
Abstract
Neuroimaging studies of professional athletic or musical training have demonstrated considerable practice-dependent plasticity in various brain structures, which may reflect distinct training demands. In the present study, structural and functional brain alterations were examined in professional badminton players and compared with healthy controls using magnetic resonance imaging (MRI) and resting-state functional MRI. Gray matter concentration (GMC) was assessed using voxel-based morphometry (VBM), and resting-brain functions were measured by amplitude of low-frequency fluctuation (ALFF) and seed-based functional connectivity. Results showed that the athlete group had greater GMC and ALFF in the right and medial cerebellar regions, respectively. The athlete group also demonstrated smaller ALFF in the left superior parietal lobule and altered functional connectivity between the left superior parietal and frontal regions. These findings indicate that badminton expertise is associated with not only plastic structural changes in terms of enlarged gray matter density in the cerebellum, but also functional alterations in fronto-parietal connectivity. Such structural and functional alterations may reflect specific experiences of badminton training and practice, including high-capacity visuo-spatial processing and hand-eye coordination in addition to refined motor skills.
Introduction
T
Compared with structural changes in skilled professionals, evidence supporting functional brain alterations in elite athletes or musicians is relatively limited. Using functional magnetic resonance imaging (fMRI), two recent studies examined neural activations during preshot routines of expert golfers (Milton et al., 2007) and archers (Kim et al., 2008). The results demonstrated that professionals tended to recruit a more focused and efficient organization of visual and motor networks directly related to task requirements, whereas novices tended to engage more brain regions related to high-level cognitive control functions. However, findings from these task-related fMRI studies depend heavily on the task demands and specific cognitive or motor paradigms used in the studies, which makes the comparison of results across studies difficult.
The use of resting-state functional connectivity fMRI (fc-fMRI) that examines functional brain changes have been emerging since the first report of functional connectivity between bilateral motor cortices (Biswal et al., 1995). Resting-state fc-fMRI measures the synchronization of low-frequency blood oxygen level dependent (BOLD) oscillations between spatially distinct brain regions in the absence of tasks. Therefore, it provides a complimentary method to the conventional task-related fMRI for detecting functional brain changes associated with training and experience. Since resting-state fc-fMRI does not involve any particular motor or cognitive task requirements, it may reflect a cumulative effect of experience over time (Lewis et al., 2009). Recent fc-fMRI studies have suggested that motor training altered resting-state brain activity (Xiong et al., 2009; Albert et al., 2009). For example, Albert et al. (2009) examined resting-state functional brain changes during motor training using independent component analysis (ICA) and found that motor learning, but not motor performance, significantly modulated resting-state functional connectivity in the fronto-parietal and cerebellar networks. However, to our knowledge, the effects of long-term training on resting-state brain function remain largely unknown.
In this study, we used MRI to examine both structural and resting-state functional brain alterations in professional racquet players, specifically badminton players. To the best of our knowledge, structural and functional characteristics of the badminton players' brain have not been explored using neuroimaging methods. Similar to other racquet sports, badminton requires refined hand-eye coordination and visuo-spatial ability (Ward et al., 2002; Shim et al., 2005), which may result in structural and functional brain changes that are different from other sports that have been studied, for example, golf, diving, or basketball. We predict that professional badminton players will show significant structural and functional alterations in brain regions related to visuo-spatial processing and hand-eye coordination. A group of professional badminton players and matched nonathlete controls were recruited in the present study. Standard voxel-based morphometry (VBM) (Ashburner and Friston, 2000) was first conducted to examine the structural differences between the two groups. The amplitudes of low-frequency fluctuations (ALFF; Zang et al., 2007) of resting-state fMRI were then used to measure regional properties of the brain's intrinsic neural activity and compared between the two groups. Finally, the seed-based functional connectivity analyses (Fox et al., 2005) of resting-state fMRI were conducted to characterize functional integration, using regions that have been shown to be different between professional players and controls from the VBM and ALFF analyses. We chose to use the hypothesis-driven, seed-based functional connectivity analysis instead of data-driven methods such as ICA in order to determine the specific brain regions and networks that are related to cultivated visuo-spatial processing and hand-eye coordination skills after long-term badminton training and practice.
Methods
Subjects
Twenty badminton players (10 males) were recruited for the study. All subjects were members of professional teams or professional university teams in Beijing and had completed at least 3 years of badminton training (mean=8.9 years, range from 3 to 16 years). Subjects had either completed college-level education or were current college students during the study. The control group consisted of 18 age- and gender-matched college students (9 men). No control subjects had formal training or experience in badminton or other racquet sports (Table 1). All athletes and controls were right handed. Written informed consent was obtained from all subjects at the beginning of the study.
MRI scan
All subjects were scanned using a Siemens 3T Trio scanner located in the Beijing Center for Magnetic Resonance Imaging, Beijing, China. Subjects first completed a task-independent, resting-state scan, during which time they were asked to open their eyes to look at a blank screen. No additional task was required. The resting scan lasted 6 min, with a TR of 2 sec. As a result, 180 images were obtained for each subject. BOLD images were obtained using an echo-planar imaging sequence with a 12-channel coil. Acquisition parameters were as follows: TE=30 ms; TR=2000 ms; flip angle=90o; 32 interleaved axial slices; slice thickness=4 mm, no gap; image matrix=64×64; FOV=200×200 mm; bandwidth=2232. At the end of the scan, a high spatial resolution, T1-weighted magnetization prepared rapid gradient-echo (MP-RAGE) structural image was obtained. Scan parameters were as follows: TR=2530 ms; TE=1.64; flip angle=7.0; 176 slices; slice thickness=1 mm; image matrix=256×256; FOV=256×256 mm.
VBM analysis
Standard VBM analysis was conducted using SPM8 software (
Preprocessed GMC images were then entered into a second-level, two-sample t-test model to compare group level differences. Total Gray matter volume (GMV), gender, and age were added to the model as covariates. Since the smoothness of GMC images is nonstationary (Ashburner and Friston, 2000), nonstationary cluster extent correction was adopted to correct multiple comparisons (Worsley et al., 1999). The amplitude threshold was first set at p<0.001, and then, the cluster extent was corrected at p<0.05 using the nonstationarity toolbox (Hayasaka et al., 2004). The clusters identified in the group analysis were labeled using the Talairach Daemon (Talairach and Tournoux, 1988; Lancaster et al., 2000) after accounting for the discrepancy between MNI space and Talairach space (Lancaster et al., 2007). This analysis revealed two clusters in the right cerebellum that were defined as the seed regions for further functional connectivity analysis.
Resting-state fMRI data preprocessing
Preprocessing of resting-state functional images was completed using SPM8. First, the two initial resting-state images were discarded for each subject, leaving a total of 178 images per subject. All functional images were motion corrected using the realign function. Functional images from two athletes and two controls were discarded, because at least one of the motion parameter estimates was larger than 3 mm or 3 degrees. As a result, images from 18 athletes and 16 controls were included in the subsequent analysis. For each subject, the functional images were first registered to the subject's own high-resolution structural image. The structural image was then normalized to the T1 template. Next, all functional images were normalized to the standard MNI space using the parameters obtained from normalizing the subject's structural images. Finally, all normalized functional images were smoothed using a Gaussian filter with an 8 mm FWHM kernal.
ALFF analysis
We used the ALFF to index regional properties of the brain's intrinsic neural activity. ALFF analysis was completed using the resting-state fMRI data analysis toolkit, (REST) v1.4 (
Functional connectivity analysis
Functional connectivity analysis was conducted using the functional connectivity toolbox, v.12.p (Chai et al., 2012;
Results
VBM results
The mean total GMV of the athlete group was 699.1 mL (SD=60.2), while the mean total GMV of the control group was 717.7 mL (SD=59.3). There was no difference in total GMV observed between the two groups (t=−0.96, p=0.35).
A voxel-wise comparison revealed greater GMC in two right cerebellum clusters for the athlete group than the control group (Fig. 1 and Table 2). One cluster was located in the posterior right cerebellar tonsil (lobule 8), and the other region was located in the anterior right cerebellar hemisphere (lobule 6). However, no region showed greater GMC for the control group than the athlete group.

Larger gray matter concentration in the athlete group than the control group. Threshold was set as cluster level false discovery rate (FDR) corrected p<0.05. L: left; R: right.
Threshold was p<0.001 with a cluster-level nonstationary correction of p<0.05.
R, right; L, left; MNI, Montreal Neurological Institute.
The voxel-wise comparison of GMV showed no differences between the two groups using a threshold of p<0.05 with nonstationary correction. However, greater GMV in similar cerebellum clusters was observed using a more liberal threshold of p<0.001 and cluster size>141 voxels, (Table 3).
Threshold was p<0.001 with a cluster extent of 141 voxels. The clusters in bold corresponded to the clusters reported in Gray matter concentration analysis (Table 2).
ALFF results
Larger ALFFs were also found in the cerebellum for the athlete group than the control group (Fig. 2 and Table 4), including the anterior part of the culmen, and the medial portion of the anterior lobe (vermis lobule 6). In contrast, there was a cluster in the left superior parietal lube (BA 7/19) that revealed larger ALFFs in the control group than the athlete group.
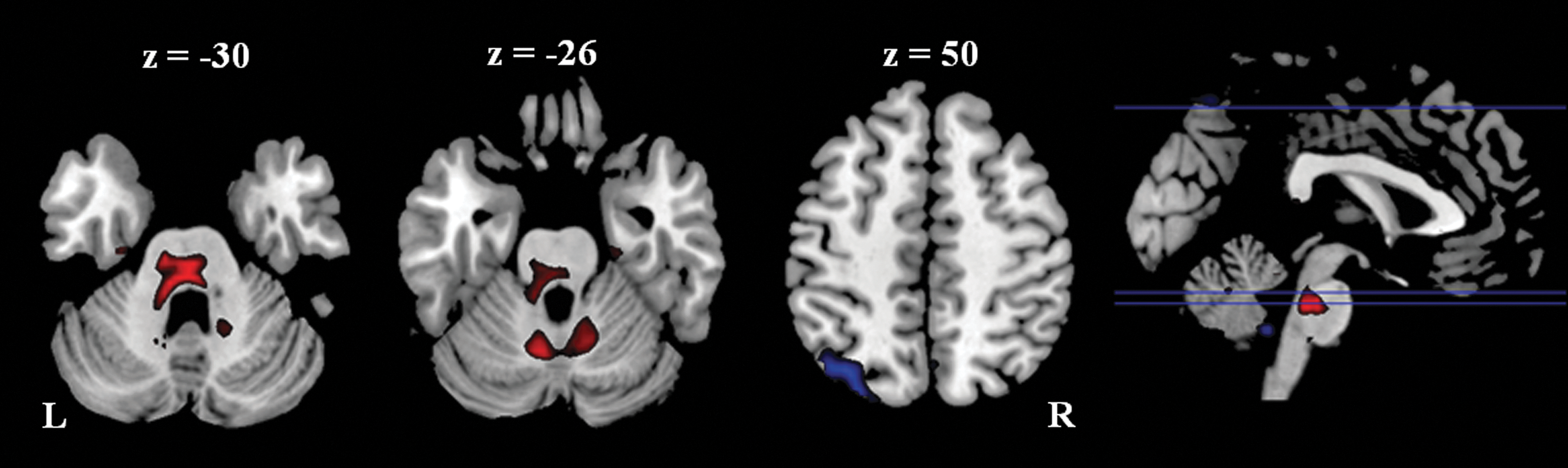
Different amplitude of low-frequency fluctuation between the athlete group and the control group. Threshold was set as cluster level FDR corrected p<0.05. Red: athlete>control; Blue: control>athlete. L: left; R: right.
Threshold was p<0.001 with a cluster-level false discovery rate correction of p<0.05.
BA, Brodmann's area.
Functional connectivity analysis
Functional connectivity analysis for all subjects revealed that seeding regions belonged to distinct functional networks (Fig. 3). The right posterior cerebellum seed showed positive functional connectivity to the left dorsal-lateral prefrontal cortex, left parietal cortex, and left temporal lobe (Fig. 3B); while the right anterior cerebellum and the medial part of the cerebellum seeds showed positive connectivity with bilateral sensorimotor areas (Fig. 3D, F). The left superior parietal lobule seed showed connectivity with typical task-positive networks, including the bilateral dorsal lateral prefrontal cortex, the bilateral parietal cortex, the bilateral MT lobe, and the bilateral posterior cerebellum (Fig. 3H).
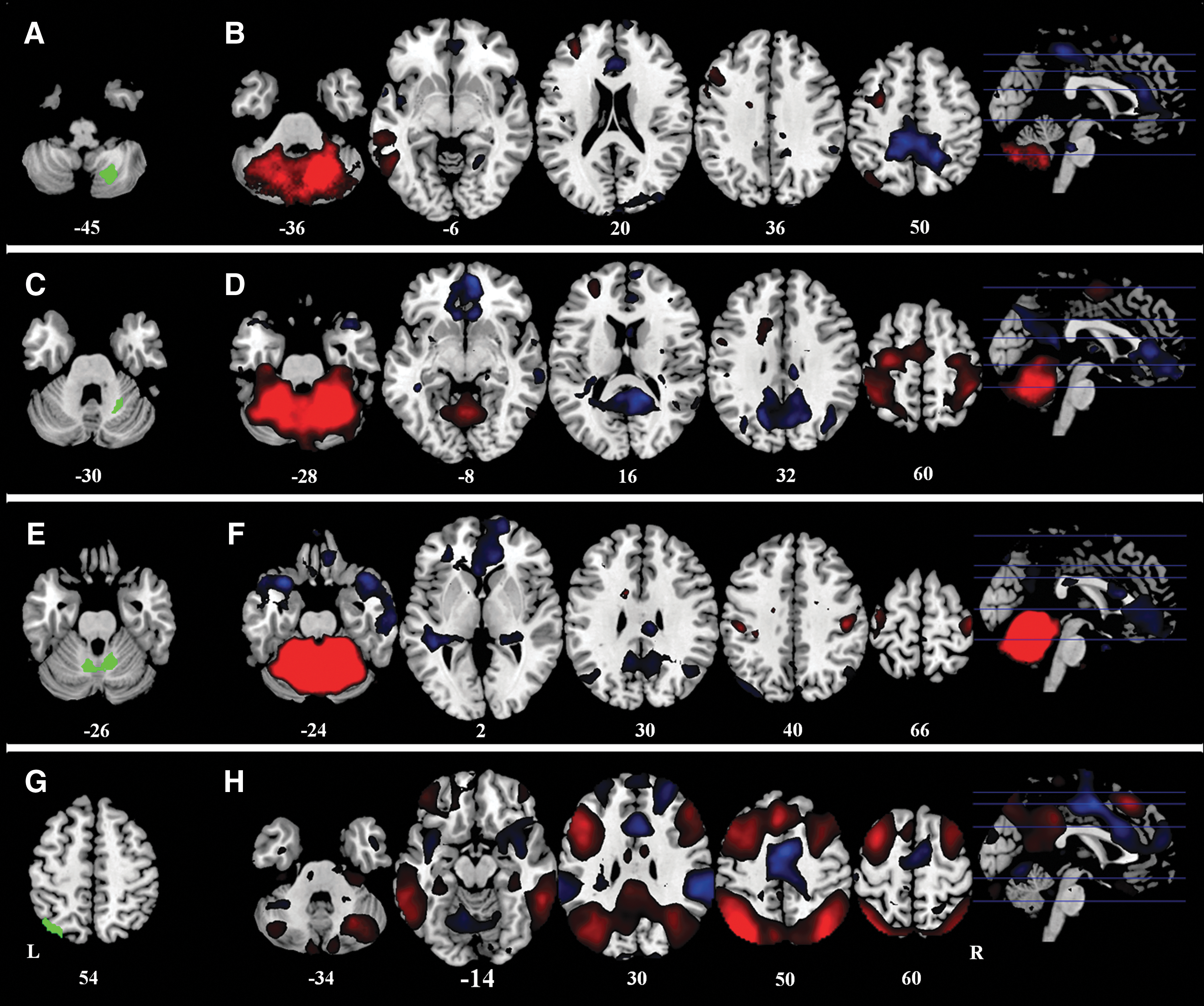
Positive (red) and negative (blue) correlation map of all the subjects with the four seeding regions.
No differences were found in functional connectivity maps of both groups when the right posterior and the right anterior cerebellum regions were used as seeds, respectively. The medial cerebellum seed region revealed less functional connectivity with the right anterior cingulate cortex in the athlete group than the control group (Fig. 4). However, this cluster was mainly located in the WM; therefore, it was not further discussed.
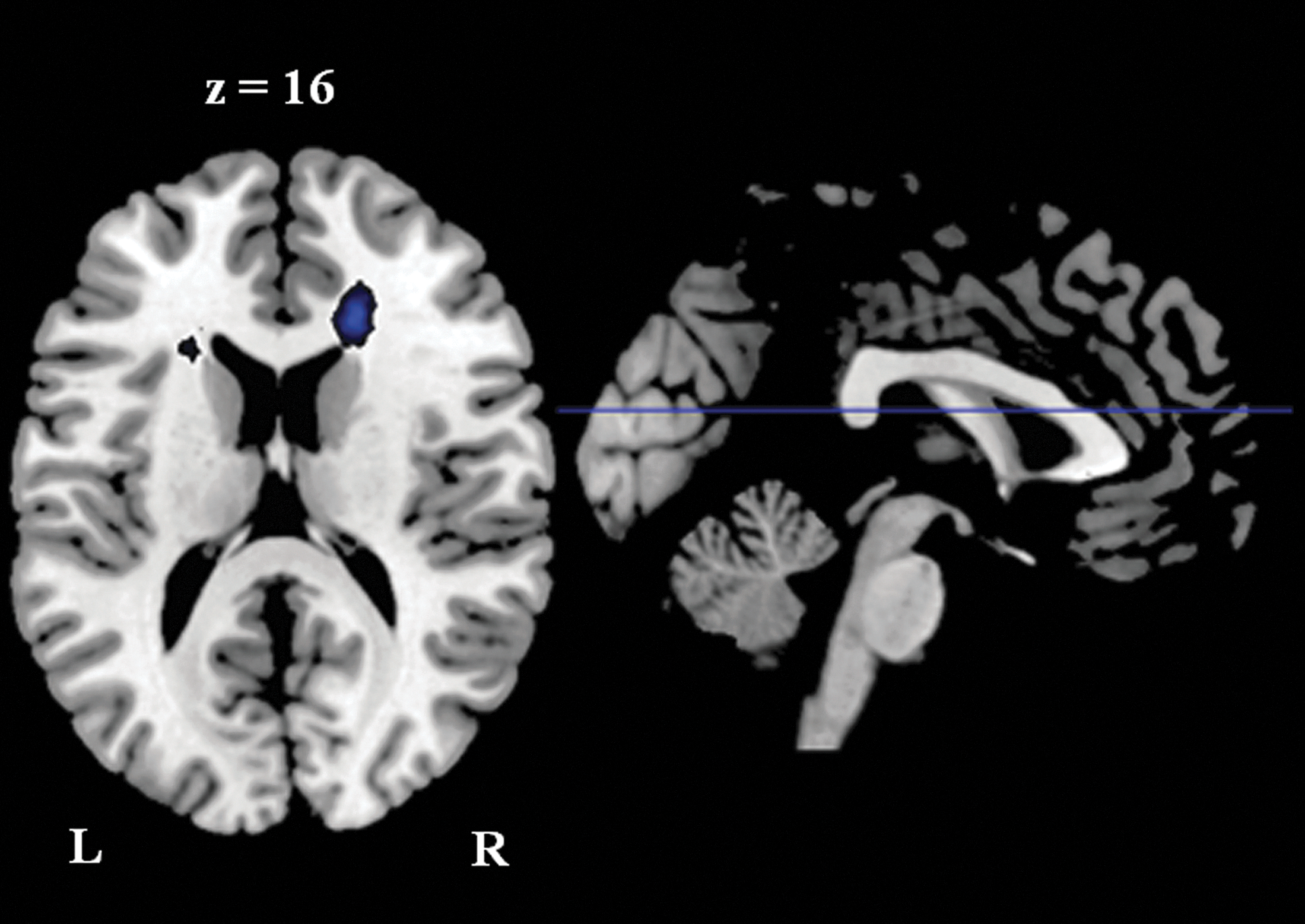
Cluster showed reduced functional connectivity with the medial cerebellum seed region in the athlete group than in the control group. Threshold was set as cluster level FDR corrected p<0.05. L: left; R: right.
The left parietal seed region showed greater functional connectivity with the right anterior cingulate cortex (BA 32/24) and the left middle frontal cortex (BA 6), and weaker functional connectivity with the left inferior frontal cortex (BA 9) and the bilateral middle frontal cortex (BA 9) in the athlete group than the control group (Fig. 5 and Table 5). Using a positive correlation map of all subjects as an inclusive mask, we further confirmed that the two left middle frontal cortex clusters, which showed different functional connectivity with the left superior parietal lobule between the two groups, belonged to the positive correlation network (Fig. 6).
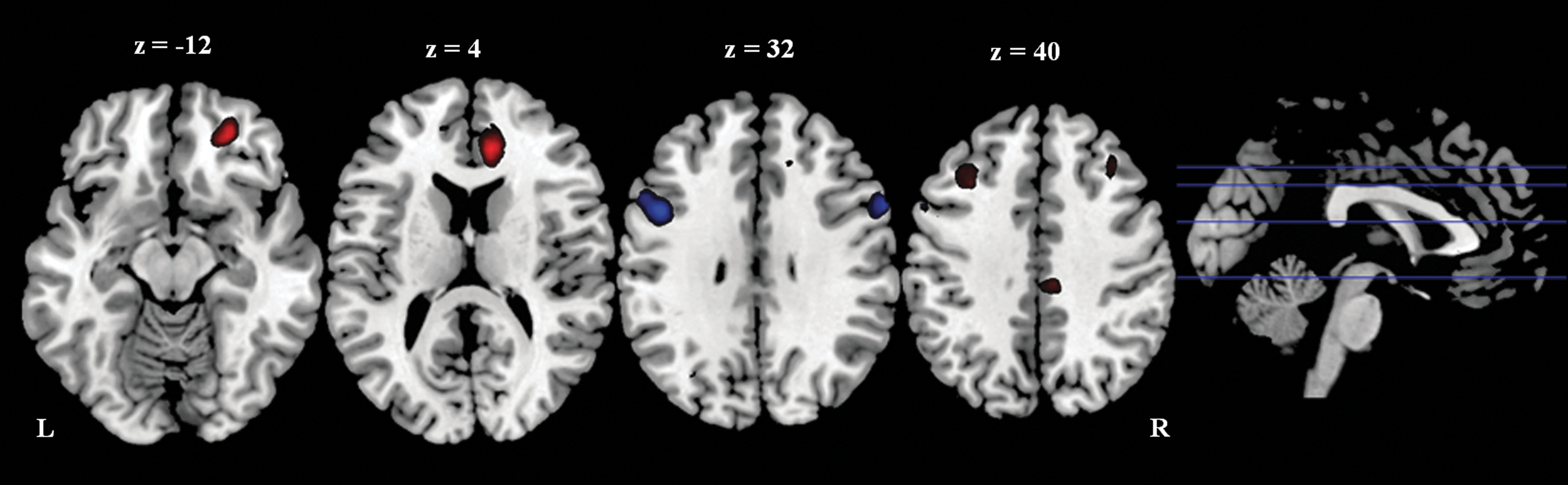
Cluster showed different functional connectivity with the left parietal seed region between groups. Threshold was set as cluster level FDR corrected p<0.05. Red: athlete>control; Blue: control>athlete. L: left; R: right.
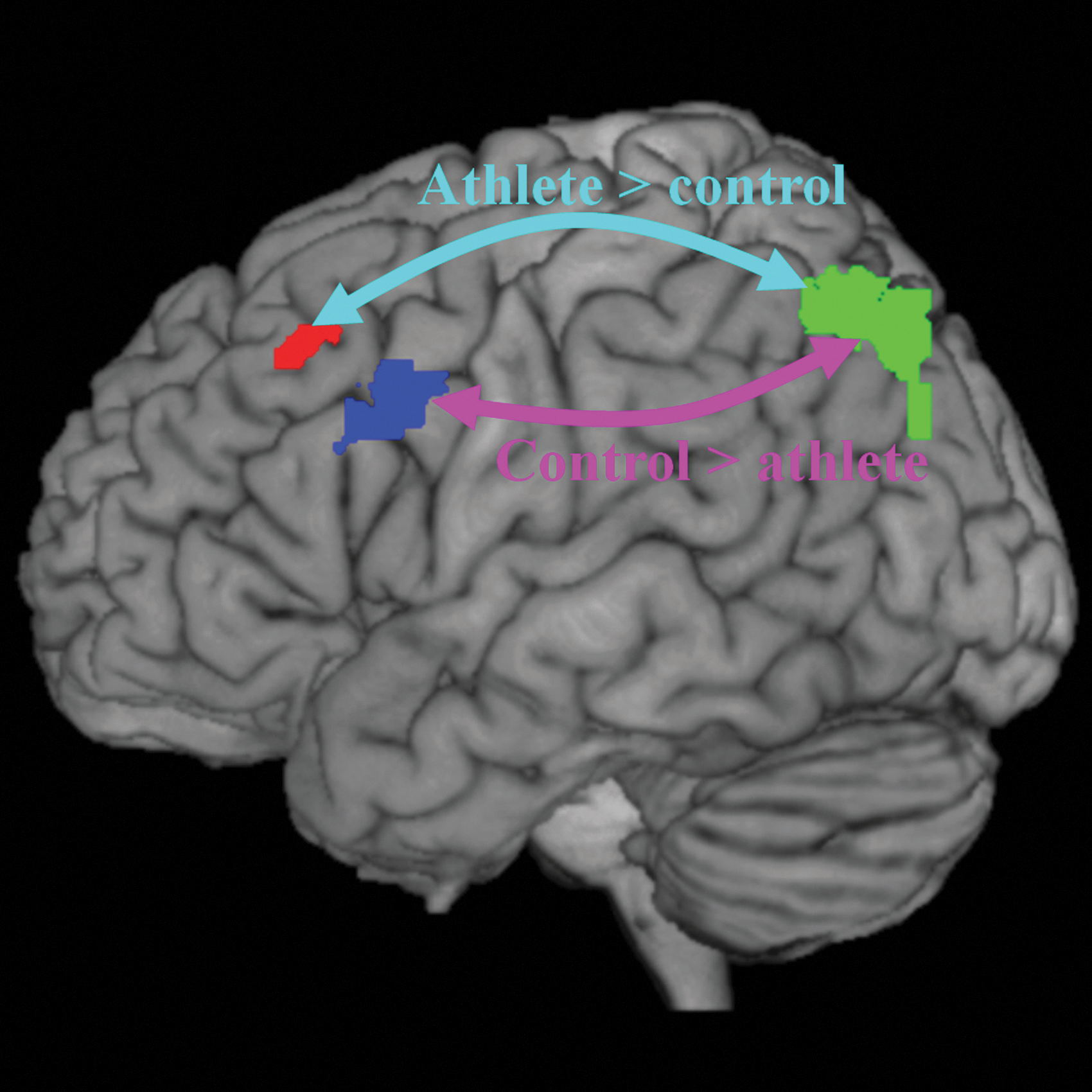
Illustration of different functional connectivity between the athlete group and controls within the left fronto-parietal network. Arrow in cyan represents enhanced functional connectivity between the left superior parietal lobule (green) and the left middle frontal gyrus (BA6) (red) in the athlete group than in the control group. Arrow in purple represents lower functional connectivity between the left superior parietal lobule (green) and the left middle frontal gyrus (BA9) (blue) in the athlete group than in the control group.
Threshold was p<0.001 with a cluster-level false discovery rate correction of p<0.05.
Discussion
The two main findings of the present study are as follows: First, badminton athletes demonstrated greater GMC in the right anterior and posterior lobes of the cerebellum and greater ALFFs in the medial portion of the cerebellum. Second, badminton athletes showed smaller ALFFs in the left superior parietal lobule and altered functional connectivity between the left superior parietal lobule and frontal regions.
The role of the cerebellum
The greater cerebellum GMC in badminton players is consistent with other cross-sectional studies of athletes, such as basketball players (Park et al., 2009) and keyboard musicians (Hutchinson et al., 2003). Structural expansion may result from microstructural changes at the neuronal level (Anderson et al., 1994; Kleim et al., 1997; Kleim et al., 1998). Changes in the cerebellum were also observed in the ALFF analysis, which is consistent with the recent finding that short-term motor training enhances resting-state independent cerebellar component activity (Albert et al., 2009). There is also evidence that task-related neural responses, as measured by fMRI, may be shaped by motor training (Doyon et al., 2002; Jenkins et al., 1994; Jueptner et al., 1997). These findings suggest that the cerebellum plays a key role in long-term professional badminton training and practice. However, the current cross-sectional study cannot rule out the alternative explanation that individuals with larger cerebellar density, and altered intrinsic activity might be more likely to master the fine motor dexterity that is necessary for elite badminton skills. Future studies using a longitudinal design are needed to answer these questions.
It is noteworthy that Hutchinson et al. (2003) and Park et al. (2009) only measured the global volume of the cerebellum or cerebellar lobules. The voxel-wise method adopted in the present study enabled the determination of region-specific anatomical differences within the cerebellar lobes. Using functional connectivity analysis, we further demonstrated that regions of the cerebellum which showed structural differences with VBM analysis were not restricted solely to the motor network. The right posterior cerebellum region is functionally connected to the left dorsal-lateral prefrontal cortex, the left parietal cortex, and the left temporal lobe, suggesting that the posterior cerebellum region belongs to the left fronto-parietal executive network (Habas et al., 2009). Therefore, the expansion of the cerebellum resulting from motor training may not only support motor control functions, but also involve other nonmotor functions such as visuo-motor coordination and executive control.
It is also worthy to point out that the nonmodulated GMC from VBM analysis did not compensate for the effect of spatial normalization; therefore, it cannot reflect absolute volume changes (Mechelli et al., 2005). Instead, the GMC findings might reflect the differences of relative concentration or the density of cerebellum in the two groups. However, the GMV analysis after modulation showed very similar results, supporting the modulation of badminton training on the cerebellum structure.
The role of the fronto-parietal network
The present study found, for the first time, that the function of the fronto-parietal network is altered in athletes. First, regional ALFFs in the left superior parietal lobule were smaller in the athlete group. The superior parietal lobule is thought to act as a relay that passes information from visual-processing areas to motor-processing areas, regions which support visuomotor coordination (Caminiti et al., 1996; Wise et al., 1997). Enhanced neural activity was observed in the superior parietal lobule after short-term motor training (Jenkins et al., 1994; Jueptner et al., 1997; Sakai et al., 1998). However, lower ALFFs appear to be inconsistent with a recent finding that the strength of the fronto-parietal network increased after 11 min of visuomotor training (Albert et al., 2009). We argue that this finding may reflect different consequences of training phases between short-term training in Albert et al. (2009) and long-term athletic training in the present study. This notion is consistent with Xiong et al. (2009), who showed that region-specific brain activation increases first, then decreases, during a longer period of motor training.
In addition to the amplitude of fluctuation, our results reveal altered connectivity within the fronto-parietal network. The fronto-parietal network, which is supported by the underlying superior longitudinal fasciculus (Makris et al., 2005), plays a key role in hand-eye coordination (Battaglia-Mayer et al., 2001; Marconi et al., 2001), visual-guided reaching (Battaglia-Mayer et al., 2003; Burnod et al., 1999), and grasping (Grol et al., 2007). This is consistent with the notion that badminton athletes have higher visuomotor skills than individuals who do not play racquet sports. In addition, this network is reported in key regions to support the anticipation of badminton serving (Wright et al., 2010; Wright and Jackson 2007).
Our results suggest that functional connectivity has been rewired within the fronto-parietal network (Fig. 6) in badminton athletes but not controls. In the athlete group, the left superior parietal lobule revealed enhanced functional connectivity with the premotor cortex (BA6); whereas in the control group, the left superior parietal lobule revealed stronger functional connectivity with the middle frontal gyrus (BA9). Since the premotor cortex supports the sensory guidance of movement (Passingham, 1985), this result indicates that the fronto-parietal network in the athlete group may involve superior motor-related processing in comparison with the control group, thus reflecting a cumulative effect of the motor training or the prerequisition of superior visual-spatial capability in professional badminton athletes.
Conclusion
In conclusion, the present study revealed significant structural and functional alterations in professional badminton players compared with normal controls. Specifically, badminton players showed altered intrinsic neural activity and functional integration in the fronto-parietal network in addition to structural changes in the cerebellum. These alterations may reflect specific task demands and cognitive processes during badminton training, including refined visuo-spatial processing and hand-eye coordination in addition to motor skills.
Footnotes
Acknowledgments
This research was supported in part by the National Nature Science Foundation of China Grants 31070984, 30770730, 30921064, 31171082, 90820307, and 91124004; the National Basic Research Program of China 2012CB720701; and NIH Grants R01 HL102119, R21 DA032022, and R03 DA027098. Tha authors thank Marc Korczykowski for his helpful comments.
Author Disclosure Statement
No competing financial interests exist.