
Abstract
The last 10 years witnessed a considerable increase in our knowledge of brain function in survivors to severe brain injuries with disorders of consciousness (DOC). At the same time, a growing interest developed for the use of functional neuroimaging as a new diagnostic tool in these patients. In this context, particular attention has been devoted to connectivity studies—as these, more than measures of brain metabolism, may be more appropriate to capture the dynamics of large populations of neurons. Here, we will review the pros and cons of various connectivity methods as potential diagnostic tools in brain-damaged patients with DOC. We will also discuss the relevance of the study of the level versus the contents of consciousness in this context.
Introduction
T
DOC remain among the most poorly understood and less treatable conditions of modern neurology. They are commonly divided into three main clinical entities. Coma is defined by a lack of both arousal and awareness. Vegetative state (VS) is defined by a lack of behavioral signs of consciousness, while arousal (defined as an alternation of eye opening and closing) is preserved (Laureys et al., 2004). In contrast, a minimally conscious state (MCS) is defined by the presence of nonreflexive behaviors, although patients remain unable to communicate (Giacino et al., 2002). Recent studies showed that in the absence of use of an appropriate clinical scale such as the Coma Recovery Scale–Revised (CRS-R), the risk of underestimating the level of consciousness in DOC patients is about 40% (Schnakers et al., 2009). This fact illustrates the difficulties inherent to the behavioral assessment in these patients. Many patients' behaviors, such as grimacing or localization of sounds, for example, remain equivocal and difficult to interpret as evidence for the presence or absence of consciousness. Research, therefore, aims at finding reliable brain activity markers of consciousness that are applicable to these patients. Finding such markers would improve both diagnosis and disease understanding, bringing possibilities for the setup of new treatments (Boly, 2011). Functional neuroimaging studies performed in unresponsive brain-damaged patients can be categorized into two main classes: active and passive paradigms (Boly and Seth, 2012). Active paradigms aim at obtaining a response to command in patients, while bypassing the motor response itself (Boly et al., 2007; Owen et al., 2006). Passive paradigms gather information on the patients' brain function, identifying neural activation reflecting “automatic” processing, that is, occurring without the patients' willful intervention (Boly et al., 2009a). [18F]-fluorodeoxyglucose positron emission tomography (PET) studies showed a widespread impairment of fronto-parietal cortices in VS patients as compared with controls (Laureys, 2005; Laureys et al., 1999). Although metabolic studies certainly provide valuable information, connectivity may, however, better grasp the neurons' collective behavior. Brain connectivity can be studied at both the structural (Fernandez-Espejo et al., 2011) and functional (Noirhomme et al., 2010; Soddu et al., 2009, 2011) levels. Functional studies are not necessarily trying to univocally reflect structural connectivity. Although recent attempts at predicting functional from structural connectivity are very promising (Honey et al., 2007), some discrepancies may exist when comparing the predicted functional connectivity with the empirical results obtained by measuring the resting-state blood-oxygen-level-dependent (BOLD) signal through functional magnetic resonance imaging (fMRI). The present review will focus on the functional aspects.
On the functional side, it may be useful to introduce here some relevant concepts that will be further discussed. Functional connectivity measures the deviations from statistical independence among distant neural populations. Statistical dependence is estimated based on temporal relationships, by applying nondirected (for example, linear correlation and coherence-based analyses) or directed (Granger causality and transfer entropy) measures to time series of neuronal activity (Bressler and Seth, 2011). Measures of functional connectivity are largely model free but do not make any explicit reference to the underlying structural model. Hence, for instance, functional connectivity among distant elements may be high either because these elements are actually connected by direct structural links or just because they share a common input. On the other hand, effective connectivity attempts to extract networks of causal effects among neural elements. Causal effects can be inferred directly by detecting the large-scale effects produced by a controlled (peripheral, cortical-electric, and cortical-magnetic) perturbation of the system. Alternatively, patterns of effective connectivity can be extracted from spontaneous or evoked activity by means of dynamic causal modeling (DCM). This approach requires specifying a model, including structural parameters, that informs on the original neuronal mechanism at the basis of the observed signals. The next section will review how these different approaches to the study of connectivity may be employed in DOC patients.
Studying Brain Connectivity in DOC—the Different Approaches
To date, three different connectivity approaches have been used in patients with DOC: (1) functional connectivity or “resting state” studies using, for example, fMRI; (2) a perturbational approach using transcranial magnetic stimulation (TMS) combined with electroencephalography (EEG); and (3) other perturbational approaches investigating brain responses to external stimuli, including sensory evoked potentials (ERP). After summarizing the findings obtained in DOC with each of these techniques, we will review their respective practical strengths and weaknesses. Figure 1 condenses the main concepts of the present section.

Pro (+) and cons (−) of different approaches applicable to the study of brain connectivity in patients with disorders of consciousness. Detailed description can be found in the main text. Upper panels are reproduced with permission from (Boly et al., 2011; Massimini et al., 2009; Vanhaudenhuyse et al., 2010). fMRI, functional magnetic resonance imaging; TMS, transcranial magnetic stimulation; EEG, electroencephalography; ERP, evoked potentials; DCM, dynamic causal modeling; BA, Brodmann area; A1, primary auditory cortex; STG, superior temporal gyrus; IFG, inferior frontal gyrus. Right panel: brain area 1=A1, area 2=STG, area 3=IFG. Arrows represents connection between the different network nodes as for example studied by DCM.
Functional connectivity studies
Functional connectivity investigates linear relationships among the measured activities in the different regions of interest of the brain. Both univariate, comparing each time pairs of regions, and multivariate, assessing all the regions at the same time, approaches are nowadays available, each with its own advantages and limitations. Functional connectivity can, in theory, be applied to a variety of brain imaging techniques. Resting-state fMRI functional connectivity has proved useful in the identification of a correlation between the fronto-parietal networks connectivity and the level of consciousness in DOC patients (Boly et al., 2009b; Fernandez-Espejo et al., 2010; Vanhaudenhuyse et al., 2010) or anesthesia (Boveroux et al., 2010; Martuzzi et al., 2010; Schrouff et al., 2011). Some decreases in fronto-parietal connectivity have been described in deep nonrapid eye movement (non-REM) sleep (Horovitz et al., 2009; Samann et al., 2011). Other studies, however, show globally preserved connectivity across sleep stages (Koike et al., 2011). Anticorrelations between a default network and lateral fronto-parietal cortices have also been shown to decrease in VS (Boly et al., 2009b), anesthesia (Boveroux et al., 2010), and non-REM sleep (Samann et al., 2011) as compared with normal wakefulness.
Although this technique is very useful in collecting information about whole-brain function in one single data set, the functional significance of resting-state fMRI signals has been debated (Birn et al., 2008). It is now commonly accepted that the BOLD signal tightly correlates with population-level neuronal activity, especially with regard to local field potentials (Nir et al., 2007, 2008). The relationship between the BOLD signal and regional electrophysiological activity is likely to be similar for task-related fMRI or the resting state. Consequently, the functional significance of spontaneous or task-evoked BOLD fluctuations is also likely to be similar. Resting-state and task-related fMRI are prone to noise contamination, such as by movement and physiological artifacts, which are difficult to avoid in HV as in DOC patients (Soddu et al., 2011, 2012). Indeed, DOC patients are noncollaborative and may present uncontrollable reflexive movements. Even if different methods exist for dealing with them (Fox et al., 2005; Jones et al., 2008; Perlbarg et al., 2007; Soddu et al., 2012), none of them are completely satisfactory. fMRI temporal resolution is also slower than the temporal scale where conscious scenes are likely to arise. This temporal scale is indeed thought to be of the order of hundreds of milliseconds, rather than seconds. fMRI studies could, thus, be usefully complemented by EEG.
Resting-state spontaneous brain activity analyses can also be performed using EEG. The loss of consciousness during non-REM sleep or anesthesia is usually associated with a raise of delta power and the occurrence of slow waves in the EEG (Murphy et al., 2011). These slow waves travel across a wide range of brain areas, especially in the default network (Murphy et al., 2009, 2011). The bispectral index is a commercial EEG-based measure that is mainly based on power spectral changes in the EEG between different states, and for small value ranges is largely based on changes in power in the delta band. The fact that the bispectral index can differentiate between MCS and VS patients (Schnakers et al., 2008) also suggests a negative correlation between the EEG delta power and the level of consciousness in DOC patients. A small number of studies have described decreased scalp EEG coherence in coma (Kane et al., 1998), and in one VS (Davey et al., 2000) and one MCS (Kobylarz and Schiff, 2005) patient. Recent reports have, however, described increased EEG coherence during propofol-induced loss of consciousness (Murphy et al., 2011; Supp et al., 2011). Granger causality estimates causal statistical influences between simultaneously recorded EEG time series data, either in the absence of identifiable behavioral events or in the context of task performance. Granger causality measures between default network areas and signals reconstructed from spontaneous high-density EEG recordings have been reported to be increased during propofol-induced loss of consciousness (Barrett et al., 2012). On the other hand, connectivity assessment using Transfer Entropy, a method used to measure directional information transfer between the EEG signals recorded from different scalp electrodes (Schreiber, 2000), suggests a decrease of front-to-back connectivity in this state (Ku et al., 2011; Lee et al., 2009). This result suggests a possible impairment of top-down cerebral processes during propofol sedation. EEG coherence and Granger Causality connectivity also seem decreased in MCS patients as compared with severely disabled patients who emerged from DOC (Pollonini et al., 2010). Further studies using, for example, intracranial data are warranted to complement these findings.
The limitation of spontaneous brain activity connectivity measures (and related graphical approaches) of not being able to infer the underlying neuronal mechanism can be partially resolved by the use of mathematical models such as DCM (Friston et al., 2003). Correlation-based measures and their variants are also prone to third driver effects, that is, they cannot differentiate between direct causal interactions in brain areas and the influence of a third common generator driving the observed correlation, which would be located somewhere else in the brain. Perturbational approaches may help in reducing this bias by recording brain activity changes directly due to experimentally induced causal neuronal interactions.
Perturbational approach I: TMS-EEG studies
Consciousness is considered as emerging from fast, effective interactions among distributed areas, especially within a cortico-thalamic neuronal complex (Tononi, 2008). One way of effectively gauging connectivity within human thalamo-cortical networks involves directly perturbing a subset of cortical neurons with TMS and performing the source modeling of high-density EEG data to detect, on a millisecond time scale, the chain of effects triggered in the rest of the brain by this initial perturbation (Casali et al., 2010; Ilmoniemi et al., 1997; Litvak et al., 2007; Morishima et al., 2009; Stamm et al., 2011). Since TMS tends to activate a large set of cortical axons in a way that is difficult to fully control (Wagner et al., 2007), TMS-EEG is not ideally suited for capturing causal interactions within specific cortico-cortical circuits. On the other hand, TMS-evoked activations provide an overall, broad estimation of the effective connectivity that may constitute an advantage when studying DOC; indeed, theoretical works (Tononi, 2004; Tononi and Koch, 2008), experimental data (Alkire et al., 2008), and clinical evidence (Markowitsch and Kessler, 2000; Mataro et al., 2001; Schiff, 2010) suggest that consciousness does not depend so much on some specific circuits, but rather on the capacity of distributed regions of the brain to interact through divergent cortico-cortical and cortico-thalamo-cortical connections. Thus, the combination of TMS and EEG was tested in different conditions in which consciousness was physiologically or pharmacologically altered. First, TMS-evoked cortical activations were recorded during the transition from wakefulness to deep non-REM sleep early in the night, when consciousness fades. These measurements showed that, while during wakefulness the brain is able to sustain long-range, complex patterns of activation, during non-REM sleep, this ability is lost: The thalamocortical system, despite being active and reactive, either breaks down into causally independent modules (producing a local response) (Massimini et al., 2005), or bursts into an explosive and nonspecific response (producing a simple, global slow wave that invades most of the cortex) (Bergmann et al., 2012; Massimini et al., 2005, 2007). In a subsequent study, TMS-evoked activations were recorded during REM sleep, when the brain is still disconnected from the external world but consciousness returns in the form of a dream (Massimini et al., 2010). In this condition, TMS-EEG responses were similar to the ones observed during wakefulness. Finally, the spread of TMS-evoked cortical activations was studied during LOC induced by a pharmacological agent, midazolam, at anesthetic concentrations (Ferrarelli et al., 2010). In this case, unconsciousness was associated with a breakdown of the long-range pattern of cortical effective connectivity. Based on these findings TMS-EEG has been proposed as a tool to investigate effective connectivity at the bedside of DOC patients (Massimini et al., 2009). In this category of patients, TMS-EEG may be especially useful, because it does not rely on a subject's ability to process sensory stimuli, to understand and follow instructions, or to communicate. Notably, a recent TMS-EEG study demonstrated a clear-cut recovery of effective connectivity in the brain of noncommunicating patients that paralleled the recovery of consciousness from the VS (Rosanova et al., 2012). Practically, TMS-EEG is still a rather challenging technique that requires the implementation of a complex set-up in the intensive care environment. Hence, simpler ways of estimating effective connectivity in DOC patients should be developed in parallel; this could be achieved by using standard sensory stimulation.
Perturbational approach II: response to external stimuli
Responses to external stimuli have been studied for quite a while in patients with DOC. The first H2 15O-PET studies performed in DOC patients, measuring changes in regional cerebral blood flow, showed that at the group level, VS activation in response to simple auditory and somatosensory stimulation was restricted to primary sensory cortices (Laureys et al., 2000, 2002). Patients in MCS showed a more widespread activation, including fronto-parietal cortices, as well as default network deactivation, in response to external stimuli (Boly et al., 2004, 2008a). fMRI studies showed the presence of the activation of higher-order cortices in some VS patients, which is often correlated with a good prognosis (Coleman et al., 2009; Di et al., 2007).
Recent EEG studies suggested the importance of long-latency components (of more than 100–200 msec delay after the onset of stimuli) in normal conscious perception (Del Cul et al., 2007; Fitzgerald et al., 2001). The amplitude of these long-latency components indeed selectively increases with the probability of perception of a given stimulus. This amplitude also correlates with the level of consciousness in VS and MCS patients (Boly et al., 2011), during propofol anesthesia (Heinke et al., 2004) or during non-REM sleep (Ruby et al., 2008). Previous studies using DCM in HV suggested a privileged role of backward connectivity in the generation of long-latency ERP components (Garrido et al., 2007). Indeed, this work suggests that while the modulation of forward connections is sufficient in a cortical model to explain the generation of the early components of ERP responses, the activity of backward connections is necessary to generate longer-latency responses. Since long-latency components have also been found to correlate with conscious perception, this result suggests that the activity of backward connections could also be important for consciousness. Correspondingly, a recent DCM analysis suggested that backward connectivity from frontal to temporal cortices could be impaired in VS as compared with MCS and controls (Boly et al., 2011). Altogether, these results stress the likely importance of backward connectivity in higher-order cortices for conscious processing (Crick and Koch, 1995; Lamme, 2006). Further studies conducted on sleep and anesthesia, as well as the use of other ERP paradigms (King et al., 2011), will attempt at evaluating the generalizability and diagnostic value of these findings. In addition, other connectivity assessment methods—such as correlation, coherence, Granger Causality, partial directed coherence, or transfer entropy—should also be used to identify dynamical connectivity changes during external stimulation (Bressler and Seth, 2011) in MCS and VS.
The advantages of external stimuli, thus, rely on providing a perturbational approach that investigates cortical connectivity, without the technical burden of TMS-EEG stimulation. ERP amplitudes are more comparable from one subject to another in a between-subject setting, allowing straightforward statistics at the group level (Boly et al., 2011). However, responses to external stimuli may be filtered through afferent pathways and subcortical gates before reaching the thalamo-cortical system. This means, for instance, that brainstem lesions may obliterate response to external stimuli even though thalamo-cortical interactions are not impaired (Massimini et al., 2009). The brainstem function is, however, thought to be essentially preserved in VS and MCS (Laureys et al., 2004).
The temporal blurring of ERP responses to external stimuli—that is, the observed ERP impulse-response duration for a given single input—is much bigger than that of TMS-EEG stimuli. This blurring increases the overlap between the responses in the different brain areas. This potentially decreases the utility of auditory ERP that investigates cortical response differentiation, based on a time series examination of brain activity. An alternative method of studying the complexity of brain responses to external stimuli could be, for example, to study the nonlinearity of the brain responses during responses to stimuli. For example, in the presence or loss of cortical differentiation, the activity in one brain area would be expected to elicit a similar brain activity pattern, with a similar frequency content, in surrounding regions (Boly et al., 2009a). In contrast, the observed brain activity pattern would be very different in the presence of differentiated and integrated cerebral activity (Alkire et al., 2008). Investigating cross-frequency interactions between brain regions could, therefore, be another way of investigating the complexity of cerebral activity in response to stimulation (Ahrens et al., 2008). Following this logic, cross-frequency interactions could be expected to decrease in anesthesia, sleep, and DOC. Previous studies suggested the preponderance of nonlinearity in backward connections, as compared with forward connections (Chen et al., 2009). The integrity of cerebral backward connectivity could, thus, be hypothesized as being correlated with preserved brain activity complexity in patients with DOC. Further studies should, however, investigate this issue in greater detail.
Diagnosing Consciousness: Level Versus Contents
Figure 2 summarizes the present section—which will aim at reviewing the conceptual and practical differences between the study of the level versus the contents of human consciousness, and at emphasizing their relevance for the study of brain function in DOC patients.
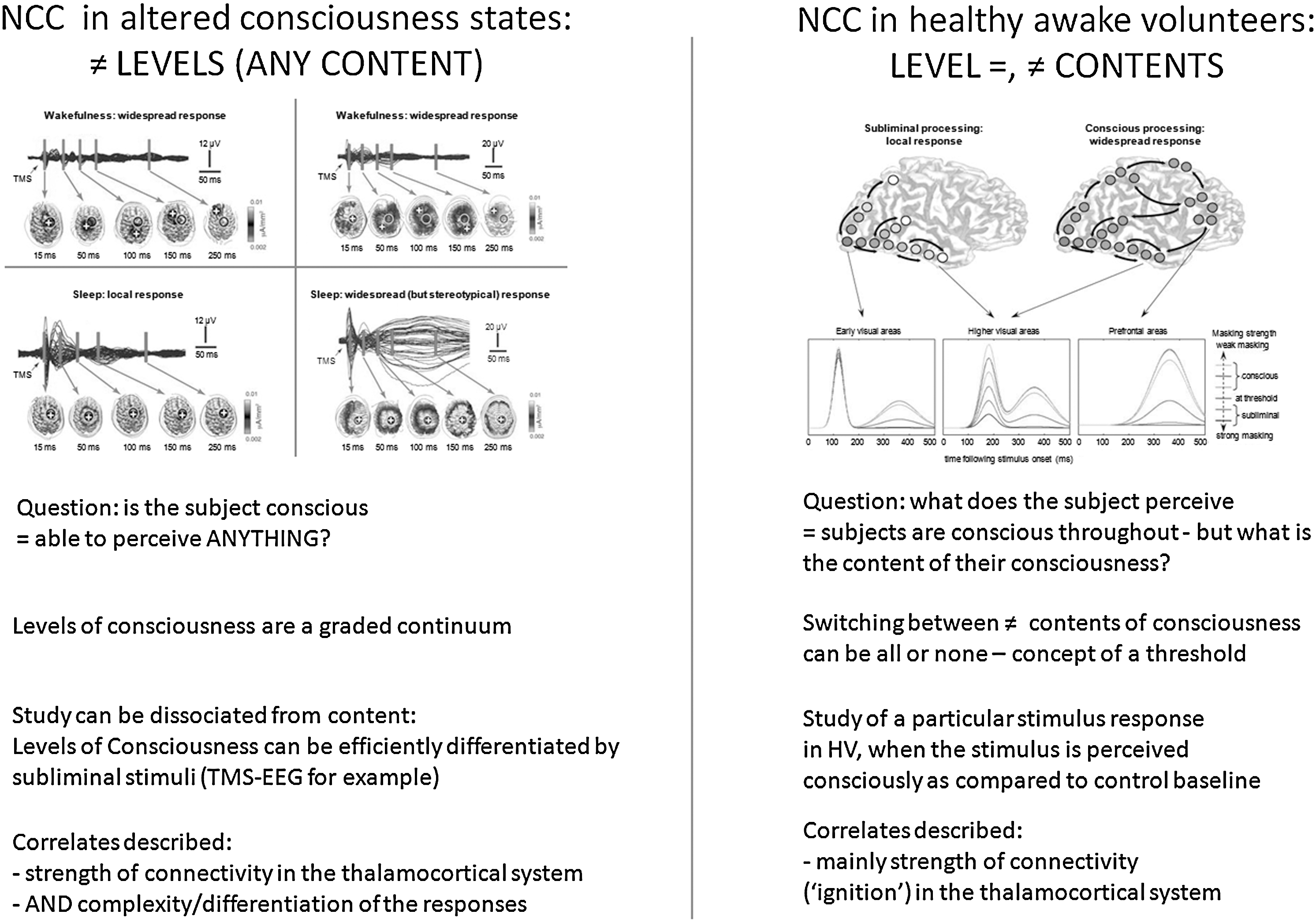
Conceptual and practical differences in the study of NCC in states such as coma, anesthesia, and sleep, or in healthy awake volunteers. Detailed description can be found in the main text. Upper panels are reproduced with permission from (Alkire et al., 2008; Del Cul et al., 2007). NCC, neural correlates of consciousness; HV, healthy awake volunteers.
As previously discussed by several authors in the literature (Baars, 1988; Frith, 2011; Zeman, 2001), consciousness continues to be an ill-defined concept. When scientifically approaching NCC, there is an important distinction to be made between the level and the contents of consciousness (Dehaene and Changeux, 2005; Frith, 2011; Overgaard and Overgaard, 2010). In the present context, consciousness is defined as what disappears during dreamless sleep (Tononi, 2008): the ability to perceive anything. The contents of consciousness refer to any of the particular perceptions that can be experienced during a normal wakefulness state. These contents may be internally driven or generated through an interaction with the environment (Vanhaudenhuyse et al., 2011). The level of consciousness refers to a dimension that varies from coma at one extreme, through sleep, and to alert wakefulness at the other extreme (Frith, 2011). The level of consciousness can in this view be compared with a potential to experience a lot of different contents, though each cognitive or sensory function by itself would not be necessary for the existence of consciousness (Boly and Seth, 2012). By definition, this potential would disappear in unconscious states. Understanding the levels of consciousness is of particular relevance to the studies of DOC patients, as well as to brain function in different stages of anesthesia and sleep. Ethically, detecting a potential ability to perceive—anything—is a very important question in noncommunicative severely brain-damaged patients.
Alert wakefulness is characterized by a consciousness of an ever-changing stream of specific mental states—comparable to a wandering exploration of the potential repertoire of states available to our consciousness at each single moment (Sadaghiani et al., 2010). These transient states that we are aware of at each instant determine the contents of our consciousness. In studies of the NCC in HV, there is great interest in contrasting the neural activity associated with stimuli that influence the contents of consciousness with the neural activity associated with the same stimuli when they affect behavior in the absence of any change in the content of consciousness (Frith et al., 1999). The key difference between the study of NCC in HV and DOC patients is, however, that in the first case, subjects are basically conscious all the time—what changes is the content of their experiences at particular experimentally controlled moments (Boly and Seth, 2012). As noted by (Frith, 2011), though a certain minimal level of consciousness is necessary for there to be any contents of consciousness, the level of consciousness does not determine what the contents of consciousness will be. A certain level of consciousness is prerequisite to any perception, but a series of other factors—such as stimulus intensity, duration or emotional valence, underlying brain activity shaped by previous learning, and ongoing spontaneous brain activity fluctuations—can also influence the conscious access of some given stimuli (Boly et al., 2008b; Dehaene and Changeux, 2005). As stated in (Dehaene and Changeux, 2011), theoretical approaches to NCC in HV—that is, NCC of conscious access, or consciousness contents—and in DOC—that is, NCC of the conscious level—are from two different scopes. In principle, theoretical predictions in terms of NCC for these two aspects could, thus, be different.
In practice, there exists some possible dissociation between the experimental study of consciousness level and that of consciousness contents. One can, for example, differentiate various levels of consciousness, without asking any questions about conscious contents. In principle, this question can be addressed via the study of spontaneous brain activity, in an experimental setup lacking control on the subjects' conscious contents. This approach is commonly used in resting-state fMRI or spontaneous EEG acquisition studies. In the context of perturbational approaches, some subliminal stimulations have also been shown to efficiently differentiate the levels of consciousness across different conditions. The TMS-EEG stimulation is typically not perceived by HV and, in common experimental settings, does not overtly affect the subjects' conscious contents. Despite its subliminal characteristic, TMS responses to stimulation have been shown as efficiently discriminating the levels of consciousness in states such as anesthesia (Ferrarelli et al., 2010), severely brain-damaged patients (Rosanova et al., 2012), or sleep (Massimini et al., 2005). Similarly, simple auditory stimulations such as clicks (Boly et al., 2004) or beeps used in a mismatch negativity paradigm (Boly et al., 2011; Heinke et al., 2004; Ruby et al., 2008)—in which processing is commonly considered to be pre-attentive—could act by probing some basic characteristics of the system that are necessary for some level of consciousness to emerge (Massimini et al., 2009), without necessarily significantly influencing the subjects' conscious contents. Another example can be found in the results of some experiments performed during REM sleep. During this sleep stage, external sounds are usually hardly perceived, but ERP still look very similar to those observed in wakefulness. The observed brain response to auditory stimulation is, thus, likely uncovering a relatively preserved level of consciousness, without being a signature of a modification of its contents (Nir and Tononi, 2010). These different examples show that, in practice, the basic NCC of the level of consciousness can be studied independently of consciousness contents.
As shown in Figure 2 (left panel), the neural correlates of a reduced level of consciousness consist either in a local response to the stimulation (Massimini et al., 2005) or in a global, nonspecific response (Alkire et al., 2008; Kroeger and Amzica, 2007; Massimini et al., 2007). This phenomenon is generally thought to be due to the presence of bistable dynamics in the system (Massimini et al., 2012), which would prevent the emergence of complex spatiotemporal patterns of effective connectivity that are widespread and differentiated at the same time. Similarly, patterns of activity that are widespread, but stereotypical, should be expected during the loss of consciousness in epileptic patients. From this perspective, the level of consciousness seems to correlate better with the complexity of brain activation than with their spatial extent. Thus, quantifying the complexity of brain responses may be a necessary step if one wants to reliably assess the level of consciousness in noncommunicative brain-damaged patients (Boly, 2011; Boly et al., 2009a; Massimini et al., 2009). On the other hand (Fig. 2, right panel), the neural correlates of conscious perception of a specific stimulus in HV invariably consist of a stronger, and more diffuse, activation for perceived stimuli as compared with subliminal stimuli. This “ignition” process is characterized by the presence of stronger long-range coherence and fast frequency responses involving fronto-parietal cortices for stimuli that reach the threshold for consciousness (Dehaene et al., 2006). In this case, observing a widespread response may immediately signal that a subject consciously perceives the particular stimulus she/he is presented with. In unconscious subjects, it will certainly be important to better understand, and to better conceptualize, the possible dissociations between the neural correlates of the level of consciousness and the neural correlates of a specific aspect of consciousness. If used in severely brain-damaged patients, a single criteria of big response/activation for deciding the presence of consciousness presents, however, a risk of false positive, due to the occurrence of high amplitude responses commonly present in DOC—similar to the K-complex responses observed during non-REM sleep (Boly et al., 2009a). An additional check for the complexity of brain response and the presence of brain activity differentiation would, thus, be a safer bet in order to reliably detect the consciousness in DOC patients (Boly, 2011).
Toward the Use of Neuroimaging as a Diagnostic Test?
To date, the gold standard for diagnosis of the level of consciousness is behavioral. Clinical assessment is based on criteria described by (Giacino et al., 2002) to distinguish MCS from VS. However, this clinical diagnosis itself has its ownlimitations. If not using an appropriate behavioral scale, clinical misdiagnosis reaches up to 41% (Schnakers et al., 2009). Clinical scales themselves are quite variable in their reliability. A recent meta-analysis revealed that the CRS-R was the best attempt that explicitly incorporates the diagnostic criteria for MCS in clinical diagnosis (Seel et al., 2010). However, the reliability of the CRS-R itself depends on examiner experience (Lovstad et al., 2010), and this scale is quite long and heavy to administer in a routine basis in acute patients. Even if this gold-standard behavioral assessment is repetitively and carefully performed in DOC patients, it can also sometimes lead to some underestimation of the patients' level of consciousness. It has indeed been shown that a minority of patients who are behaviorally totally unresponsive instead become conscious when assessed with active paradigms using fMRI (Bardin et al., 2011; Monti et al., 2010; Owen et al., 2006) or EEG (Cruse et al., 2011). On the other hand, the active neuroimaging paradigms used to detect these patients are also prone to false negatives (Bardin et al., 2011). In fact, in principle, paradigms that are based on response to command, being clinical or neuroimaging based, are not sufficient to systematically detect consciousness in each individual case (Boly, 2011). Indeed, volition does not equate consciousness: The absence of a response from the patient does not necessarily mean that the patient is unconscious (Boly et al., 2007) and even in HV, volitional activity and consciousness can, in many cases, be dissociated (Boly and Seth, 2012; Soon et al., 2008). Recent results obtained using the isolated forearm technique during anesthesia (Russell and Wang, 1996) raise even more debate on this topic, by showing frequent responses to command in the absence of a subsequent report. To date, there is also a need to fill the gap between clinical behavioral studies and neuroimaging studies by comparing the results obtained using these two approaches on a single-subject basis and identifying typical VS and MCS patterns, versus outlier patterns. Different neuroimaging techniques could, thus be compared in terms of their performance—false-positive and false-negative rates, sensitivity, specificity, and so on—if used in sufficiently large populations. Inter-assessor and inter-site reliabilities should also be assessed in order to evaluate the reproducibility of different techniques results on a larger scale. Finally, the use of automated procedures (Soddu et al., 2012), as well as the use of classifiers (Phillips et al., 2011), and of a combined study of different imaging techniques (Bruno et al., 2011) could also reduce observer-dependent bias in neuroimaging-based diagnosis.
Ultimately, a most accurate diagnosis of consciousness would require first knowing the mechanisms bridging conscious perception to the brain and then using neuroimaging techniques as a way of objectifying the presence of this mechanism. To achieve this aim, theoretical approaches attempting at describing the general mechanism underlying conscious experiences (Dehaene and Changeux, 2011; Tononi, 2008), as well as the potential use of theoretically grounded indexes for the neuroimaging-based diagnosis of consciousness (Seth et al., 2008, 2011), are of great interest. However, a lot more work is needed in this field, and we are still a long way from bringing these theories to truly testable predictions (Boly and Seth, 2012). Meanwhile, an NCC-based assessment approach, optimally using the vast information already present in the literature concerning brain function in different DOC states, may provide valuable information about the potential ability of a noncommunicative patient's brain to allow consciousness—information that could usefully complement the clinical diagnosis.
Footnotes
Acknowledgments
This work was supported by the Belgian Fonds National de la Recherche Scientifique(FNRS), European Commission, Mind Science Foundation, McDonnell Foundation, French Speaking Community Concerted Research Action (ARC 06/11-340), and Fondation Léon Frédéricq. O.G. is Research Fellow, M.B. and Q.N. are Postdoctoral Fellows, and S.L. is Senior Research Associate at the FNRS. M.G. is supported by the Wellcome Trust.
Author Disclosure Statement
The authors declare no conflict of interest.