortical fibers.
Further work involving more participants with congenital hemiplegia is required to fully validate these findings. The effect of sex on the importance of sensory feedback within the developing motor system is unclear. This cohort had a sex imbalance but similar numbers of patients with left and with right hemiplegia. Again, larger studies will help resolve these issues. These results highlight the importance of the preservation of sensorimotor thalamic projections into the primary motor cortex in maintaining hand function in children with congenital hemiplegia. At this stage, the relationship between the loss of sensorimotor pathways and volume of the thalamus has not been determined. However, such markers of thalamic development that can be reliably measured in a serial fashion using ultrasonography in preterm babies (Fukuda et al., 2010) may have a significant clinical effect in the early diagnosis of motor dysfunction in high-risk neonatal groups. The clinical relevance of our findings is important for neurorehabilitation therapies: activity-dependent processes involving the sensory system may promote improved motor function in congenital hemiplegia (Johnston 2009).
Conclusion
This article presents an automated strategy, based on MRI structural connectivity, used to investigate the relationship between paretic hand function and the integrity of corticomotor white-matter pathways of the primary and somatosensory motor cortex in children with congenital hemiplegia. The sensorimotor thalamic projections correlated more significantly with paretic hand functions than did the corticospinal tracts. This finding supports the concept that preservation of sensorimotor thalamic pathways that directly project into the primary motor cortex has more influence on motor function control of the paretic hand than does preservation of corticospinal tracts.
Footnotes
Acknowledgments
The authors acknowledge the National Health and Medical Research Council of Australia for assisting in funding this research project.
Author Disclosure Statement
The authors have no conflicts of interest to declare.